
Biochemistry of the Endocrine Heart


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
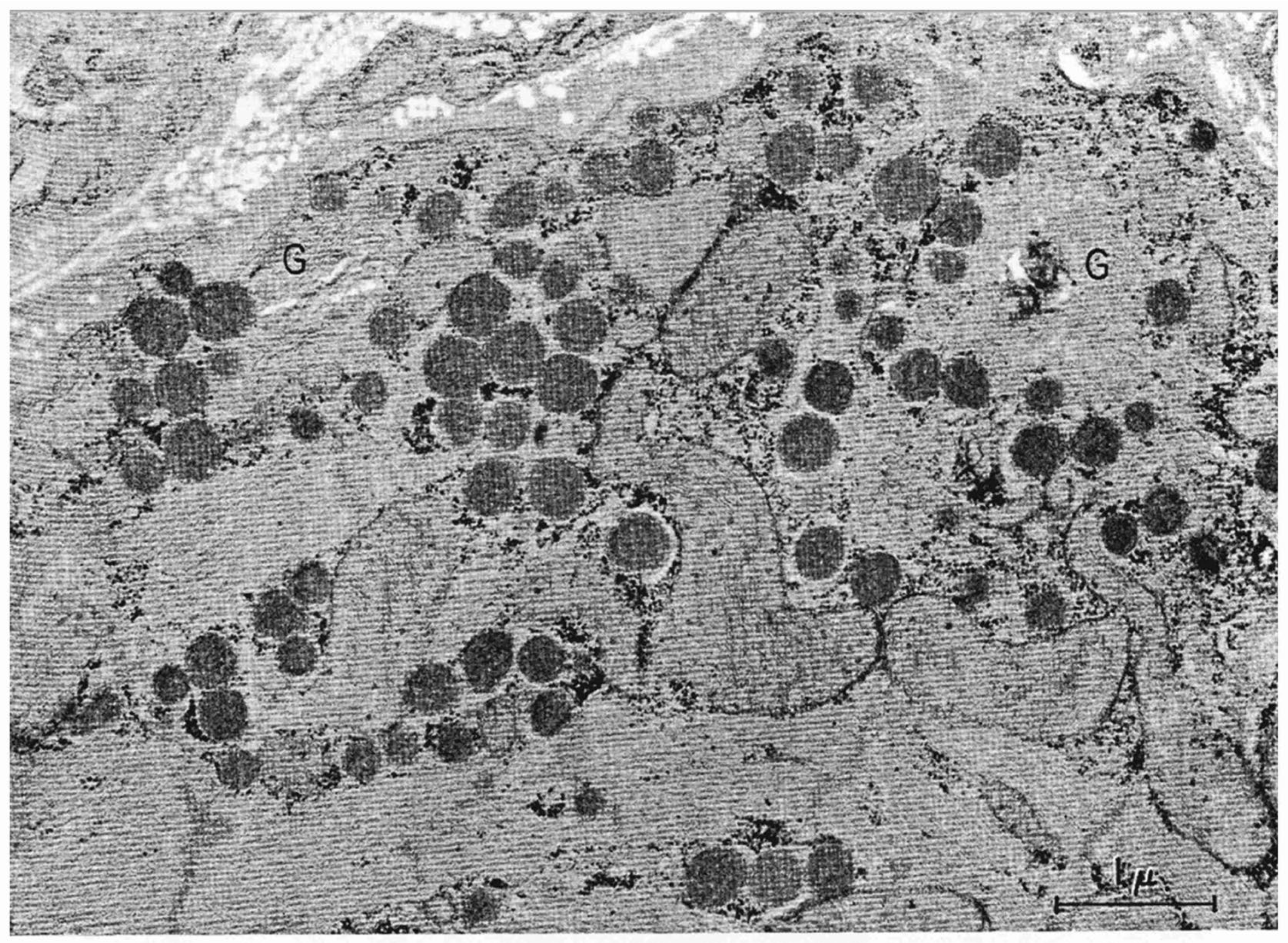
1.1. Granules
1.2. Translation
1.3. O-Glycosylation
1.4. Endoproteolysis
1.5. Other Modifications
2. Secretion
3. Natriuretic Peptides
4. Other Peptides
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Bold, A.J.; Borenstein, H.B.; Veress, A.T.; Sonnenberg, H. A rapid and potent natriuretic response to intravenous injection of atrial myocardial extract in rats. Life Sci. 1981, 28, 89–94. [Google Scholar] [CrossRef]
- Flynn, T.G.; de Bold, M.L.; de Bold, A.J. The amino acid sequence of an atrial peptide with potent diuretic and natriuretic properties. Biochem. Biophys. Res. Commun. 1983, 117, 859–865. [Google Scholar] [CrossRef]
- Kangawa, K.; Matsuo, H. Purification and complete amino acid sequence of alpha-human atrial natriuretic polypeptide (alpha-hANP). Biochem. Biophys. Res. Commun. 1984, 118, 131–139. [Google Scholar] [CrossRef]
- Sudoh, T.; Kangawa, K.; Minamino, N.; Matsuo, H. A new natriuretic peptide in porcine brain. Nature 1988, 332, 78–81. [Google Scholar] [CrossRef]
- Sudoh, T.; Minamino, N.; Kangawa, K.; Matsuo, H. C-type natriuretic peptide (CNP): A new member of natriuretic peptide family identified in porcine brain. Biochem. Biophys. Res. Commun. 1990, 168, 863–870. [Google Scholar] [CrossRef]
- Burnett, J.C., Jr.; Kao, P.C.; Hu, D.C.; Heser, D.W.; Heublein, D.; Granger, J.P.; Opgenorth, T.J.; Reeder, G.S. Atrial natriuretic peptide elevation in congestive heart failure in the human. Science 1986, 231, 1145–1147. [Google Scholar] [CrossRef]
- Daniels, L.B.; Maisel, A.S. Natriuretic peptides. J. Am. Coll. Cardiol. 2007, 50, 2357–2368. [Google Scholar] [CrossRef] [Green Version]
- Richards, A.M. N-terminal B-type natriuretic peptide in heart failure. Heart Fail. Clin. 2018, 14, 27–39. [Google Scholar] [CrossRef]
- Goetze, J.P.; Bruneau, B.G.; Ramos, H.R.; Ogawa, T.; de Bold, M.K.; de Bold, A.J. Cardiac natriuretic peptides. Nat. Rev. Cardiol. 2020, 17, 698–717. [Google Scholar] [CrossRef]
- Yan, W.; Wu, F.; Morser, J.; Wu, Q. Corin, a transmembrane cardiac serine protease, acts as a pro-atrial natriuretic peptide-converting enzyme. Proc. Natl. Acad. Sci. USA 2000, 97, 8525–8529. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Yan, W.; Pan, J.; Morser, J.; Wu, Q. Processing of pro-atrial natriuretic peptide by corin in cardiac myocytes. J. Biol. Chem. 2002, 277, 16900–16905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamieson, J.D.; Palade, G.E. Specific granules in atrial muscle cells. J. Cell. Biol. 1964, 23, 151–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, H.J.; Weiler, R.; Ludescher, C.; Schmid, K.W.; Winkler, H. Chromogranins A and B are co-localized with atrial natriuretic peptides in secretory granules of rat heart. J. Histochem. Cytochem. 1990, 38, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Pieroni, M.; Corti, A.; Tota, B.; Curnis, F.; Angelone, T.; Colombo, B.; Cerra, M.C.; Bellocci, F.; Crea, F.; Maseri, A. Myocardial production of chromogranin A in human heart: A new regulatory peptide of cardiac function. Eur. Heart J. 2007, 28, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Heidrich, F.M.; Zhang, K.; Estrada, M.; Huang, Y.; Giordano, F.J.; Ehrlich, B.E. Chromogranin B regulates calcium signaling, nuclear factor kappaB activity, and brain natriuretic peptide production in cardiomyocytes. Circ. Res. 2008, 102, 1230–1238. [Google Scholar] [CrossRef] [Green Version]
- Eipper, B.A.; May, V.; Braas, K.M. Membrane-associated peptidylglycine alpha-amidating monooxygenase in the heart. J. Biol. Chem. 1988, 263, 8371–8379. [Google Scholar] [CrossRef]
- Ouafik, L.; May, V.; Keutmann, H.T.; Eipper, B.A. Developmental regulation of peptidylglycine alpha-amidating monooxygenase (PAM) in rat heart atrium and ventricle. Tissue-specific changes in distribution of PAM activity, mRNA levels, and protein forms. J. Biol. Chem. 1989, 264, 5839–5845. [Google Scholar] [CrossRef]
- Powers, K.G.; Ma, X.M.; Eipper, B.A.; Mains, R.E. Identifying roles for peptidergic signaling in mice. Proc. Natl. Acad. Sci. USA 2019, 116, 20169–20179. [Google Scholar] [CrossRef] [Green Version]
- Bäck, N.; Luxmi, R.; Powers, K.G.; Mains, R.E.; Eipper, B.A. Peptidylglycine α-amidating monooxygenase is required for atrial secretory granule formation. Proc. Natl. Acad. Sci. USA 2020, 117, 17820–17831. [Google Scholar] [CrossRef]
- Bartels, E.D.; Goetze, J.P.; Mains, R.E.; Eipper, B.A. Commentary on: Peptidylglycine α-amidating monooxygenase is required for atrial secretory granule formation. J. Clin. Cardiol. 2021, 2, 75–80. [Google Scholar]
- Hasegawa, K.; Fujiwara, H.; Doyama, K.; Mukoyama, M.; Nakao, K.; Fujiwara, T.; Imura, H.; Kawai, C. Ventricular expression of atrial and brain natriuretic peptides in dilated cardiomyopathy. An immunohistocytochemical study of the endomyocardial biopsy specimens using specific monoclonal antibodies. Am. J. Pathol. 1993, 142, 107–116. [Google Scholar] [PubMed]
- Christoffersen, C.; Goetze, J.P.; Bartels, E.D.; Larsen, M.O.; Ribel, U.; Rehfeld, J.F.; Rolin, B.; Nielsen, L.B. Chamber-dependent expression of brain natriuretic peptide and its mRNA in normal and diabetic pig heart. Hypertension 2002, 40, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Takemura, G.; Takatsu, Y.; Doyama, K.; Itoh, H.; Saito, Y.; Koshiji, M.; Ando, F.; Fujiwara, T.; Nakao, K.; Fujiwara, H. Expression of atrial and brain natriuretic peptides and their genes in hearts of patients with cardiac amyloidosis. J. Am. Coll. Cardiol. 1998, 31, 754–765. [Google Scholar] [CrossRef] [Green Version]
- Goetze, J.P.; Johnsen, A.H.; Kistorp, C.; Gustafsson, F.; Johnbeck, C.B.; Rehfeld, J.F. Cardiomyocyte expression and cell-specific processing of procholecystokinin. J. Biol. Chem. 2015, 290, 6837–6843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetze, J.P.; Rehfeld, J.F. Procholecystokinin expression and processing in cardiac myocytes. Peptides 2019, 111, 71–76. [Google Scholar] [CrossRef]
- Siriwardena, M.; Kleffmann, T.; Ruygrok, P.; Cameron, V.A.; Yandle, T.G.; Nicholls, M.G.; Richards, A.M.; Pemberton, C.J. B-type natriuretic peptide signal peptide circulates in human blood: Evaluation as a potential biomarker of cardiac ischemia. Circulation 2010, 122, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Pemberton, C.J.; Siriwardena, M.; Kleffmann, T.; Ruygrok, P.; Palmer, S.C.; Yandle, T.G.; Richards, A.M. First identification of circulating prepro-A-type natriuretic peptide (preproANP) signal peptide fragments in humans: Initial assessment as cardiovascular biomarkers. Clin. Chem. 2012, 58, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Pemberton, C.J.; Siriwardena, M.; Kleffmann, T.; Richards, A.M. C-type natriuretic peptide (CNP) signal peptide fragments are present in the human circulation. Biochem. Biophys. Res. Commun. 2014, 449, 301–306. [Google Scholar] [CrossRef]
- Hunt, P.J.; Yandle, T.G.; Nicholls, M.G.; Richards, A.M.; Espiner, E.A. The amino-terminal portion of pro-brain natriuretic peptide (Pro-BNP) circulates in human plasma. Biochem. Biophys. Res. Commun. 1995, 214, 1175–1183. [Google Scholar] [CrossRef]
- Shimizu, H.; Masuta, K.; Aono, K.; Asada, H.; Sasakura, K.; Tamaki, M.; Sugita, K.; Yamada, K. Molecular forms of human brain natriuretic peptide in plasma. Clin. Chim. Acta 2002, 316, 129–135. [Google Scholar] [CrossRef]
- Goetze, J.P.; Kastrup, J.; Pedersen, F.; Rehfeld, J.F. Quantification of pro-B-type natriuretic peptide and its products in human plasma by use of an analysis independent of precursor processing. Clin. Chem. 2002, 48, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Schellenberger, U.; O’Rear, J.; Guzzetta, A.; Jue, R.A.; Protter, A.A.; Pollitt, N.S. The precursor to B-type natriuretic peptide is an O-linked glycoprotein. Arch. Biochem. Biophys. 2006, 45, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.H.; Madsen, T.D.; Goth, C.K.; Clausen, H.; Chen, Y.; Dzhoyashvili, N.; Iyer, S.R.; Sangaralingham, S.J.; Burnett, J.C., Jr.; Rehfeld, J.F.; et al. Discovery of O-glycans on atrial natriuretic peptide (ANP) that affect both its proteolytic degradation and potency at its cognate receptor. J. Biol. Chem. 2019, 294, 12567–12578. [Google Scholar] [CrossRef] [Green Version]
- Lewis, L.K.; Raudsepp, S.D.; Prickett, T.C.R.; Yandle, T.G.; Doughty, R.N.; Frampton, C.M.; Pemberton, C.J.; Richards, A.M. ProBNP that is not glycosylated at Threonine 71 is decreased with obesity in patients with heart failure. Clin. Chem. 2019, 65, 1115–1124. [Google Scholar] [CrossRef]
- Madsen, T.D.; Hansen, L.H.; Hintze, J.; Ye, Z.; Jebari, S.; Andersen, D.B.; Joshi, H.J.; Ju, T.; Goetze, J.P.; Martin, C.; et al. An atlas of O-linked glycosylation on peptide hormones reveals diverse biological roles. Nat. Commun. 2020, 11, 4033. [Google Scholar] [CrossRef] [PubMed]
- Amplatz, B.; Sarg, B.; Faserl, K.; Hammerer-Lercher, A.; Mair, J.; Lindner, H.H. Exposing the high heterogeneity of circulating pro B-type natriuretic peptide fragments in healthy individuals and heart failure patients. Clin. Chem. 2020, 66, 1200–1209. [Google Scholar] [CrossRef]
- Semenov, A.G.; Postnikov, A.B.; Tamm, N.N.; Seferian, K.R.; Karpova, N.S.; Bloshchitsyna, M.N.; Koshkina, E.V.; Krasnoselsky, M.I.; Serebryanaya, D.V.; Katrukha, A.G. Processing of pro-brain natriuretic peptide is suppressed by O-glycosylation in the region close to the cleavage site. Clin. Chem. 2009, 55, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Ichiki, T.; Huntley, B.K.; Heublein, D.M.; Sandberg, S.M.; McKie, P.M.; Martin, F.L.; Jougasaki, M.; Burnett, J.C., Jr. Corin is present in the normal human heart, kidney, and blood, with pro-B-type natriuretic peptide processing in the circulation. Clin. Chem. 2011, 57, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Suda, M.; Yokoyama, H.; Kanda, T.; Sakamaki, T.; Tanaka, S.; Nagai, R.; Abe, S.; Takeuchi, T. Stretch-induced hypertrophic growth of cardiocytes and processing of brain-type natriuretic peptide are controlled by proprotein-processing endoprotease furin. J. Biol. Chem. 1997, 272, 20545–20554. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.C.; Damholt, A.B.; Billestrup, N.; Kisbye, T.; Galante, P.; Michelsen, B.; Kofod, H.; Nielsen, J.H. Developmental expression of proprotein convertase 1/3 in the rat. Mol. Cell. Endocrinol. 1999, 155, 27–35. [Google Scholar] [CrossRef]
- Bloomquist, B.T.; Eipper, B.A.; Mains, R.E. Prohormone-converting enzymes: Regulation and evaluation of function using antisense RNA. Mol. Endocrinol. 1991, 5, 2014–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaubien, G.; Schäfer, M.K.; Weihe, E.; Dong, W.; Chrétien, M.; Seidah, N.G.; Day, R. The distinct gene expression of the pro-hormone convertases in the rat heart suggests potential substrates. Cell Tissue Res. 1995, 279, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Mishiro, E.; Sakakibara, Y.; Liu, M.C.; Suiko, M. Differential enzymatic characteristics and tissue-specific expression of human TPST-1 and TPST-2. J. Biochem. 2006, 140, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Dautzenberg, F.M.; Müller, D.; Richter, D. Dephosphorylation of phosphorylated atrial natriuretic peptide by protein phosphatase 2A. Eur. J. Biochem. 1993, 211, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Canaff, L.; Brechler, V.; Reudelhuber, T.L.; Thibault, G. Secretory granule targeting of atrial natriuretic peptide correlates with its calcium-mediated aggregation. Proc. Natl. Acad. Sci. USA 1996, 93, 9483–9487. [Google Scholar] [CrossRef] [Green Version]
- Baertschi, A.J.; Monnier, D.; Schmidt, U.; Levitan, E.S.; Fakan, S.; Roatti, A. Acid prohormone sequence determines size, shape, and docking of secretory vesicles in atrial myocytes. Circ. Res. 2001, 89, E23–E29. [Google Scholar] [CrossRef] [Green Version]
- John, S.W.; Krege, J.H.; Oliver, P.M.; Hagaman, J.R.; Hodgin, J.B.; Pang, S.C.; Flynn, T.G.; Smithies, O. Genetic decreases in atrial natriuretic peptide and salt-sensitive hypertension. Science 1995, 267, 679–681. [Google Scholar] [CrossRef]
- Masters, R.G.; Davies, R.A.; Veinot, J.P.; Hendrym, P.J.; Smith, S.J.; de Bold, A.J. Discoordinate modulation of natriuretic peptides during acute cardiac allograft rejection in humans. Circulation 1999, 100, 287–291. [Google Scholar] [CrossRef] [Green Version]
- de Bold, A.J. Determinants of brain natriuretic peptide gene expression and secretion in acute cardiac allograft rejection. Curr. Opin. Cardiol. 2007, 22, 146–150. [Google Scholar] [CrossRef]
- Zois, N.E.; Bartels, E.D.; Hunter, I.; Kousholt, B.S.; Olsen, L.H.; Goetze, J.P. Natriuretic peptides in cardiometabolic regulation and disease. Nat. Rev. Cardiol. 2014, 11, 403–412. [Google Scholar] [CrossRef]
- Goetze, J.P.; Kastrup, J.; Rehfeld, J.F. The paradox of increased natriuretic hormones in congestive heart failure patients: Does the endocrine heart also fail in heart failure? Eur. Heart J. 2003, 24, 1471–1472. [Google Scholar] [CrossRef]
- Burger, A.J.; Burger, M.R. Nesiritide: Past, present, and future. Minerva Cardioangiol. 2005, 53, 509–522. [Google Scholar] [PubMed]
- Chen, M.M.; Ashley, E.A.; Deng, D.X.; Tsalenko, A.; Deng, A.; Tabibiazar, R.; Ben-Dor, A.; Fenster, B.; Yang, E.; King, J.Y.; et al. Novel role for the potent endogenous inotrope apelin in human cardiac dysfunction. Circulation 2003, 108, 1432–1439. [Google Scholar] [CrossRef]
- Parikh, M.; Shah, S.; Basu, R.; Famulski, K.S.; Kim, D.; Mullen, J.C.; Halloran, P.F.; Oudit, G.Y. Transcriptomic signatures of end-stage human dilated cardiomyopathy hearts with and without left ventricular assist device support. Int. J. Mol. Sci. 2022, 23, 2050. [Google Scholar] [CrossRef] [PubMed]
- Chong, K.S.; Gardner, R.S.; Morton, J.J.; Ashley, E.A.; McDonagh, T.A. Plasma concentrations of the novel peptide apelin are decreased in patients with chronic heart failure. Eur. J. Heart Fail. 2006, 8, 355–360. [Google Scholar] [CrossRef]
- Goetze, J.P.; Rehfeld, J.F.; Carlsen, J.; Videbaek, R.; Andersen, C.B.; Boesgaard, S.; Friis-Hansen, L. Apelin: A new plasma marker of cardiopulmonary disease. Regul. Pept. 2006, 133, 134–138. [Google Scholar] [CrossRef]
- Sans-Roselló, J.; Casals, G.; Rossello, X.; de la Presa, B.G.; Vila, M.; Duran-Cambra, A.; Morales-Ruiz, M.; Ferrero-Gregori, A.; Jiménez, W.; Sionis, A. Prognostic value of plasma apelin concentrations at admission in patients with ST-segment elevation acute myocardial infarction. Clin. Biochem. 2017, 50, 279–284. [Google Scholar] [CrossRef]
- Chandrasekaran, B.; Kalra, P.R.; Donovan, J.; Hooper, J.; Clague, J.R.; McDonagh, T.A. Myocardial apelin production is reduced in humans with left ventricular systolic dysfunction. J. Card. Fail. 2010, 16, 556–561. [Google Scholar] [CrossRef]
- Helske, S.; Kovanen, P.T.; Lommi, J.; Turto, H.; Kupari, M. Transcardiac gradients of circulating apelin: Extraction by normal hearts vs. release by hearts failing due to pressure overload. J. Appl. Physiol. 2010, 109, 1744–1748. [Google Scholar] [CrossRef]
- Hisaw, F.L. Experimental relaxation of the pubic ligament of the guinea pig. Proc. Soc. Exp. Biol. Med. 1926, 23, 661–663. [Google Scholar] [CrossRef]
- Bathgate, R.A.; Halls, M.L.; van der Westhuizen, E.T.; Callander, G.E.; Kocan, M.; Summers, R.J. Relaxin family peptides and their receptors. Physiol. Rev. 2013, 93, 405–480. [Google Scholar] [CrossRef]
- Dschietzig, T.; Richter, C.; Bartsch, C.; Laule, M.; Armbruster, F.P.; Baumann, G.; Stangl, K. The pregnancy hormone relaxin is a player in human heart failure. FASEB J. 2001, 15, 2187–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupari, M.; Mikkola, T.S.; Turto, H.; Lommi, J. Is the pregnancy hormone relaxin an important player in human heart failure? Eur. J. Heart Fail. 2005, 7, 195–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bani, D.; Masini, E.; Bello, M.G.; Bigazzi, M.; Sacchi, T.B. Relaxin protects against myocardial injury caused by ischemia and reperfusion in rat heart. Am. J. Pathol. 1998, 152, 1367–1376. [Google Scholar]
- Fisher, C.; Berry, C.; Blue, L.; Morton, J.J.; McMurray, J. N-terminal pro B type natriuretic peptide, but not the new putative cardiac hormone relaxin, predicts prognosis in patients with chronic heart failure. Heart 2003, 89, 879–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metra, M.; Teerlink, J.R.; Cotter, G.; Davison, B.A.; Felker, G.M.; Filippatos, G.; Greenberg, B.H.; Pang, P.S.; Ponikowski, P.; Voors, A.A.; et al. Effects of serelaxin in patients with acute heart failure. N. Engl. J. Med. 2019, 381, 716–726. [Google Scholar] [CrossRef]
- Gibril, F.; Curtis, L.T.; Termanini, B.; Fritsch, M.K.; Lubensky, I.A.; Doppman, J.L.; Jensen, R.T. Primary cardiac gastrinoma causing Zollinger-Ellison syndrome. Gastroenterology 1997, 112, 567–574. [Google Scholar] [CrossRef]
- Lindstedt, G.; Bengtsson, C.; Lapidus, L.; Nyström, E. Hypergastrinemia–a risk factor for myocardial infarction? Clin. Chem. 1985, 31, 585–590. [Google Scholar] [CrossRef]
- Goetze, J.P.; Hunter, I.; Zois, N.E.; Terzic, D.; Valeur, N.; Olsen, L.H.; Smith, J.; Plomgaard, P.; Hansen, L.H.; Rehfeld, J.F.; et al. Cardiac procholecystokinin expression during haemodynamic changes in the mammalian heart. Peptides 2018, 108, 7–13. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goetze, J.P.; Bartels, E.D.; Shalmi, T.W.; Andraud-Dang, L.; Rehfeld, J.F. Biochemistry of the Endocrine Heart. Biology 2022, 11, 971. https://doi.org/10.3390/biology11070971
Goetze JP, Bartels ED, Shalmi TW, Andraud-Dang L, Rehfeld JF. Biochemistry of the Endocrine Heart. Biology. 2022; 11(7):971. https://doi.org/10.3390/biology11070971
Chicago/Turabian StyleGoetze, Jens P., Emil D. Bartels, Theodor W. Shalmi, Lilian Andraud-Dang, and Jens F. Rehfeld. 2022. "Biochemistry of the Endocrine Heart" Biology 11, no. 7: 971. https://doi.org/10.3390/biology11070971
APA StyleGoetze, J. P., Bartels, E. D., Shalmi, T. W., Andraud-Dang, L., & Rehfeld, J. F. (2022). Biochemistry of the Endocrine Heart. Biology, 11(7), 971. https://doi.org/10.3390/biology11070971