
High-Temperature Stress Effect on the Red Cusk-Eel (Geypterus chilensis) Liver: Transcriptional Modulation and Oxidative Stress Damage

 ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish Sampling and Experimental Design
2.3. AST, ALT and AP Evaluation
2.4. Oxidative Stress Assays in Liver
2.5. Liver RNA Extraction, Library Preparation and Illumina Sequencing
2.6. Reads Filtering, Differential Expression, and GO Enrichment Analysis
2.7. RNA-seq Validation by qPCR
2.8. Statistical Analysis
3. Results
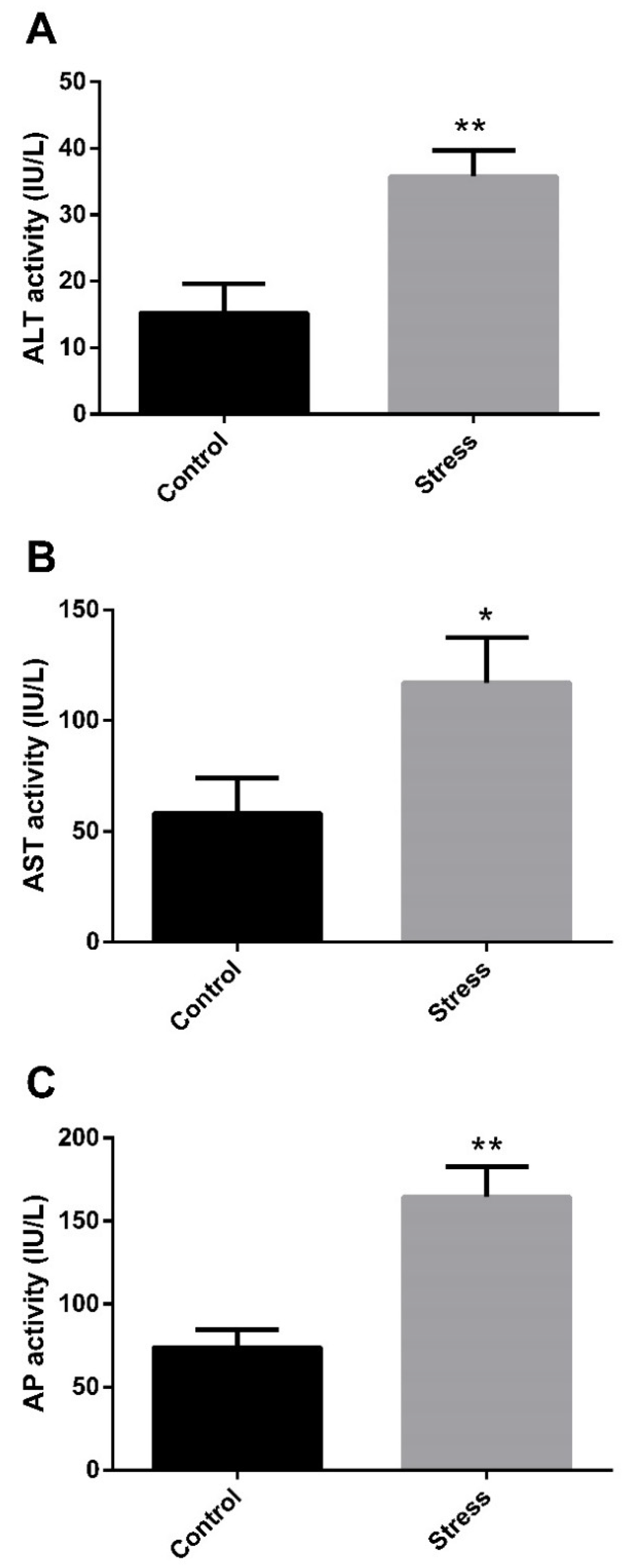
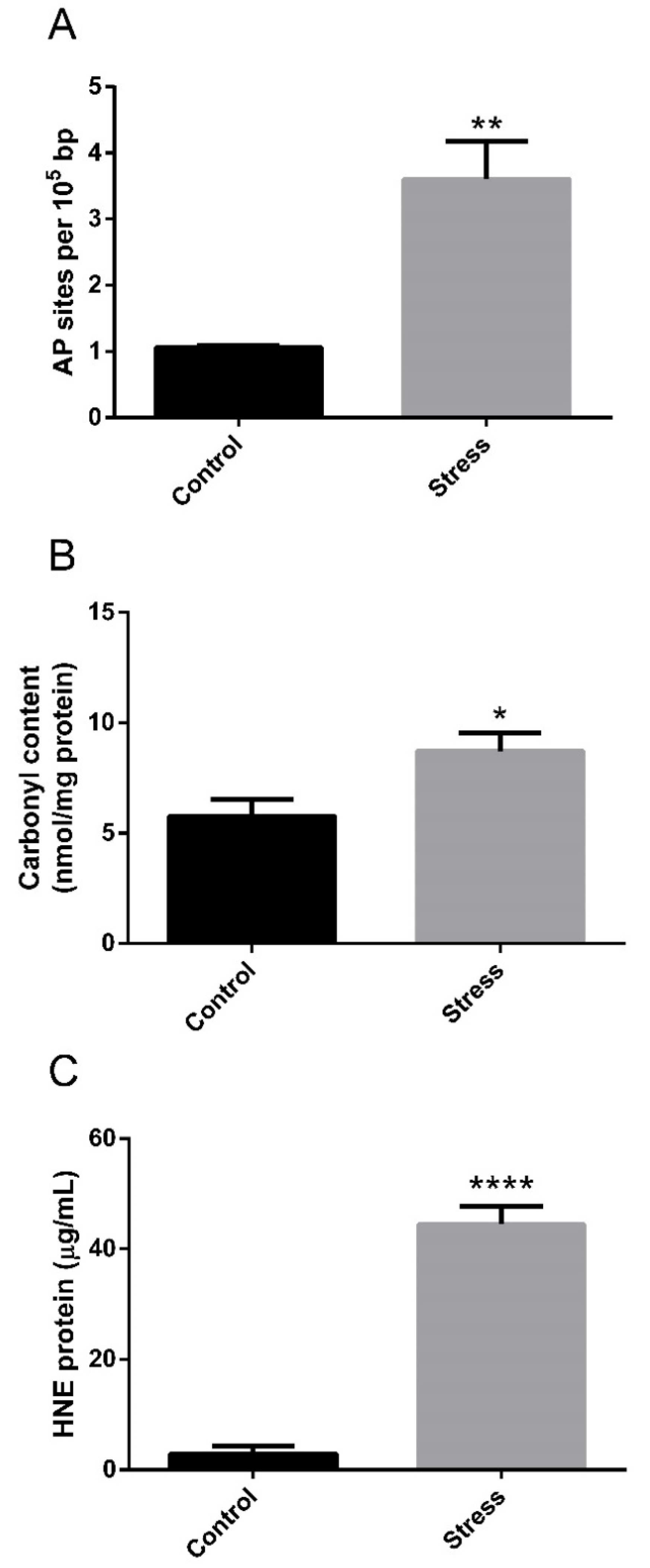
3.1. Hepatic Enzyme Activity and Oxidative Stress Response to High-Temperature Stress
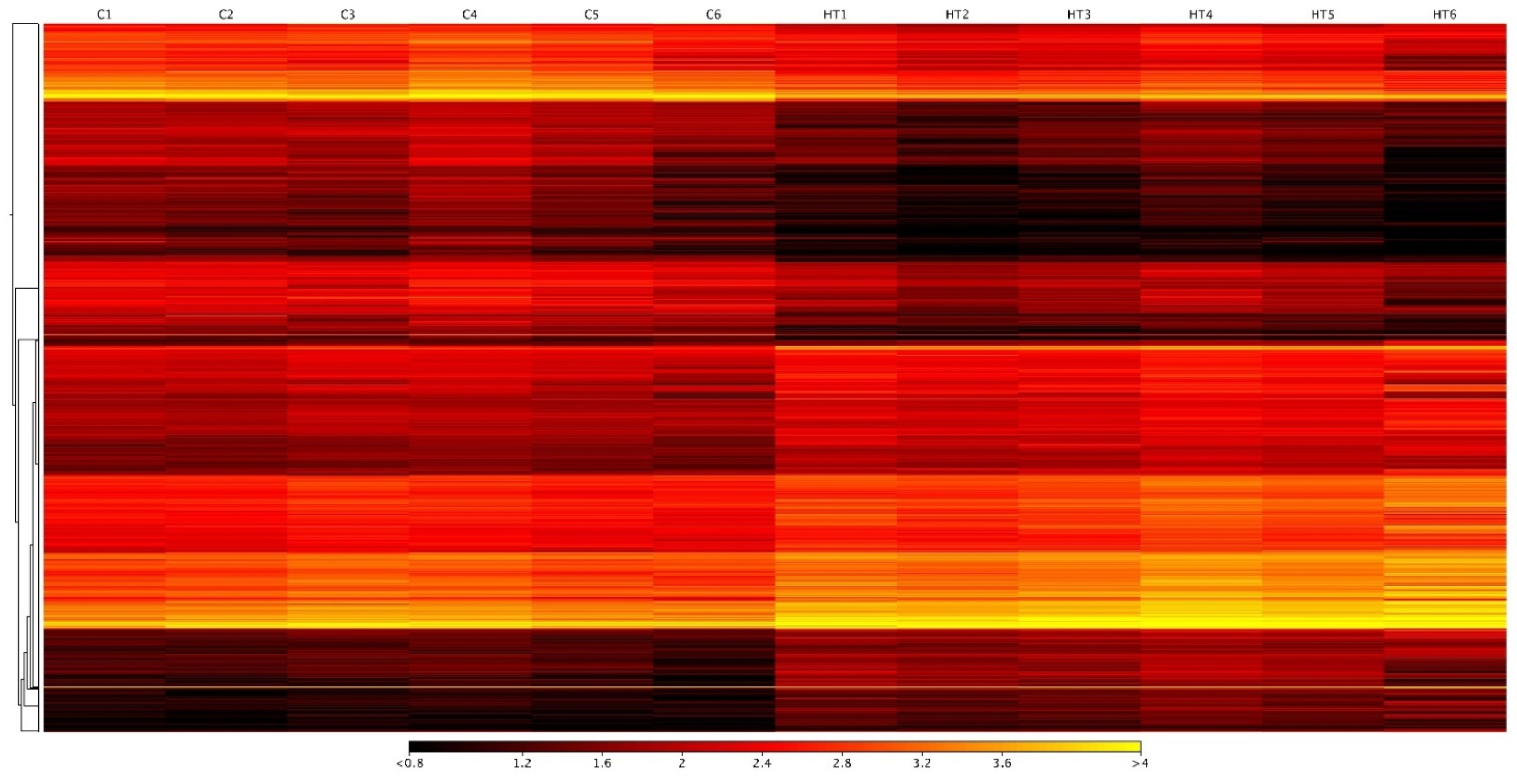
3.2. Differentially Expressed Transcripts in Hepatic Response to High-Temperature Stress
3.3. GO Enrichment and Pathway Analysis in the Liver
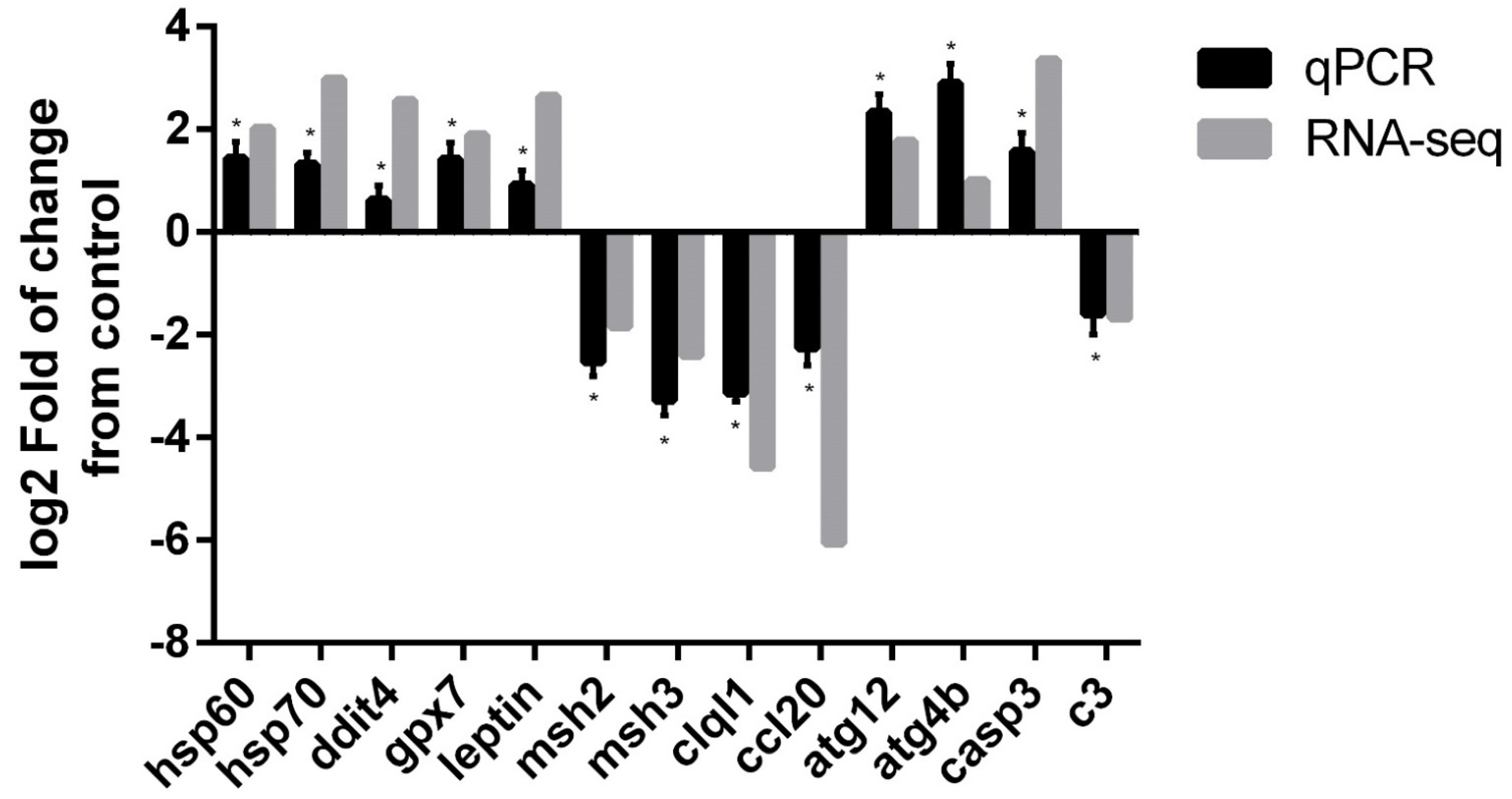
3.4. qPCR Validation of Differentially Expressed Genes
4. Discussion
4.1. High-Temperature Effect on Hepatic Enzymes
4.2. Oxidative Stress under High-Temperature Stress
4.3. High-Temperature Stress in Hepatic Protein Processing and Folding
4.4. Heat Shock Protein as Thermal Stress Biomarkers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Somero, G.N. The physiology of climate change: How potentials for acclimatization and genetic adaptation will determine ‘winners’ and ‘losers’. J. Exp. Biol. 2010, 213, 912–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Li, Y.; Liu, Z.; Kang, Y.; Wang, J. Transcriptomic responses to heat stress in rainbow trout Oncorhynchus mykiss head kidney. Fish Shellfish Immunol. 2018, 82, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, A.; Gunther, O.P.; Houde, A.L.; Li, S.; Ming, T.J.; Jeffries, K.M.; Hinch, S.G.; Miller, K.M. Developing specific molecular biomarkers for thermal stress in salmonids. BMC Genom. 2018, 19, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalvi, R.S.; Pal, A.K.; Tiwari, L.R.; Baruah, K. Influence of acclimation temperature on the induction of heat-shock protein 70 in the catfish Horabagrus brachysoma (Gunther). Fish Physiol. Biochem. 2012, 38, 919–927. [Google Scholar] [CrossRef]
- Aursnes, I.A.; Rishovd, A.L.; Karlsen, H.E.; Gjoen, T. Validation of reference genes for quantitative RT-qPCR studies of gene expression in Atlantic cod (Gadus morhua L.) during temperature stress. BMC Res. Notes 2011, 4, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, D.; Vinagre, C.; Costa, P.M.; Diniz, M.S. Histopathological alterations, physiological limits, and molecular changes of juvenile Sparus aurata in response to thermal stress. Mar. Ecol. Prog. Ser. 2014, 505, 256–266. [Google Scholar] [CrossRef]
- Dobretsov, S.; Coutinho, R.; Rittschof, D.; Salta, M.; Ragazzola, F.; Hellio, C. The oceans are changing: Impact of ocean warming and acidification on biofouling communities. Biofouling 2019, 35, 585–595. [Google Scholar] [CrossRef]
- Wang, B.; Luo, X.; Yang, Y.M.; Sun, W.; Cane, M.A.; Cai, W.; Yeh, S.W.; Liu, J. Historical change of El Nino properties sheds light on future changes of extreme El Nino. Proc. Natl. Acad. Sci. USA 2019, 116, 22512–22517. [Google Scholar] [CrossRef] [Green Version]
- Flores-Kossack, C.; Montero, R.; Kollner, B.; Maisey, K. Chilean aquaculture and the new challenges: Pathogens, immune response, vaccination and fish diversification. Fish Shellfish Immunol. 2020, 98, 52–67. [Google Scholar] [CrossRef]
- Vega, R.; Pradenas, M.; Estrada, J.M.; Ramirez, D.; Valdebenito, I.; Mardones, A.; Dantagnan, P.; Alfaro, D.; Encina, F.; Pichara, C. Evaluation and comparison of the efficiency of two incubation systems for Genypterus chilensis (Guichenot, 1848) eggs. Lat. Am. J. Aquat. Res. 2012, 40, 187–200. [Google Scholar] [CrossRef]
- Chong, J.; Gonzalez, P. Reproductive cycle and maturity mean size of the red cusk eel, Genypterus chilensis (Guichenot, 1881) in the coast off Talcahuano, Chile. Rev. Biol. Mar. Oceanogr. 2009, 44, 257–262. [Google Scholar] [CrossRef]
- Gonzalez, P.; Dettleff, P.; Valenzuela, C.; Estrada, J.M.; Valdes, J.A.; Meneses, C.; Molina, A. Evaluating the genetic structure of wild and commercial red cusk-eel (Genypterus chilensis) populations through the development of novel microsatellite markers from a reference transcriptome. Mol. Biol. Rep. 2019, 46, 5875–5882. [Google Scholar] [CrossRef] [PubMed]
- Vega, R.; Estrada, J.M.; Ramirez, D.; Flores, C.; Zamorano, J.; Encina, F.; Mardones, A.; Valdebenito, I.; Dantagnan, P. Growth of cusk eel Genypterus chilensis juveniles in culture conditions. Lat. Am. J. Aquat. Res. 2015, 43, 344–350. [Google Scholar] [CrossRef]
- SERNAPESCA. Anuario Estadístico de Pesca y Acuicultura 2020; Ministeriode Economía: Santiago, Chile, 2021. [Google Scholar]
- Aluru, N.; Vijayan, M.M. Stress transcriptomics in fish: A role for genomic cortisol signaling. Gen. Comp. Endocrinol. 2009, 164, 142–150. [Google Scholar] [CrossRef]
- Ellis, T.; Yildiz, H.Y.; Lopez-Olmeda, J.; Spedicato, M.T.; Tort, L.; Overli, O.; Martins, C.I.M. Cortisol and finfish welfare. Fish Physiol. Biochem. 2012, 38, 163–188. [Google Scholar] [CrossRef]
- Faught, E.; Aluru, N.; Vijayan, M. The molecular stress response. In Biology of Stress in Fish; Schreck, C.B., Tort, L., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 35, pp. 114–149. [Google Scholar]
- Logan, C.A.; Somero, G.N. Effects of thermal acclimation on transcriptional responses to acute heat stress in the eurythermal fish Gillichthys mirabilis (Cooper). Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 300, R1373–R1383. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.M.; Liu, J.H.; Shu, L.H.; Chen, C.H. Anti-oxidative responses of zebrafish (Danio rerio) gill, liver and brain tissues upon acute cold shock. Comp. Biochem. Phys. A 2015, 187, 202–213. [Google Scholar] [CrossRef]
- Yang, C.G.; Jiang, M.; Wen, H.; Tian, J.; Liu, W.; Wu, F.; Gou, G.W. Analysis of differential gene expression under low-temperature stress in Nile tilapia (Oreochromis niloticus) using digital gene expression. Gene 2015, 564, 134–140. [Google Scholar] [CrossRef]
- Nakano, T.; Kameda, M.; Shoji, Y.; Hayashi, S.; Yamaguchi, T.; Sato, M. Effect of severe environmental thermal stress on redox state in salmon. Redox Biol. 2014, 2, 772–776. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, Z.; Li, Z.; Shi, H.; Kang, Y.; Wang, J.; Huang, J.; Jiang, L. Effects of heat stress on respiratory burst, oxidative damage and SERPINH1 (HSP47) mRNA expression in rainbow trout Oncorhynchus mykiss. Fish Physiol. Biochem. 2016, 42, 701–710. [Google Scholar] [CrossRef]
- Dettleff, P.; Zuloaga, R.; Fuentes, M.; Estrada, J.M.; Molina, A.; Valdes, J.A. Temperature effect on oxidative stress and egg quality-related genes on post-ovulatory eggs and ovary of red cusk-eel (Genypterus chilensis). J. Fish Biol. 2021, 98, 1475–1480. [Google Scholar] [CrossRef] [PubMed]
- Dettleff, P.; Zuloaga, R.; Fuentes, M.; González, J.; Aedo, J.; Estrada, J.M.; Molina, A.; Valdés, J.A. Physiological and molecular responses to thermal stress in red cusk-eel (Genypterus chilensis) juveniles reveals atrophy and oxidative damage in skeletal muscle. J. Therm. Biol. 2020, 94, 102750. [Google Scholar] [CrossRef] [PubMed]
- Naour, S.; Espinoza, B.M.; Aedo, J.E.; Zuloaga, R.; Maldonado, J.; Bastias-Molina, M.; Silva, H.; Meneses, C.; Gallardo-Escarate, C.; Molina, A.; et al. Transcriptomic analysis of the hepatic response to stress in the red cusk-eel (Genypterus chilensis): Insights into lipid metabolism, oxidative stress and liver steatosis. PLoS ONE 2017, 12, e0176447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dettleff, P.; Hormazabal, E.; Aedo, J.; Fuentes, M.; Meneses, C.; Molina, A.; Valdes, J.A. Identification and Evaluation of Long Noncoding RNAs in Response to Handling Stress in Red Cusk-Eel (Genypterus chilensis) via RNA-seq. Mar. Biotechnol. 2020, 22, 94–108. [Google Scholar] [CrossRef] [PubMed]
- SHOA. Servicio Hidrográfico y Oceanográfico de la Armada de Chile 2019; Servicio Hidrografico y Oceanografico de la Armada: Valparaiso, Chile, 2019. [Google Scholar]
- Aedo, J.E.; Maldonado, J.; Estrada, J.M.; Fuentes, E.N.; Silva, H.; Gallardo-Escarate, C.; Molina, A.; Valdes, J.A. Sequencing and de novo assembly of the red cusk-eel (Genypterus chilensis) transcriptome. Mar. Genom. 2014, 18, 105–107. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean acidification: The other CO2 problem. Annu. Rev. Mar. Sci. 2009, 1, 169–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alimi, O.S.; Farner Budarz, J.; Hernandez, L.M.; Tufenkji, N. Microplastics and Nanoplastics in Aquatic Environments: Aggregation, Deposition and Enhanced Contaminant Transport. Environ. Sci. Technol. 2018, 52, 1704–1724. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, J.; Nishimura, N.; Shimada, Y. Toxicological interactions of microplastics/nanoplastics and environmental contaminants: Current knowledge and future perspectives. J. Hazard. Mater. 2021, 405, 123913. [Google Scholar] [CrossRef] [PubMed]
- Aballai, V.; Aedo, J.E.; Maldonado, J.; Bastias-Molina, M.; Silva, H.; Meneses, C.; Boltana, S.; Reyes, A.; Molina, A.; Valdes, J.A. RNA-seq analysis of the head-kidney transcriptome response to handling-stress in the red cusk-eel (Genypterus chilensis). Comp. Biochem. Phys. D 2017, 24, 111–117. [Google Scholar] [CrossRef]
- Aedo, J.E.; Maldonado, J.; Aballai, V.; Estrada, J.M.; Bastias-Molina, M.; Meneses, C.; Gallardo-Escarate, C.; Silva, H.; Molina, A.; Valdes, J.A. mRNA-seq reveals skeletal muscle atrophy in response to handling stress in a marine teleost, the red cusk-eel (Genypterus chilensis). BMC Genom. 2015, 16, 1024. [Google Scholar] [CrossRef] [Green Version]
- Pakhira, C.; Nagesh, T.S.; Abraham, T.J.; Dash, G.; Behera, S. Stress responses in rohu, Labeo rohita transported at different densities. Aquac. Rep. 2015, 2, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Krishnani, K.K.; Singh, N.P. Oxidative and Cellular Metabolic Stress of Fish: An Appealing Tool for Biomonitoring of Metal Contamination in the Kolkata Wetland, a Ramsar Site. Arch. Environ. Contam. Toxicol. 2019, 76, 469–482. [Google Scholar] [CrossRef]
- Veisi, S.; Sarkheil, M.; Johari, S.A.; Safari, O. Dietary supplementation with melatonin: Influence on growth performance, oxidative stress status, and amelioration of silver nanoparticles-induced toxicity in Nile tilapia (Oreochromis niloticus). Trop. Anim. Health Prod. 2021, 53, 314. [Google Scholar] [CrossRef]
- Firat, O.; Tutus, R. Comparative Acute Toxicity Assessment of Organophosphate and Avermectin Insecticides on a Freshwater Fish Oreochromis niloticus. Bull. Environ. Contam. Toxicol. 2020, 105, 582–587. [Google Scholar] [CrossRef]
- Bharti, S.; Rasool, F. Analysis of the biochemical and histopathological impact of a mild dose of commercial malathion on Channa punctatus (Bloch) fish. Toxicol. Rep. 2021, 8, 443–455. [Google Scholar] [CrossRef]
- Cheng, C.H.; Guo, Z.X.; Luo, S.W.; Wang, A.L. Effects of high temperature on biochemical parameters, oxidative stress, DNA damage and apoptosis of pufferfish (Takifugu obscurus). Ecotoxicol. Environ. Saf. 2018, 150, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhao, T.; Ma, A.; Huang, Z.; Liu, Z.; Cui, W.; Zhang, J.; Zhu, C.; Guo, X.; Yuan, C. Metabolic responses in Scophthalmus maximus kidney subjected to thermal stress. Fish Shellfish Immunol. 2020, 103, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Das, A. Heat stress-induced hepatotoxicity and its prevention by resveratrol in rats. Toxicol. Mech. Methods 2011, 21, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Betteridge, D.J. What is oxidative stress? Metab. Clin. Exp. 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Madeira, D.; Narciso, L.; Cabral, H.N.; Vinagre, C.; Diniz, M.S. Influence of temperature in thermal and oxidative stress responses in estuarine fish. Comparative Biochemistry and Physiology. Part A Mol. Integr. Physiol. 2013, 166, 237–243. [Google Scholar] [CrossRef]
- Vinagre, C.; Madeira, D.; Narciso, L.; Cabral, H.N.; Diniz, M.S. Effect of temperature on oxidative stress in fish: Lipid peroxidation and catalase activity in the muscle of juvenile seabass, Dicentrarchus labrax. Ecol. Indic. 2012, 23, 274–279. [Google Scholar] [CrossRef]
- Chang, C.H.; Mayer, M.; Rivera-Ingraham, G.; Blondeau-Bidet, E.; Wu, W.Y.; Lorin-Nebel, C.; Lee, T.H. Effects of temperature and salinity on antioxidant responses in livers of temperate (Dicentrarchus labrax) and tropical (Chanos Chanos) marine euryhaline fish. J. Therm. Biol. 2021, 99, 103016. [Google Scholar] [CrossRef]
- Ale, A.; Bacchetta, C.; Rossi, A.S.; Scarabotti, P.A.; Cazenave, J. Low temperature stress in a cultured fish (Piaractus mesopotamicus) fed with Pyropia columbina red seaweed-supplemented diet. Fish Physiol. Biochem. 2021, 47, 829–839. [Google Scholar] [CrossRef]
- Bacchetta, C.; Ale, A.; Rossi, A.S.; Karakachoff, M.; Cazenave, J. Effects of cold stress on juvenile Piaractus mesopotamicus and the mitigation by beta-carotene. J. Therm. Biol. 2020, 88, 102497. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.; Perez-Jimenez, A.; Guerreiro, I.; Peres, H.; Castro-Cunha, M.; Oliva-Teles, A. Effects of temperature and dietary protein level on hepatic oxidative status of Senegalese sole juveniles (Solea senegalensis). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 163, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Roychowdhury, P.; Aftabuddin, M.; Pati, M.K. Thermal stress-induced oxidative damages in the liver and associated death in fish, Labeo rohita. Fish Physiol. Biochem. 2021, 47, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Anido, R.N.; Harrington, A.M.; Hamlin, H.J. Coping with stress in a warming Gulf: The postlarval American lobster’s cellular stress response under future warming scenarios. Cell Stress Chaperones 2021, 26, 721–734. [Google Scholar] [CrossRef]
- An, K.W.; Kim, N.N.; Shin, H.S.; Kil, G.S.; Choi, C.Y. Profiles of antioxidant gene expression and physiological changes by thermal and hypoosmotic stresses in black porgy (Acanthopagrus schlegeli). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 262–268. [Google Scholar] [CrossRef]
- Cheng, C.H.; Yang, F.F.; Liao, S.A.; Miao, Y.T.; Ye, C.X.; Wang, A.L.; Tan, J.W.; Chen, X.Y. High temperature induces apoptosis and oxidative stress in pufferfish (Takifugu obscurus) blood cells. J. Therm. Biol. 2015, 53, 172–179. [Google Scholar] [CrossRef]
- Ho, T.N.; Paul, G.V.; Chen, Y.H.; Hsu, T. Heat stress upregulates G-T mismatch binding activities in zebrafish (Danio rerio) embryos preexposed and nonexposed to a sublethal level of cadmium (Cd). Chemosphere 2019, 218, 179–188. [Google Scholar] [CrossRef]
- Baker, B.P.; Van Wie, I.; Braun, E.; Jimenez, A.G. Thermal stability vs. variability: Insights in oxidative stress from a eurytolerant fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 249, 110767. [Google Scholar] [CrossRef]
- Anelli, T.; Sitia, R. Protein quality control in the early secretory pathway. EMBO J. 2008, 27, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Malini, M.K.; Lekshmy, V.S.; Pal, M.; Chinnusamy, V.; Nagaraj Kumar, M. Unfolded protein response (UPR) mediated under heat stress in plants. Plant Physiol. Rep. 2020, 25, 569–582. [Google Scholar] [CrossRef]
- Faoro, C.; Ataide, S.F. Noncanonical Functions and Cellular Dynamics of the Mammalian Signal Recognition Particle Components. Front. Mol. Biosci. 2021, 8, 679584. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.H.; Ron, D. Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature 1999, 397, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Vattem, K.M.; Wek, R.C. Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc. Natl. Acad. Sci. USA 2004, 101, 11269–11274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, M.; Liu, Y.; Fujii, K.; Calderwood, S.K.; Nakai, A.; Imai, K.; Shinomura, Y. Oxidative Stress Impairs the Heat Stress Response and Delays Unfolded Protein Recovery. PLoS ONE 2009, 4, A57–A66. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, K.; Zaidi, S.F.; Mati-ur-Rehman; Rehman, R.; Kondo, T. Hyperthermia and protein homeostasis: Cytoprotection and cell death. J. Therm. Biol. 2020, 91, 102615. [Google Scholar] [CrossRef]
- Fe-Goncalves, L.M.; Araujo, J.D.A.; dos Santos, C.H.D.; Val, A.L.; de Almeida-Val, V.M.F. How will farmed populations of freshwater fish deal with the extreme climate scenario in 2100? Transcriptional responses of Colossoma macropomum from two Brazilian climate regions. J. Therm. Biol. 2020, 89, 102487. [Google Scholar] [CrossRef]
- Mininni, A.N.; Milan, M.; Ferraresso, S.; Petochi, T.; Di Marco, P.; Marino, G.; Livi, S.; Romualdi, C.; Bargelloni, L.; Patarnello, T. Liver transcriptome analysis in gilthead sea bream upon exposure to low temperature. BMC Genom. 2014, 15, 765. [Google Scholar] [CrossRef] [Green Version]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Dammark, K.B.; Ferchaud, A.L.; Hansen, M.M.; Sorensen, J.G. Heat tolerance and gene expression responses to heat stress in threespine sticklebacks from ecologically divergent environments. J. Therm. Biol. 2018, 75, 88–96. [Google Scholar] [CrossRef]
- Corey, E.; Linnansaari, T.; Cunjak, R.A.; Currie, S. Physiological effects of environmentally relevant, multi-day thermal stress on wild juvenile Atlantic salmon (Salmo salar). Conserv. Physiol. 2017, 5, cox014. [Google Scholar] [CrossRef]
- Hori, T.S.; Gamperl, A.K.; Afonso, L.O.; Johnson, S.C.; Hubert, S.; Kimball, J.; Bowman, S.; Rise, M.L. Heat-shock responsive genes identified and validated in Atlantic cod (Gadus morhua) liver, head kidney and skeletal muscle using genomic techniques. BMC Genom. 2010, 11, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Xu, P.; Brown, P.B.; Xie, J.; Ge, X.P.; Miao, L.H.; Zhou, Q.L.; Ren, M.C.; Pan, L.K. The effect of hyperthermia on liver histology, oxidative stress and disease resistance of the Wuchang bream, Megalobrama amblycephala. Fish Shellfish Immunol. 2016, 52, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Nakai, A.; Satoh, M.; Hirayoshi, K.; Nagata, K. Involvement of the stress protein HSP47 in procollagen processing in the endoplasmic reticulum. J. Cell Biol. 1992, 117, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Rajesh, M.; Baral, P.; Sarma, D.; Tripathi, P.H.; Akhtar, M.S.; Ciji, A.; Dubey, M.K.; Pande, V.; Sharma, P.; et al. Concurrent changes in thermal tolerance thresholds and cellular heat stress response reveals novel molecular signatures and markers of high temperature acclimation in rainbow trout. J. Therm. Biol. 2021, 102, 103124. [Google Scholar] [CrossRef]
- Long, Y.; Li, L.; Li, Q.; He, X.; Cui, Z. Transcriptomic characterization of temperature stress responses in larval zebrafish. PLoS ONE 2012, 7, e37209. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Gene Name | Forward Sequence | Reverse Sequence | Amplicon Size (bp) | Tm | Efficiency (%) |
|---|---|---|---|---|---|---|
| taf12 | Transcription initiation factor TFIID subunit 12 | GATCTGTAACGACGACGAAGAA | CAAATCAGAGGGACGTCATGTA | 92 | 62 | 101 |
| actb | Beta actin | TGTCCCTGTATGCTTCTGGT | CCCCTCTCAGTCAGAATCTTCAT | 172 | 62 | 104 |
| hsp60 | Heat shock protein 60 | GACGGTTCCAATCTCTACATCTC | CGCTCTCCAAACCAGTTACA | 86 | 62 | 99 |
| hsp70 | Heat shock protein 70 | AAGATCAGCGACGACGATAAG | CTGGTGCTCATACTCCTCTTTC | 105 | 62 | 95 |
| ddit4 | DNA damage-inducible transcript 4 | GGGAATGAGGAGTTTGGTACAT | GAAGGAAGTGGTGGACCTTATT | 88 | 62 | 96 |
| gpx7 | Glutathione peroxidase 7-like | TCTCCTTCCCTCTGTTCAGTAA | GAAATTCCAGTCGGGCTCTT | 104 | 62 | 99 |
| leptin | Leptin | CGAAGAGACTTCCTGCTTCAC | CTGATGATCTGGGTGGACTTTC | 110 | 62 | 104 |
| msh2 | DNA mismatch repair Msh2-like | GCCCGTTCCCAGATATTTGAT | CGACCGCAATGACTACTACAC | 100 | 62 | 98 |
| msh3 | DNA mismatch repair Msh3-like | CGACTTCTTCAGGGACTTTGG | TGGCTCTCTGAGTGTCTGT | 78 | 62 | 104 |
| clql1 | Complement C1q 2-like | GATGTTTGTGGCGACGTATTTG | GTTGCTTTCTCAGCCTCTGTA | 99 | 62 | 104 |
| ccl20 | C-C motif chemokine 20-like | CAGCCGTGTGTTAGGGAATA | CAGTTGTCTCGTGTCTCTCTATC | 123 | 62 | 98 |
| atg12 | Ubiquitin ATG12-like | GCCCTCACCAGATCAAGAAG | AGAGAGTCAGAGTGGAGTTAGAG | 133 | 62 | 102 |
| atg4b | Cysteine protease ATG4B-like | ATCTGGGCGATCTGATGAATG | CGGAGGGCAGAAACAAAGA | 96 | 62 | 102 |
| casp3 | Caspase 3 | GCTCCAATTCTTTCCCGTATTT | CAGATTTCCTCTACGCCTACTC | 123 | 61 | 103 |
| c3 | Complement C3-like | CTGCTTCTGGTGACCTGTTTA | CTTCGTGTCCTCTCCATCTTTC | 99 | 62 | 103 |
| Experimental Samples | Number of Reads | Average Length Number of Reads | Number of Reads after Trimming | Average Length after Trimming | Percentage of Mapped Reads |
|---|---|---|---|---|---|
| Control 1 | 51,220,468 | 101 | 51,103,248 | 95.0 | 85.1 |
| Control 2 | 43,384,328 | 101 | 43,371,547 | 94.9 | 86.0 |
| Control 3 | 50,445,708 | 101 | 50,425,183 | 94.9 | 85.4 |
| Control 4 | 70,505,074 | 101 | 70,462,858 | 94.9 | 84.7 |
| Control 5 | 65,838,802 | 101 | 65,800,592 | 94.9 | 85.7 |
| Control 6 | 57,706,004 | 101 | 57,677,646 | 94.9 | 85.3 |
| Stress 1 | 61,733,308 | 101 | 61,711,175 | 94.9 | 86.8 |
| Stress 2 | 50,630,806 | 101 | 50,612,518 | 94.9 | 84.1 |
| Stress 3 | 58,086,704 | 101 | 58,053,699 | 94.9 | 84.5 |
| Stress 4 | 65,071,756 | 101 | 65,045,365 | 94.9 | 85.5 |
| Stress 5 | 62,370,664 | 101 | 62,343,212 | 94.8 | 84.0 |
| Stress 6 | 59,632,644 | 101 | 59,610,421 | 94.9 | 85.8 |
| Average | 58,052,189 | 101 | 58,018,122 | 94.9 | 85.2 |
| GO Name | GO Category | GO ID | FDR | N° of Transcripts |
|---|---|---|---|---|
| Protein folding | BP | GO:0006457 | 2.96 × 10−11 | 28 |
| Establishment of protein localization | BP | GO:0045184 | 6.62 × 10−3 | 30 |
| Protein transport | BP | GO:0015031 | 1.36 × 10−2 | 29 |
| Nitrogen compound transport | BP | GO:0071705 | 1.65 × 10−2 | 33 |
| Protein localization to endoplasmic reticulum | BP | GO:0070972 | 3.41 × 10−2 | 6 |
| Macromolecule localization | BP | GO:0033036 | 3.41 × 10−2 | 36 |
| Transport | BP | GO:0006810 | 3.41 × 10−2 | 96 |
| Cellular protein localization | BP | GO:0034613 | 3.41 × 10−2 | 25 |
| Organic substance transport | BP | GO:0071702 | 3.41 × 10−2 | 38 |
| Protein localization | BP | GO:0008104 | 3.41 × 10−2 | 31 |
| Response to heat | BP | GO:0009408 | 3.41 × 10−2 | 5 |
| Cellular macromolecule localization | BP | GO:0070727 | 3.41 × 10−2 | 25 |
| Establishment of localization | BP | GO:0051234 | 3.80 × 10−2 | 96 |
| Golgi organization | BP | GO:0007030 | 3.80 × 10−2 | 4 |
| Protein retention in ER lumen | BP | GO:0006621 | 4.09 × 10−2 | 3 |
| Maintenance of protein localization in organelle | BP | GO:0072595 | 4.09 × 10−2 | 3 |
| Maintenance of protein localization in endoplasmic reticulum | BP | GO:0035437 | 4.09 × 10−2 | 3 |
| Intracellular transport | BP | GO:0046907 | 4.09 × 10−2 | 28 |
| Endoplasmic reticulum | CC | GO:0005783 | 3.20 × 10−6 | 33 |
| Cytoplasm | CC | GO:0005737 | 9.42 × 10−6 | 133 |
| Nuclear outer membrane-endoplasmic reticulum membrane network | CC | GO:0042175 | 1.31 × 10−5 | 24 |
| Endoplasmic reticulum sub-compartment | CC | GO:0098827 | 3.68 × 10−5 | 23 |
| Endoplasmic reticulum membrane | CC | GO:0005789 | 3.68 × 10−5 | 23 |
| Endomembrane system | CC | GO:0012505 | 6.46 × 10−5 | 50 |
| Organelle sub-compartment | CC | GO:0031984 | 7.63 × 10−4 | 29 |
| Organelle membrane | CC | GO:0031090 | 2.14 × 10−3 | 44 |
| Sarcomere | CC | GO:0030017 | 3.00 × 10−2 | 7 |
| Myofibril | CC | GO:0030016 | 3.00 × 10−2 | 7 |
| Coated membrane | CC | GO:0048475 | 3.41 × 10−2 | 10 |
| Membrane | CC | GO:0016020 | 3.41 × 10−2 | 225 |
| Membrane coat | CC | GO:0030117 | 3.41 × 10−2 | 10 |
| Coated vesicle membrane | CC | GO:0030662 | 3.41 × 10−2 | 7 |
| Cytoplasmic vesicle membrane | CC | GO:0030659 | 3.41 × 10−2 | 8 |
| Contractile fiber | CC | GO:0043292 | 3.41 × 10−2 | 7 |
| Unfolded protein binding | MF | GO:0051082 | 1.67 × 10−9 | 19 |
| Signal sequence binding | MF | GO:0005048 | 2.70 × 10−3 | 5 |
| ER retention sequence binding | MF | GO:0046923 | 4.09 × 10−2 | 3 |
| Phosphofructokinase activity | MF | GO:0008443 | 4.69 × 10−2 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dettleff, P.; Zuloaga, R.; Fuentes, M.; Gonzalez, P.; Aedo, J.; Estrada, J.M.; Molina, A.; Valdés, J.A. High-Temperature Stress Effect on the Red Cusk-Eel (Geypterus chilensis) Liver: Transcriptional Modulation and Oxidative Stress Damage. Biology 2022, 11, 990. https://doi.org/10.3390/biology11070990
Dettleff P, Zuloaga R, Fuentes M, Gonzalez P, Aedo J, Estrada JM, Molina A, Valdés JA. High-Temperature Stress Effect on the Red Cusk-Eel (Geypterus chilensis) Liver: Transcriptional Modulation and Oxidative Stress Damage. Biology. 2022; 11(7):990. https://doi.org/10.3390/biology11070990
Chicago/Turabian StyleDettleff, Phillip, Rodrigo Zuloaga, Marcia Fuentes, Pamela Gonzalez, Jorge Aedo, Juan Manuel Estrada, Alfredo Molina, and Juan Antonio Valdés. 2022. "High-Temperature Stress Effect on the Red Cusk-Eel (Geypterus chilensis) Liver: Transcriptional Modulation and Oxidative Stress Damage" Biology 11, no. 7: 990. https://doi.org/10.3390/biology11070990
APA StyleDettleff, P., Zuloaga, R., Fuentes, M., Gonzalez, P., Aedo, J., Estrada, J. M., Molina, A., & Valdés, J. A. (2022). High-Temperature Stress Effect on the Red Cusk-Eel (Geypterus chilensis) Liver: Transcriptional Modulation and Oxidative Stress Damage. Biology, 11(7), 990. https://doi.org/10.3390/biology11070990