
Midfacial Morphology and Neandertal–Modern Human Interbreeding



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
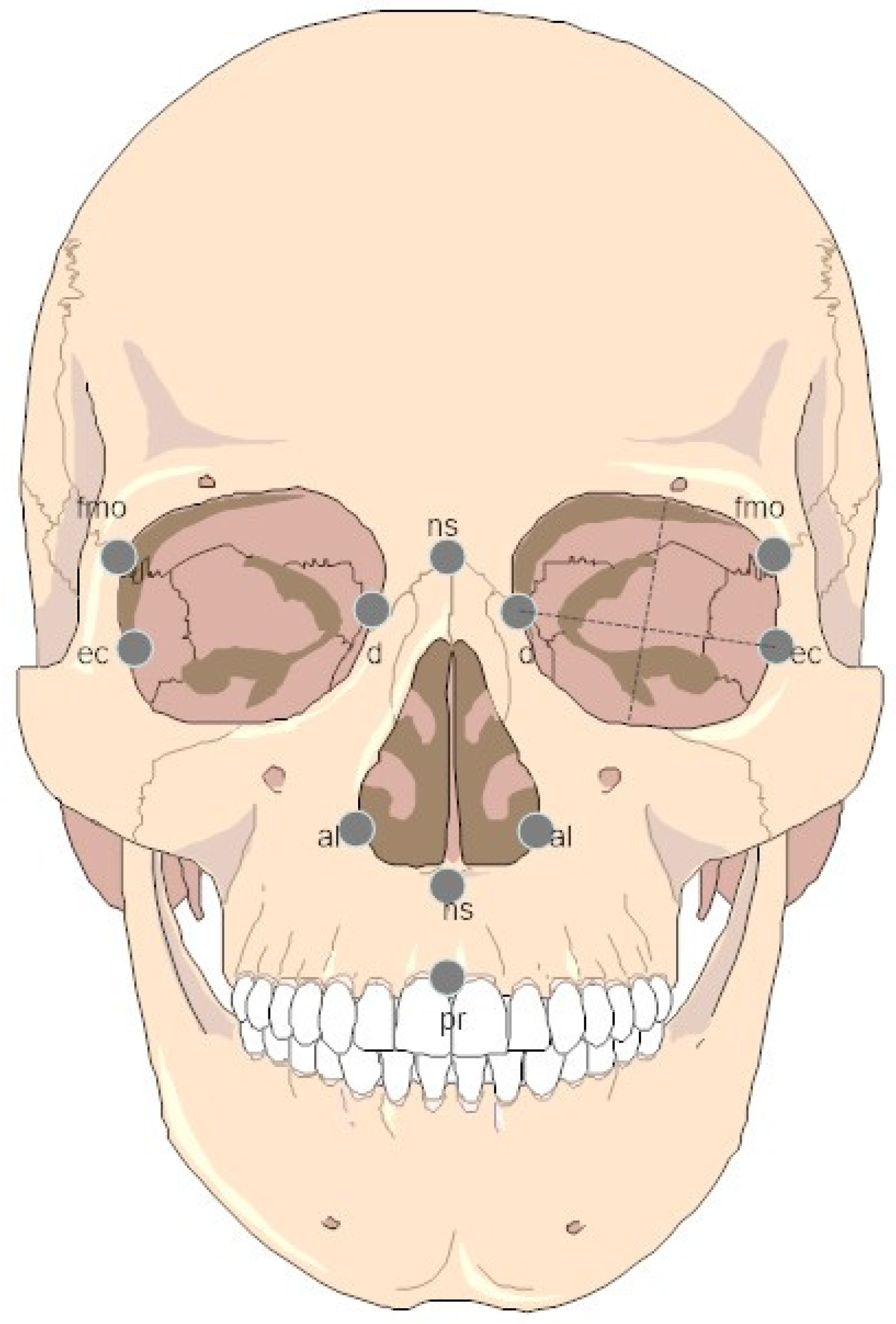
2.1. Samples and Measurements
2.2. Multivariate Statistics
2.3. Spatial Analyses
3. Results
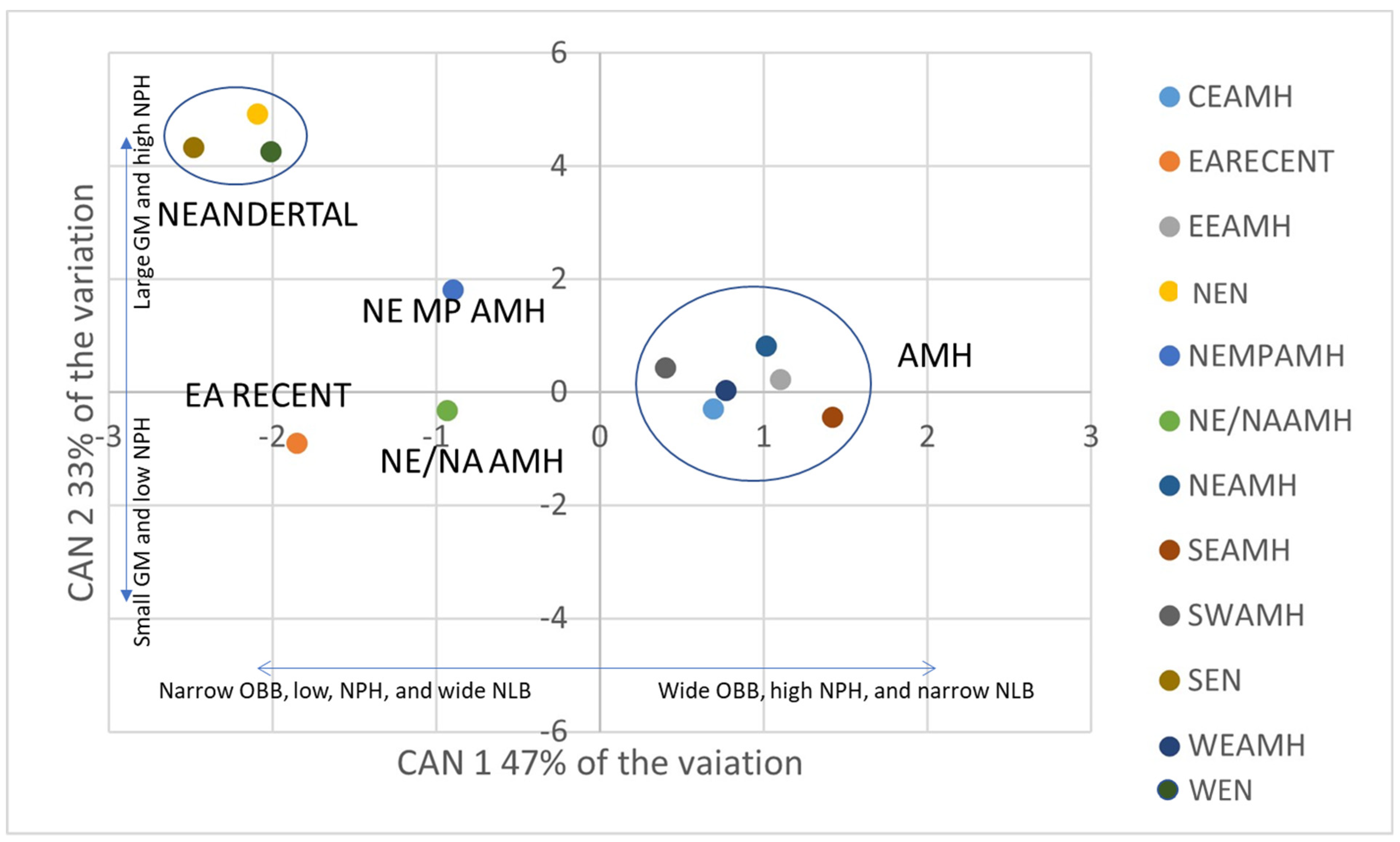
3.1. Multivariate Statistics
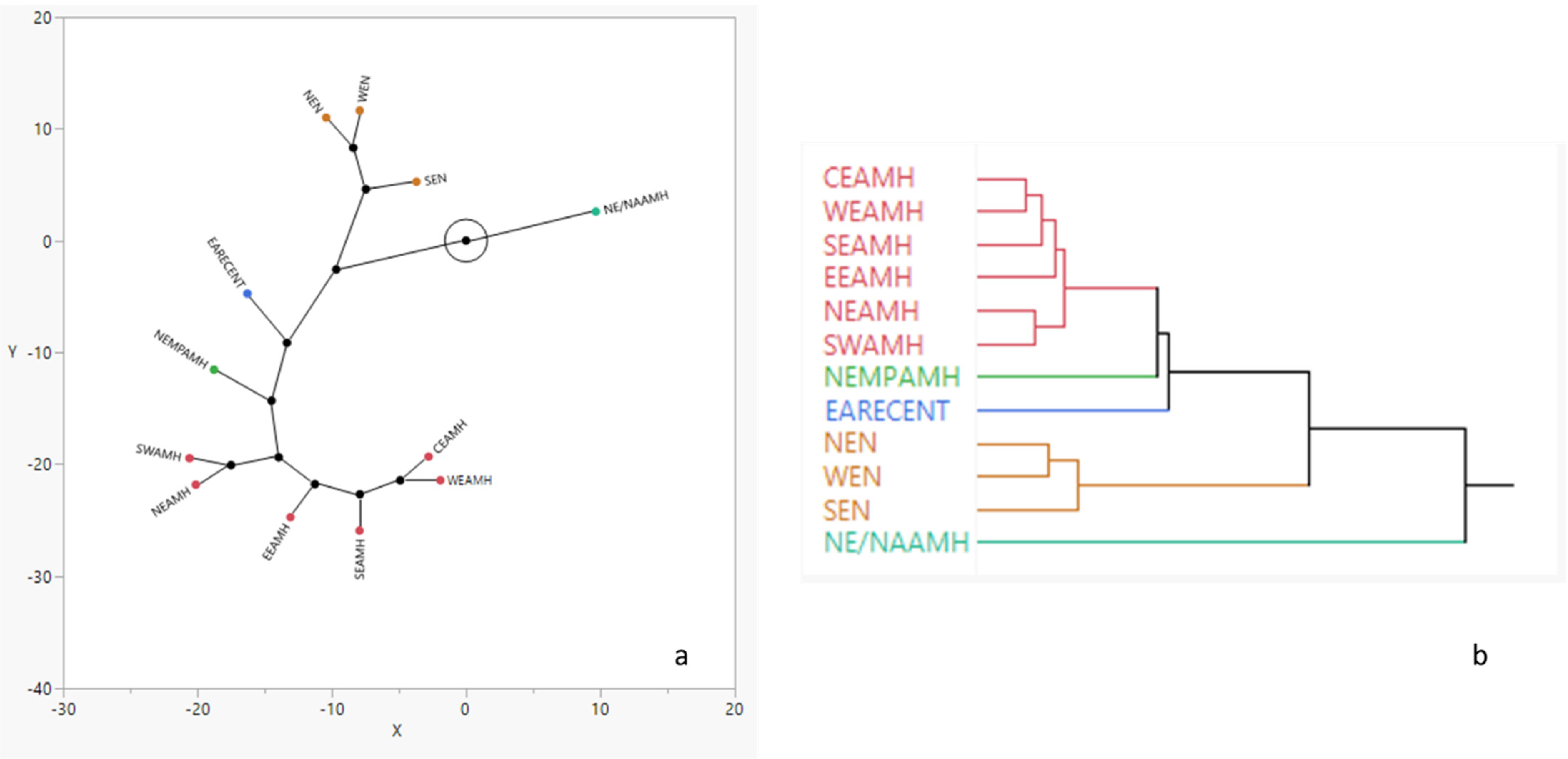
3.2. Spatial Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Krause, J.; Briggs, A.W.; Kircher, M.; Maricic, T.; Zwyns, N.; Derevianko, A.; Pääbo, S. A Complete mtDNA Genome of an Early Modern Human from Kostenki, Russia. Curr. Biol. 2010, 20, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gokcumen, O. Archaic hominin introgression into modern human genomes. Am. J. Phys. Anthr. 2019, 171, 60–73. [Google Scholar] [CrossRef]
- Brown, P.; Sutikna, T.; Morwood, M.J.; Soejono, R.P.; Jatmiko; Saptomo, E.W.; Due, R.A. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 2004, 431, 1055–1061. [Google Scholar] [CrossRef]
- Berger, L.R.; Hawks, J.; de Ruiter, D.J.; Churchill, S.E.; Schmid, P.; Delezene, L.K.; Kivell, T.L.; Garvin, H.M.; Williams, S.A.; DeSilva, J.M.; et al. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife 2015, 4, e09560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Détroit, F.; Mijares, A.S.; Corny, J.; Daver, G.; Zanolli, C.; Dizon, E.; Robles, E.; Grün, R.; Piper, P.J. A new species of Homo from the Late Pleistocene of the Philippines. Nature 2019, 568, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Hammer, M.F.; Woerner, A.E.; Mendez, F.L.; Watkins, J.C.; Wall, J.D. Genetic evidence for archaic admixture in Africa. Proc. Natl. Acad. Sci. USA 2011, 108, 15123–15128. [Google Scholar] [CrossRef] [Green Version]
- Berger, L.R.; Hawks, J.; Dirks, P.H.; Elliott, M.; Roberts, E.M. Homo naledi and Pleistocene hominin evolution in subequatorial Africa. eLife 2017, 6, e24234. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, A.P.; Gravel, S. Models of archaic admixture and recent history from two-locus statistics. PLoS Genet. 2019, 15, e1008204. [Google Scholar] [CrossRef] [Green Version]
- Hollfelder, N.; Breton, G.; Sjödin, P.; Jakobsson, M. The deep population history in Africa. Hum. Mol. Genet. 2021, 30, R2–R10. [Google Scholar] [CrossRef]
- Hublin, J.-J.; Ben-Ncer, A.; Bailey, S.E.; Freidline, S.E.; Neubauer, S.; Skinner, M.M.; Bergmann, I.; LE Cabec, A.; Benazzi, S.; Harvati-Papatheodorou, K.; et al. New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature 2017, 546, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Harvati, K.; Röding, C.; Bosman, A.; Karakostis, F.A.; Grün, R.; Stringer, C.; Karkanas, P.; Thompson, N.C.; Koutoulidis, V.; Moulopoulos, L.; et al. Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia. Nature 2019, 571, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Hershkovitz, I.; Weber, G.W.; Quam, R.; Duval, M.; Grün, R.; Kinsley, L.; Ayalon, A.; Bar-Matthews, M.; Valladas, H.; Mercier, N.; et al. The earliest modern humans outside Africa. Science 2018, 359, 456–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Welker, F.; Shen, C.-C.; Bailey, S.; Bergmann, I.; Davis, S.; Xia, H.; Wang, H.; Fischer, R.; Freidline, S.E.; et al. A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau. Nature 2019, 569, 409–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, X.; Ji, Q.; Wu, W.; Shao, Q.; Ji, Y.; Zhang, C.; Liang, L.; Ge, J.; Guo, Z.; Li, J.; et al. Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage. Innovation 2021, 2, 100130. [Google Scholar] [CrossRef]
- Reich, D.; Patterson, N.; Kircher, M.; Delfin, F.; Nandineni, M.R.; Pugach, I.; Ko, A.M.-S.; Ko, Y.-C.; Jinam, T.A.; Phipps, M.E.; et al. Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. Am. J. Hum. Genet. 2011, 89, 516–528. [Google Scholar] [CrossRef] [Green Version]
- Lachance, J.; Vernot, B.; Elbers, C.C.; Ferwerda, B.; Froment, A.; Bodo, J.-M.; Lema, G.; Fu, W.; Nyambo, T.B.; Rebbeck, T.R.; et al. Evolutionary History and Adaptation from High-Coverage Whole-Genome Sequences of Diverse African Hunter-Gatherers. Cell 2012, 150, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Slon, V.; Mafessoni, F.; Vernot, B.; De Filippo, C.; Grote, S.; Viola, B.; Hajdinjak, M.; Peyrégne, S.; Nagel, S.; Brown, S.; et al. The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature 2018, 561, 113–116. [Google Scholar] [CrossRef]
- Bailey, S.E.; Hublin, J.-J.; Antón, S.C. Rare dental trait provides morphological evidence of archaic introgression in Asian fossil record. Proc. Natl. Acad. Sci. USA 2019, 116, 14806–14807. [Google Scholar] [CrossRef] [Green Version]
- Bergström, A.; Stringer, C.; Hajdinjak, M.; Scerri, E.M.L.; Skoglund, P. Origins of modern human ancestry. Nature 2021, 590, 229–237. [Google Scholar] [CrossRef]
- Rogers, A.R.; Bohlender, R.J.; Huff, C.D. Early history of Neanderthals and Denisovans. Proc. Natl. Acad. Sci. USA 2017, 114, 9859–9863. [Google Scholar] [CrossRef] [Green Version]
- Sankararaman, S.; Patterson, N.; Li, H.; Pääbo, S.; Reich, D. The Date of Interbreeding between Neandertals and Modern Humans. PLoS Genet. 2012, 8, e1002947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanea, F.A.; Schraiber, J.G. Multiple episodes of interbreeding between Neanderthal and modern humans. Nat. Ecol. Evol. 2018, 3, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Green, R.E.; Krause, J.; Briggs, A.W.; Maricic, T.; Stenzel, U.; Kircher, M.; Patterson, N.; Li, H.; Zhai, W.; Fritz, M.H.-Y.; et al. A Draft Sequence of the Neandertal Genome. Science 2010, 328, 710–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhlwilm, M.; Gronau, I.; Hubisz, M.; De Filippo, C.; Prado-Martinez, J.; Kircher, M.; Fu, Q.; Burbano, H.A.; Lalueza-Fox, C.; De La Rasilla, M.; et al. Ancient gene flow from early modern humans into Eastern Neanderthals. Nature 2016, 530, 429–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.; Kircher, M.; Gansauge, M.-T.; Li, H.; Racimo, F.; Mallick, S.; Schraiber, J.G.; Jay, F.; Prüfer, K.; De Filippo, C.; et al. A High-Coverage Genome Sequence from an Archaic Denisovan Individual. Science 2012, 338, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Wall, J.D.; Yang, M.A.; Jay, F.; Kim, S.K.; Durand, E.Y.; Stevison, L.; Gignoux, C.; Woerner, A.; Hammer, M.F.; Slatkin, M. Higher Levels of Neanderthal Ancestry in East Asians than in Europeans. Genetics 2013, 194, 199–209. [Google Scholar] [CrossRef]
- Currat, M.; Excoffier, L. Modern Humans Did Not Admix with Neanderthals during Their Range Expansion into Europe. PLoS Biol. 2004, 2, e421. [Google Scholar] [CrossRef]
- Hajdinjak, M.; Fu, Q.; Hübner, A.; Petr, M.; Mafessoni, F.; Grote, S.; Skoglund, P.; Narasimham, V.; Rougier, H.; Crevecoeur, I.; et al. Reconstructing the genetic history of late Neanderthals. Nature 2018, 555, 652–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hublin, J.-J. The modern human colonization of western Eurasia: When and where? Quat. Sci. Rev. 2015, 118, 194–210. [Google Scholar] [CrossRef] [Green Version]
- Quilodrán, C.S.; Tsoupas, A.; Currat, M. The Spatial Signature of Introgression After a Biological Invasion with Hybridization. Front. Ecol. Evol. 2020, 8, 569620. [Google Scholar] [CrossRef]
- Fu, Q.; Li, H.; Moorjani, P.; Jay, F.; Slepchenko, S.M.; Bondarev, A.A.; Johnson, P.L.F.; Aximu-Petri, A.; Prüfer, K.; De Filippo, C.; et al. Genome sequence of a 45,000-year-old modern human from western Siberia. Nature 2014, 514, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Vernot, B.; Akey, J.M. Complex History of Admixture between Modern Humans and Neandertals. Am. J. Hum. Genet. 2015, 96, 448–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguin-Orlando, A.; Korneliussen, T.S.; Sikora, M.; Malaspinas, A.-S.; Manica, A.; Moltke, I.; Albrechtsen, A.; Ko, A.; Margaryan, A.; Moiseyev, V.; et al. Genomic structure in Europeans dating back at least 36,200 years. Science 2014, 346, 1113–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Hajdinjak, M.; Moldovan, O.T.; Constantin, S.; Mallick, S.; Skoglund, P.; Patterson, N.; Rohland, N.; Lazaridis, I.; Nickel, B.; et al. An early modern human from Romania with a recent Neanderthal ancestor. Nature 2015, 524, 216–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soficaru, A.; Doboş, A.; Trinkaus, E. Early modern humans from the Peştera Muierii, Baia de Fier, Romania. Proc. Natl. Acad. Sci. USA 2006, 103, 17196–17201. [Google Scholar] [CrossRef] [Green Version]
- Rougier, H.; Milota, S.; Rodrigo, R.; Gherase, M.; Sarcinǎ, L.; Moldovan, O.; Zilhão, J.; Constantin, S.; Franciscus, R.G.; Zollikofer, C.P.E.; et al. Peştera cu Oase 2 and the cranial morphology of early modern Europeans. Proc. Natl. Acad. Sci. USA 2007, 104, 1165–1170. [Google Scholar] [CrossRef] [Green Version]
- Trinkaus, E. European early modern humans and the fate of the Neandertals. Proc. Natl. Acad. Sci. USA 2007, 104, 7367–7372. [Google Scholar] [CrossRef] [Green Version]
- Harvaati, K.; Roksandic, M. The Human Fossil Record from Romania: Early Upper Paleolithic European Mandibles and Neanderthal Admixture. In Paleoanthropology of the Balkans and Anatolia; Springer: Berlin/Heidelberg, Germany, 2016; pp. 51–68. [Google Scholar] [CrossRef]
- Hajdinjak, M.; Mafessoni, F.; Skov, L.; Vernot, B.; Hübner, A.; Fu, Q.; Essel, E.; Nagel, S.; Nickel, B.; Richter, J.; et al. Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry. Nature 2021, 592, 253–257. [Google Scholar] [CrossRef]
- Prüfer, K.; Posth, C.; Yu, H.; Stoessel, A.; Spyrou, M.A.; Deviese, T.; Mattonai, M.; Ribechini, E.; Higham, T.; Velemínský, P.; et al. A genome sequence from a modern human skull over 45,000 years old from Zlatý kůň in Czechia. Nat. Ecol. Evol. 2021, 5, 820–825. [Google Scholar] [CrossRef]
- Bräuer, G.; Broeg, H.; Stringer, C.B. Earliest Upper Paleolithic crania from Mladeč, Czech Republic, and the question of Neanderthal-modern continuity: Metrical evidence from the fronto-facial region. In Neanderthals Revisited: New Approaches and Perspectives. Vertebrate Paleobiology and Paleoanthropology; Hublin, J.J., Harvati, K., Harrison, T., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 269–279. [Google Scholar] [CrossRef]
- Frayer, D.W. The Persistence of Neanderthal Features in post-Neanderthal Europeans. In Continuity or Replacement: Controversies in Homo Sapiens Evolution; Bräuer, G., Smith, F., Eds.; A.A. Balkema: Rotterdam, The Netherlnads, 1992; pp. 179–188. [Google Scholar]
- Bräuer, G.; Broeg, H. On the Degree of Neandertal-modern Continuity in the Earliest Upper Palaeolithic Crania from the Czech Republic: Evidence from Non-metrical Features. In The Origins and Past of Modern Humans—Towards Reconciliation; Omoto, K., Tobias, P.V., Eds.; World Scientific: Singapore, 1998; pp. 106–125. [Google Scholar]
- Churchill, S.E.; Smith, F.H. Makers of The Early Aurignacian Of Europe. Am. J. Phys. Anthropol. 2000, 113, 61–115. [Google Scholar] [CrossRef]
- Condemi, S.; Mounier, A.; Giunti, P.; Lari, M.; Caramelli, D.; Longo, L. Possible Interbreeding in Late Italian Neanderthals? New Data from the Mezzena Jaw (Monti Lessini, Verona, Italy). PLoS ONE 2013, 8, e59781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, C.; Maurício, J.; Pettitt, P.B.; Souto, P.; Trinkaus, E.; van der Plicht, H.; Zilhão, J. The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia. Proc. Natl. Acad. Sci. USA 1999, 96, 7604–7609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozzi, R.; Fernando, V.; d’Errico, F.; Vanhaeren, M.; Grootes, P.M.; Kerautret, B.; Dujardin, V. Cutmarked Human Remains Bearing Neandertal Features and Modern Human Remains Associated with the Aurignacian at Les Rois. J. Anthropol. Sci. 2009, 87, 153–185. [Google Scholar]
- Compton, T.; Skinner, M.M.; Humphrey, L.; Pope, M.; Bates, M.; Davies, T.W.; Parfitt, S.A.; Plummer, W.P.; Scott, B.; Shaw, A.; et al. The morphology of the Late Pleistocene hominin remains from the site of La Cotte de St Brelade, Jersey (Channel Islands). J. Hum. Evol. 2021, 152, 102939. [Google Scholar] [CrossRef]
- Walker, M.J.; Gibert, J.; López, M.V.; Lombardi, A.V.; Pérez-Pérez, A.; Zapata, J.; Ortega, J.; Higham, T.; Pike, A.; Schwenninger, J.-L.; et al. Late Neandertals in Southeastern Iberia: Sima de las Palomas del Cabezo Gordo, Murcia, Spain. Proc. Natl. Acad. Sci. USA 2008, 105, 20631–20636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolpoff, M.H. Paleoanthropology, 2nd ed.; McGraw-Hill: Boston, MA, USA, 1999. [Google Scholar]
- Trinkaus, E.; Churchill, S.E.; Ruff, C.B.; Vandermeersch, B. Long Bone Shaft Robusticity and Body Proportions of the Saint-Césaire 1 Châtelperronian Neanderthal. J. Archaeol. Sci. 1999, 26, 753–773. [Google Scholar] [CrossRef]
- Di Vincenzo, F.; Churchill, S.E.; Manzi, G. The Vindija Neanderthal scapular glenoid fossa: Comparative shape analysis suggests evo-devo changes among Neanderthals. J. Hum. Evol. 2012, 62, 274–285. [Google Scholar] [CrossRef]
- Wolpoff, M.H.; Smith, F.H.; Malez, M.; Radovčić, J.; Rukavina, D. Upper pleistocene human remains from Vindija cave, Croatia, Yugoslavia. Am. J. Phys. Anthr. 1981, 54, 499–545. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.H.; Trinkaus, E. Les origines de l’homme moderne en Europe centrale: Un cas de continuité. In Aux Origines d’Homo sapiens; Hublin, J.-J., Tillier, A.-M., Eds.; Presses Universitaires de France: Paris, France, 1991; pp. 251–290. [Google Scholar]
- Ahern, J.C.; Karavanić, I.; Paunović, M.; Janković, I.; Smith, F.H. New discoveries and interpretations of hominid fossils and artifacts from Vindija Cave, Croatia. J. Hum. Evol. 2003, 46, 27–67. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, R.R. Phenotypic traits of primate hybrids: Recognizing admixture in the fossil record. Evol. Anthr. Issues News Rev. 2010, 19, 258–270. [Google Scholar] [CrossRef]
- Rink, W.J.; Schwarcz, H.P.; Smith, F.H.; Radovĉiĉ, J. ESR ages for Krapina hominids. Nature 1995, 378, 24. [Google Scholar] [CrossRef]
- Rosas, A.; Bastir, M. An assessment of the late Middle Pleistocene occipital from Apidima 1 skull (Greece). L'Anthropologie 2020, 124, 102745. [Google Scholar] [CrossRef]
- Smith, H.F. Which cranial regions reflect molecular distances reliably in humans? Evidence from three-dimensional morphology. Am. J. Hum. Biol. 2008, 21, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.W.; Hershkovitz, I.; Gunz, P.; Neubauer, S.; Ayalon, A.; Latimer, B.; Bar-Matthews, M.; Yasur, G.; Barzilai, O.; May, H. Before the massive modern human dispersal into Eurasia: A 55,000-year-old partial cranium from Manot Cave, Israel. Quat. Int. 2019, 551, 29–39. [Google Scholar] [CrossRef]
- Trinkaus, E. Modern Human versus Neandertal Evolutionary Distinctiveness. Curr. Anthr. 2006, 47, 597–620. [Google Scholar] [CrossRef]
- Freidline, S.E.; Gunz, P.; Harvati, K.; Hublin, J.-J. Middle Pleistocene human facial morphology in an evolutionary and developmental context. J. Hum. Evol. 2012, 63, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Arensburg, B. New Upper Palaeolithic Human Remains from Israel. In Eretz-Israel: Archaeological, Historical and Geographical Studies; Israel Exploration Society (IES): Jerusalem, Israel, 1977; pp. 208–215. Available online: http://www.jstor.org/stable/23618749 (accessed on 5 July 2022).
- Brewster, C.; Meiklejohn, C.; Von Cramon-Taubadel, N.; Pinhasi, R. Craniometric analysis of European Upper Palaeolithic and Mesolithic samples supports discontinuity at the Last Glacial Maximum. Nat. Commun. 2014, 5, 4094. [Google Scholar] [CrossRef] [Green Version]
- Lahr, M.M. The question of robusticity and the relationship between cranial size and shape inHomo sapiens. J. Hum. Evol. 1996, 31, 157–191. [Google Scholar] [CrossRef]
- Trinkaus, E.; Milota, S.; Rodrigo, R.; Mircea, G.; Moldovan, O. Early modern human cranial remains from the Peştera cu Oase, Romania. J. Hum. Evol. 2003, 45, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandermeersch, B.; Arensburg, B.; Ofer, B.-Y.; Belfer-Cohen, A. Upper Paleolithic Human Remains from Qafzeh Cave, Israel. Mitekufat Haeven. J. Isr. Prehist. Soc. 2013, 43, 7–21. Available online: http://www.jstor.org/stable/23784046. (accessed on 5 July 2022).
- Weaver, T.D.; Stringer, C.B. Unconstrained cranial evolution in Neandertals and modern humans compared to common chimpanzees. Proc. R. Soc. B Boil. Sci. 2015, 282, 20151519. [Google Scholar] [CrossRef] [Green Version]
- Fabre, V.; Condemi, S.; Degioanni, A. Genetic Evidence of Geographical Groups among Neanderthals. PLoS ONE 2009, 4, e5151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, A.; Williams, S. Ancestry Studies in Forensic Anthropology: Back on the Frontier of Racism. Biology 2021, 10, 602. [Google Scholar] [CrossRef] [PubMed]
- Howells, W.W. Cranial Variation in Man; Harvard University Press: Cambridge, MA, USA, 1973. [Google Scholar]
- Stone, J.H.; Chew, K.; Ross, A.H.; Verano, J.W. Craniofacial plasticity in ancient Peru. Anthr. Anz. 2015, 72, 169–183. [Google Scholar] [CrossRef]
- Hubbe, M.; Hanihara, T.; Harvati, K. Climate Signatures in the Morphological Differentiation of Worldwide Modern Human Populations. Anat. Rec. 2009, 292, 1720–1733. [Google Scholar] [CrossRef]
- Ross, A.H.; Ubelaker, D.H. Effect of Intentional Cranial Modification on Craniofacial Landmarks. J. Craniofacial Surg. 2009, 20, 2185–2187. [Google Scholar] [CrossRef]
- Sardi, M.L.; Rozzi, F.R.; Dahinten, S.L.; Pucciarelli, H.M. Amerindians: Testing the hypothesis about their homogeneity. Comptes Rendus Palevol. 2004, 3, 403–409. [Google Scholar] [CrossRef]
- Metzger, M.J.; Bunce, R.G.H.; Jongman, R.H.G.; Sayre, R.; Trabucco, A.; Zomer, R. A high-resolution bioclimate map of the world: A unifying framework for global biodiversity research and monitoring. Glob. Ecol. Biogeogr. 2012, 22, 630–638. [Google Scholar] [CrossRef] [Green Version]
- Frumkin, A.; Bar-Yosef, O.; Schwarcz, H.P. Possible paleohydrologic and paleoclimatic effects on hominin migration and occupation of the Levantine Middle Paleolithic. J. Hum. Evol. 2011, 60, 437–451. [Google Scholar] [CrossRef]
- Nicholson, C.M. Eemian paleoclimate zones and Neanderthal landscape-use: A GIS model of settlement patterning during the last interglacial. Quat. Int. 2017, 438, 144–157. [Google Scholar] [CrossRef]
- Pederzani, S.; Aldeias, V.; Dibble, H.L.; Goldberg, P.; Hublin, J.-J.; Madelaine, S.; McPherron, S.P.; Sandgathe, D.; Steele, T.E.; Turq, A.; et al. Reconstructing Late Pleistocene paleoclimate at the scale of human behavior: An example from the Neandertal occupation of La Ferrassie (France). Sci. Rep. 2021, 11, 1419. [Google Scholar] [CrossRef] [PubMed]
- Darroch, J.N.; Mosimann, J.E. Canonical and principal components of shape. Biometrika 1985, 72, 241–252. [Google Scholar] [CrossRef]
- Ross, A.H. Regional isolation in the Balkan region: An analysis of craniofacial variation. Am. J. Phys. Anthr. 2003, 124, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Pietrusewsky, M. Traditional Morphometrics and Biological Distance: Methods and an Example. In Biological Anthropology of the Human Skeleton, 3rd ed.; Katzenberg, M.A., Grauer, A.L., Eds.; Wiley Blackwell: Oxford, UK, 2019; pp. 547–591. [Google Scholar]
- SAS Institute Inc. SAS 9.4; SAS Institute Inc.: Cary, NC, USA, 1989. [Google Scholar]
- SAS Institute Inc. JMP, Version 16 ed.; SAS Institute Inc.: Cary, NC, USA, 2016. [Google Scholar]
- Dutilleul, P.; Clifford, P.; Richardson, S.; Hemon, D. Modifying the t Test for Assessing the Correlation Between Two Spatial Processes. Biometrics 1993, 49, 305. [Google Scholar] [CrossRef]
- Rosenberg, M.S.; Anderson, C.D. PASSaGE: Pattern Analysis, Spatial Statistics and Geographic Exegesis. Version 2. Methods Ecol. Evol. 2010, 2, 229–232. [Google Scholar] [CrossRef]
- Franciscus, R.G. Later Pleistocene Nasofacial Variation in Western Eurasia and Africa and Modern Human Origins. Ph.D. Thesis, University of New Mexico, Albuquerque, NM, USA, 1995. [Google Scholar]
- Franciscus, R.G. Internal nasal floor configuration in Homo with special reference to the evolution of Neandertal facial form. J. Hum. Evol. 2003, 44, 701–729. [Google Scholar] [CrossRef]
- Hublin, J.-J. Climatic changes, paleogeography, and the evolution of the Neandertals. In Neandertals and Modern Humans in Western Asia; Akazawa, T., Aoki, K., Bar-Yosef, O., Eds.; Plenum: New York, NY, USA, 1998; pp. 295–310. [Google Scholar]
- Trinkaus, E. Neandertal faces were not long; modern human faces are short. Proc. Natl. Acad. Sci. USA 2003, 100, 8142–8145. [Google Scholar] [CrossRef] [Green Version]
- Trinkaus, E. The Neandertal face: Evolutionary and functional perspectives on a recent hominid face. J. Hum. Evol. 1987, 16, 429–443. [Google Scholar] [CrossRef]
- Ross, A.H.; Ubelaker, D.H. Complex Nature of Hominin Dispersals: Ecogeographical and Climatic Evidence for Pre-Contact Craniofacial Variation. Sci. Rep. 2019, 9, 11743. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | Abbreviation | Description |
|---|---|---|
| Upper Facial Height (n-pr) | NPH | [71] |
| Bifrontal Breadth (fmo-fmo) | FMB | [71] |
| Nasal Height (n-ns) | NLH | [71] |
| Nasal Breadth (al-al) | NLB | [71] |
| Orbital Height | OBH | [71] |
| Orbital Breadth (d-ec) | OBB | [71] |
| CE AMH | EA RECENT | EE AMH | NE MP AMH | NE NEAN | NE/NA AMH | NE AMH | SE AMH | SW AMH | SE NEAN | WE AMH | WE NEAN | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NPH | 67 (5.21) | 63 (4.8) | 70 (5.28) | 75 (2.7) | 88 (6.06) | 65 (4.16) | 72 (4.45) | 67 (4.15) | 70 (4.6) | 88 (14.14) | 68 (5.2) | 85 (4.9) |
| FMB | 96 (4.87) | 97 4.04) | 96 (5.32) | 103 (5.54) | 114 (4.18) | 103.5 (3.5) | 101 (4.62) | 94 (5.05) | 99 (5.01) | 111 (10.52) | 98 (4.56) | 113 (1.2) |
| NLH | 50 (4.7) | 48 (3.55) | 52 (3.5) | 54 (1.3) | 63 (3.9) | 46 (1.76) | 52 (3.19) | 49 (3.16) | 52 (3.71) | 59 (4.28) | 50 (4.09) | 60 (0.6) |
| NLB | 25 (2.21) | 28 (1.87) | 25 (2.58) | 30 (1.41) | 34 (2.88) | 26 (1.54) | 24 (2.06) | 24 (2.05) | 25 (1.9) | 34 (4.3) | 25 (2.17) | 32 (3.8) |
| OBB | 42 (3.04) | 39 (1.76) | 43 (2.54) | 44 (1.79) | 46 (1.24) | 42 (3.8) | 43 (1.73 | 42 (.67) | 42 (2.56) | 44 (1.22) | 41 (2.9) | 44 (2.1) |
| OBH | 30 (2.41) | 33 (1.9) | 32 (2.25) | 33 (3.39) | 36 (0.89) | 28 (1.53) | 32 (2.46) | 30 (2.5) | 33 (3.31) | 37 (1.52) | 30 (2.0) | 37 (1.2) |
| CEA AMH | EA RECENT | EE AMH | NE MP AMH | NE NEAN | NE/NA AMH | NE AMH | SE AMH | SW AMH | SE NEAN | WE AMH | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| EA RECENT | 7.15 | 0 | |||||||||
| EE AMH | 1.22 | 10.63 | 0 | ||||||||
| NE MP AMH | 7.78 | 9.62 | 7.96 | 0 | |||||||
| NE NEAN | 35.19 | 34.54 | 32.99 | 11.88 | 0 | ||||||
| NE/NA AMH | 51.76 | 52.84 | 57.27 | 59.71 | 81.24 | 0 | |||||
| NE AMH | 2.59 | 11.59 | 1.99 | 7.64 | 28.19 | 54.78 | 0 | ||||
| SE AMH | 0.89 | 11.13 | 1.51 | 11.56 | 42.00 | 55.13 | 2.57 | 0 | |||
| SWAMH | 1.70 | 7.15 | 1.22 | 6.08 | 27.66 | 54.25 | 0.82 * | 2.60 | 0 | ||
| SE NEAN | 32.87 | 29.27 | 32.20 | 11.27 | 2.99 * | 69.72 | 26.82 | 38.68 | 25.92 | 0 | |
| WE AMH | 0.56 * | 8.04 | 1.81 | 7.81 | 32.83 | 52.40 | 1.15 | 1.14 | 1.18 | 30.58 | 0 |
| WE NEAN | 28.71 | 26.83 | 27.15 | 9.25 | 1.28 * | 74.71 | 21.29 | 34.40 | 20.87 | 2.03 * | 25.76 |
| No. | Eigenvalue | Cumulative % | Proportion | Likelihood Ratio | Approximate F | Num DF | Den DF | Pr > F |
|---|---|---|---|---|---|---|---|---|
| 1 | 1.80 | 0.47 | 0.47 | 0.08 | 12.17 | 77 | 1793 | <0.0001 |
| 2 | 1.28 | 0.80 | 0.33 | 0.23 | 8.62 | 60 | 1572 | <0.0001 |
| 3 | 0.51 | 0.96 | 0.03 | 0.78 | 4.79 | 45 | 1111 | <0.0001 |
| 4 | 0.12 | 0.98 | 0.03 | 0.70 | 2.48 | 32 | 1542 | <0.0001 |
| 5 | 0.09 | 0.99 | 0.02 | 0.87 | 2.04 | 21 | 868 | 0.004 |
| Variable | CAN 1 | CAN 2 | CAN 3 | CAN 4 | CAN 5 |
|---|---|---|---|---|---|
| GM | −0.27 | 0.93 | −0.08 | 0.05 | −0.07 |
| NPHs | 0.50 | 0.60 | 0.15 | 0.04 | 0.06 |
| FMBs | −0.06 | −0.25 | 0.31 | −0.56 | 0.39 |
| NLHs | 0.38 | 0.25 | −0.04 | 0.12 | −0.13 |
| NLBs | −0.81 | −0.11 | 0.12 | 0.29 | 0.11 |
| OBBs | 0.71 | −0.12 | 0.07 | 0.07 | −0.07 |
| OBHs | −0.37 | −0.45 | −0.28 | −0.31 | −0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Churchill, S.E.; Keys, K.; Ross, A.H. Midfacial Morphology and Neandertal–Modern Human Interbreeding. Biology 2022, 11, 1163. https://doi.org/10.3390/biology11081163
Churchill SE, Keys K, Ross AH. Midfacial Morphology and Neandertal–Modern Human Interbreeding. Biology. 2022; 11(8):1163. https://doi.org/10.3390/biology11081163
Chicago/Turabian StyleChurchill, Steven E., Kamryn Keys, and Ann H. Ross. 2022. "Midfacial Morphology and Neandertal–Modern Human Interbreeding" Biology 11, no. 8: 1163. https://doi.org/10.3390/biology11081163
APA StyleChurchill, S. E., Keys, K., & Ross, A. H. (2022). Midfacial Morphology and Neandertal–Modern Human Interbreeding. Biology, 11(8), 1163. https://doi.org/10.3390/biology11081163