
From Molecular to Functional Effects of Different Environmental Lead Exposure Paradigms

 , ,
, , 
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
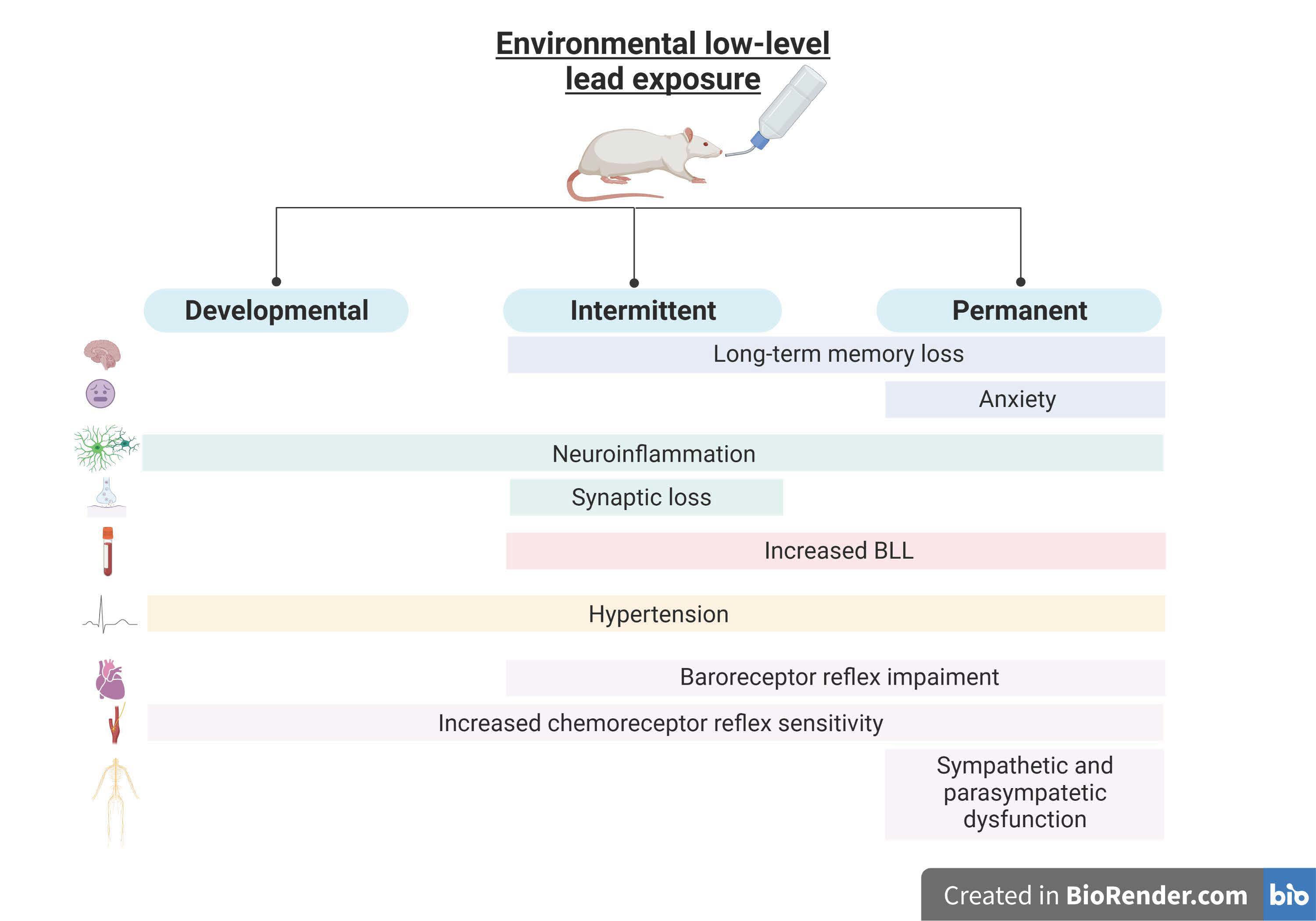
2.1. Experimental Groups
2.2. Behavioral Evaluation
2.2.1. Elevated plus Maze
2.2.2. Open-Field Exploration Test
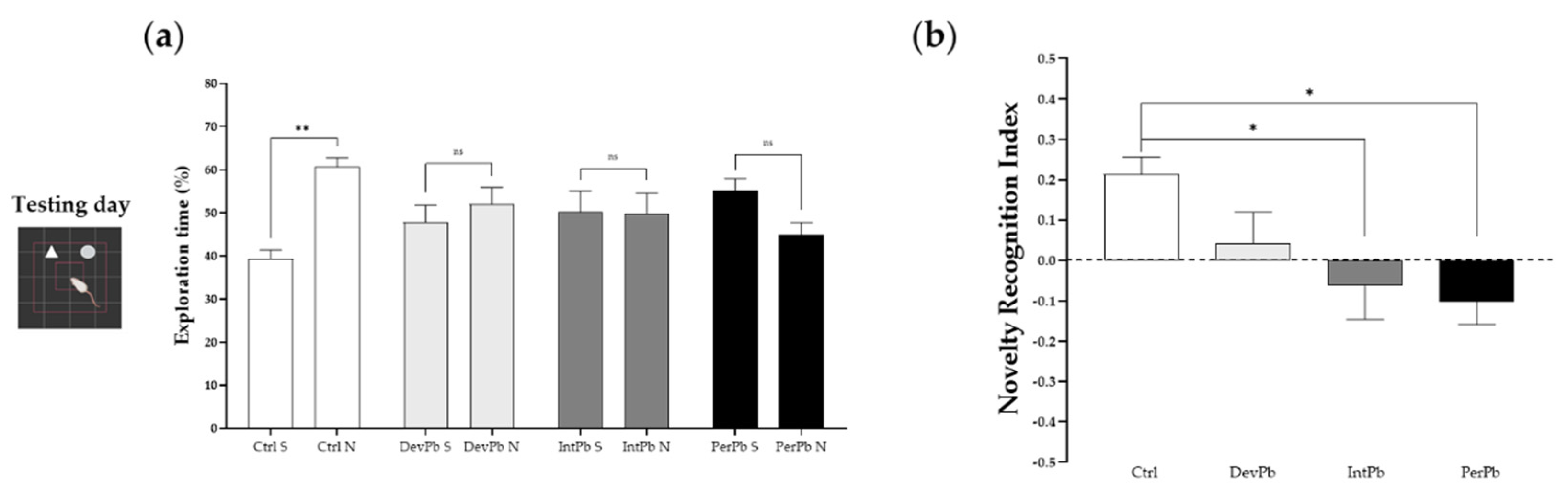
2.2.3. Novel Object Recognition Test
2.3. Metabolic Evaluation
2.4. Functional Evaluation
2.4.1. Acute Physiological Studies
2.4.2. Data Acquisition and Analysis
2.4.3. Baro- and Chemoreceptor Reflex Analysis
2.5. Immunohistochemistry (IHC)
2.6. Statistical Analysis
3. Results
3.1. Lead Exposure from the Fetal Period until Adulthood Caused Anxiety and Cognitive Impairment
3.1.1. Permanent Exposure to Lead Caused Strong Anxiety without Locomotor Changes
3.1.2. Permanent and Intermittent Lead Exposures Generated Strong Long-Term Memory Impairment
3.2. Lead Exposure from the Fetal Period until Adulthood Provoked Neuroinflammation and Synaptic Alterations without Neurodegeneration in the Dentate Gyrus Region
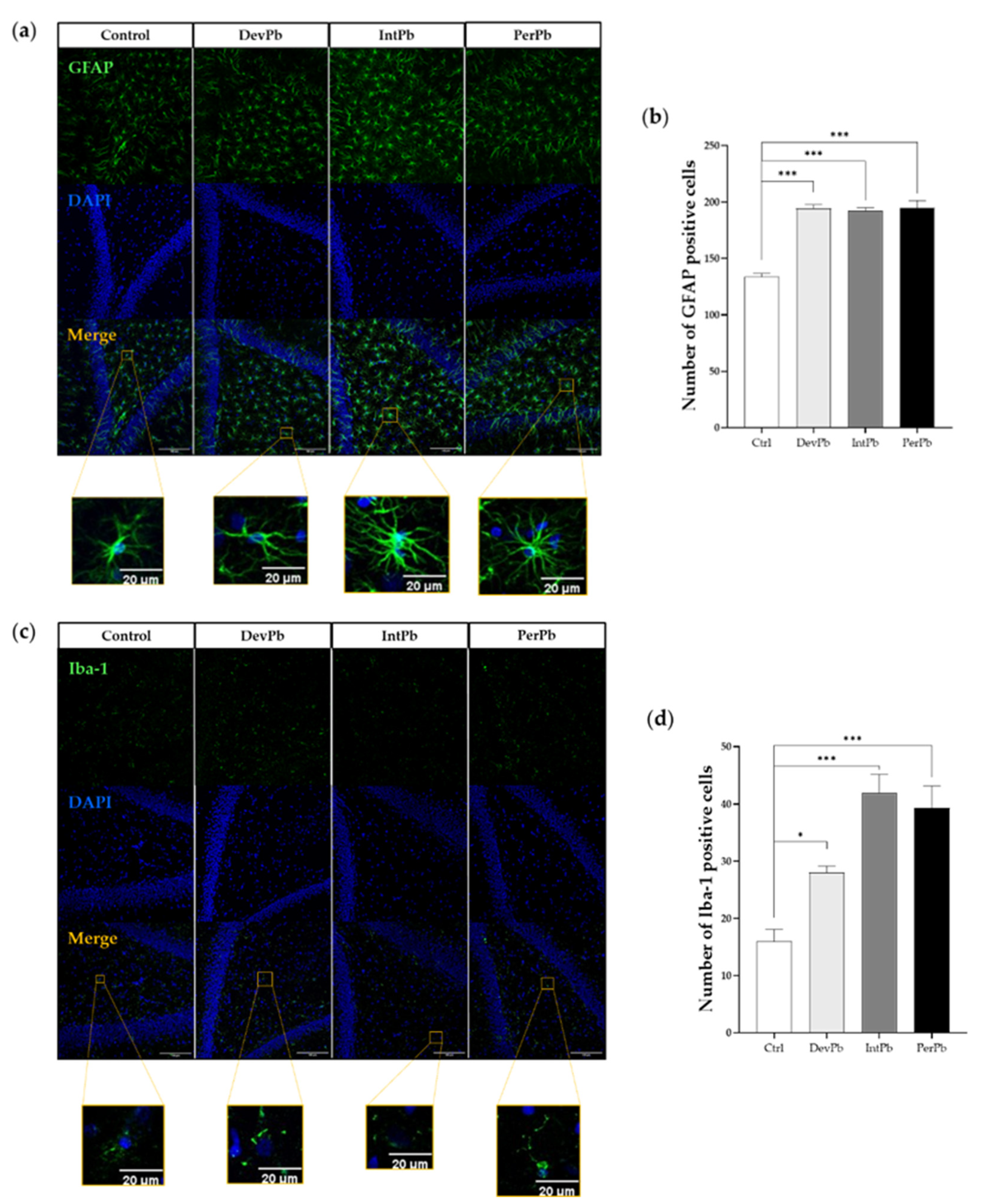
3.2.1. All Types of Lead Exposures Caused Strong Astrocytic and Microglial Activation
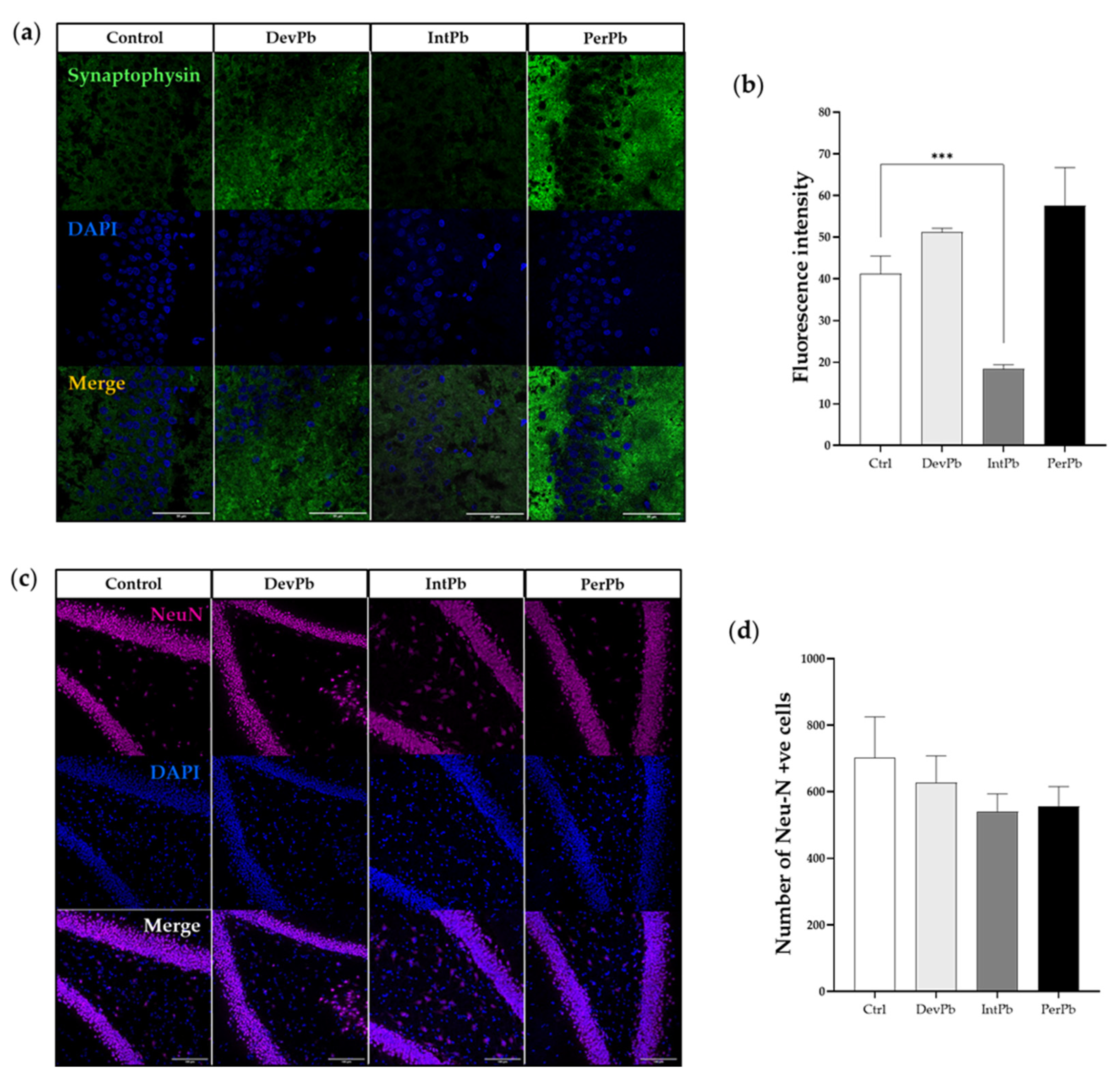
3.2.2. Intermittent Lead Exposure Caused Synaptic Loss, without Neuronal Degeneration
3.3. Lead Exposure from the Fetal Period until Adulthood Caused an Increase in Blood Lead Levels, with no Effects on Food and Liquid Intake or Feces and Urine Production
3.4. Lead Exposure from the Fetal Period until Adulthood Caused Hypertension Concomitant with Autonomic Dysfunction
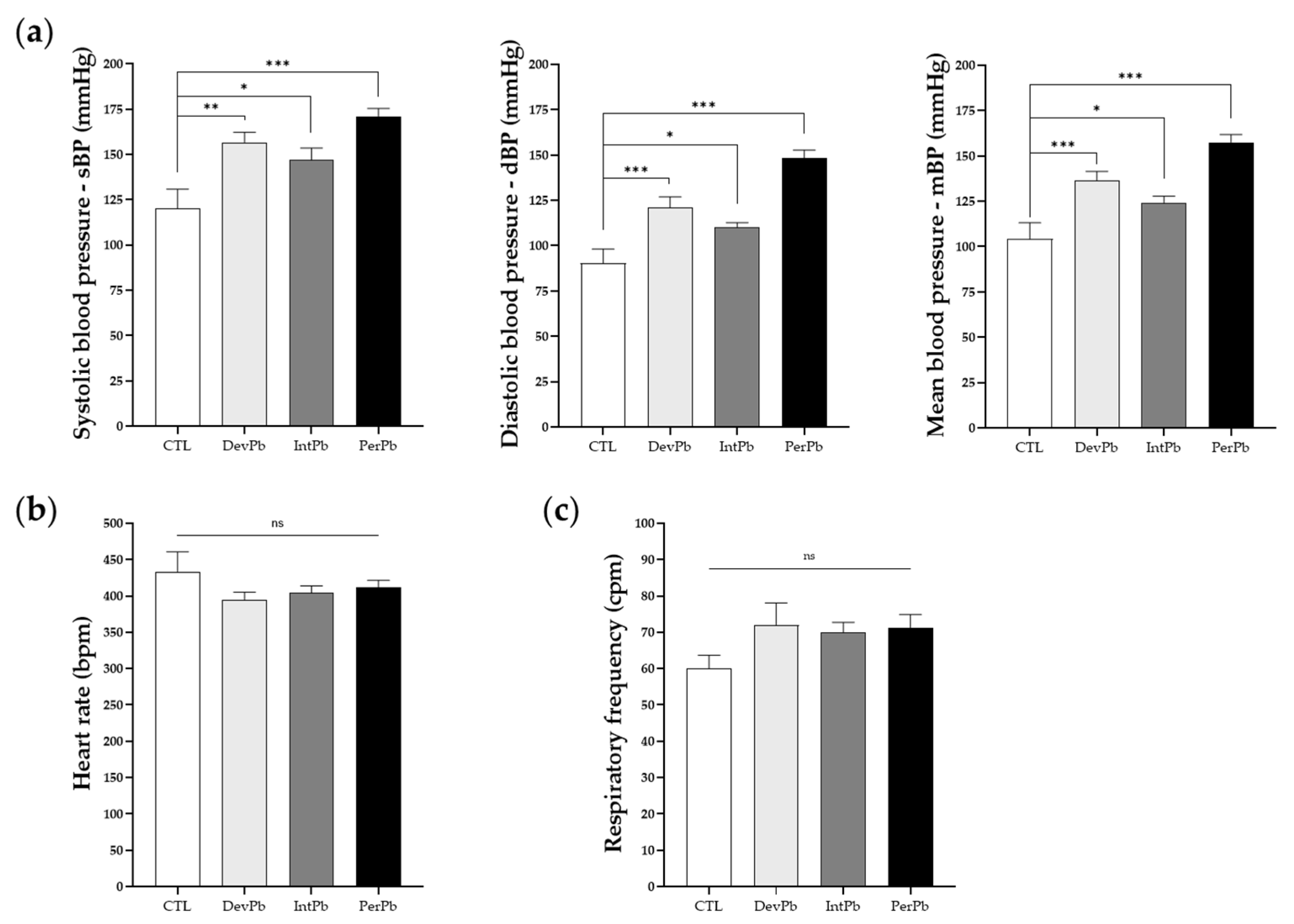
3.4.1. All Types of Lead Exposure Caused Hypertension without Heart Rate or Respiratory Alterations
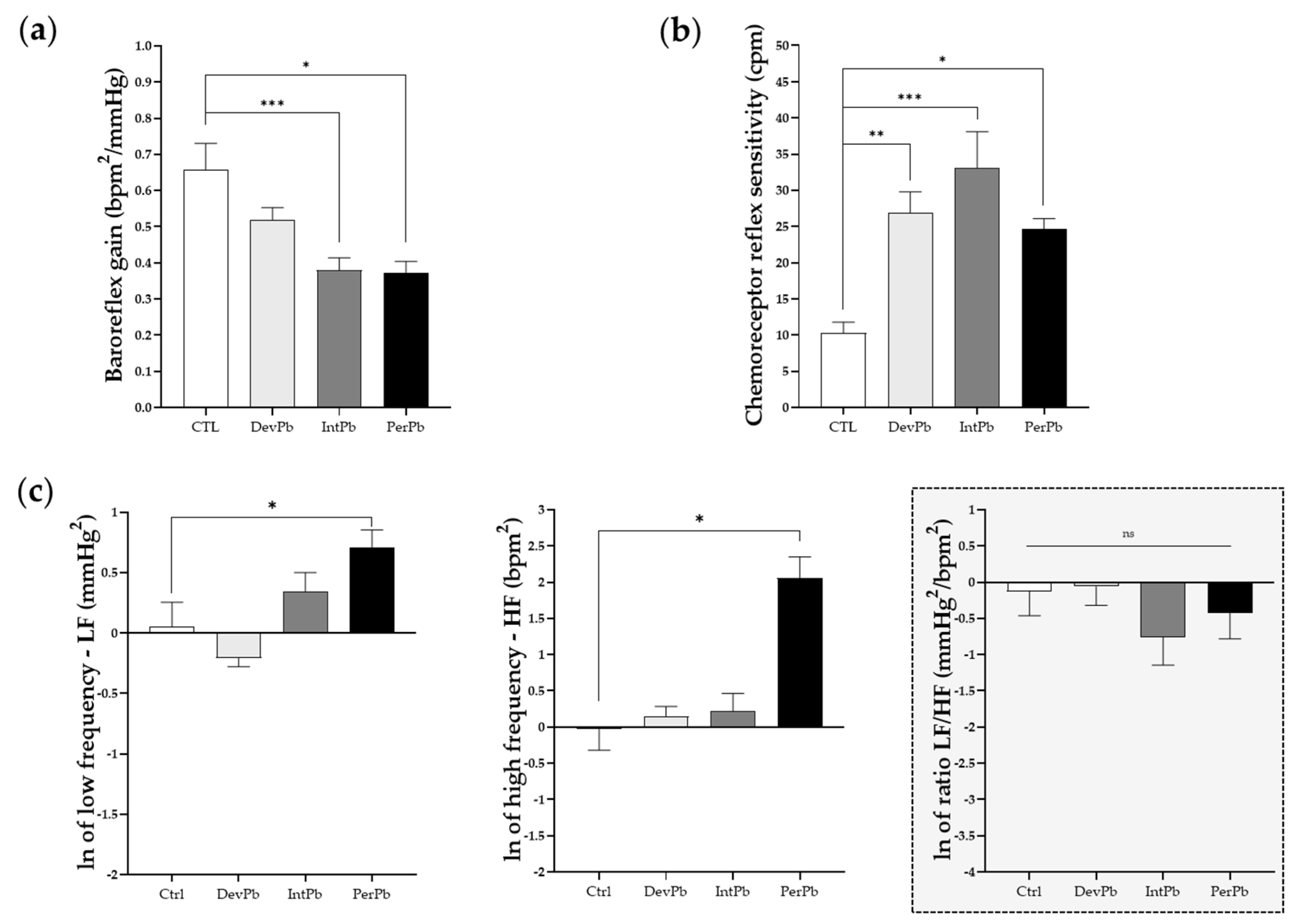
3.4.2. All Lead Exposures Caused Increased Chemoreceptor Reflex Sensitivity, Permanent Lead Exposure and Intermittent Lead Exposure Caused Strong Baroreflex Impairment, and Only the Permanent Lead Exposure Triggered Sympathetic and Parasympathetic Overexcitation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lead Poisoning. Available online: https://www.who.int/news-room/fact-sheets/detail/lead-poisoning-and-health (accessed on 24 May 2022).
- McQuirter, J.L.; Rothenberg, S.J.; Dinkins, G.A.; Kondrashov, V.; Manalo, M.; Todd, A.C.; Killgro, K.; Kosnett, M.J.; Wedeen, R.P.; Rothenberg, S.J.; et al. Lead and Lead Poisoning from Antiquity to Modern Times. Environ. Health Perspect. 2013, 115, 1–2. [Google Scholar] [CrossRef]
- Chemical Safety and Health. Available online: https://www.who.int/data/gho/data/themes/topics/indicator-groups/legally-binding-controls-on-lead-paint (accessed on 24 May 2022).
- Wani, A.L.; Ara, A.; Usmani, J.A. Lead Toxicity: A Review. Interdiscip. Toxicol. 2015, 8, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gidlow, D.A. Lead Toxicity. Occup. Med. 2015, 65, 348–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loghman-Adham, M. Renal Effects of Environmental and Occupational Lead Exposure. Environ. Health Perspect. 2008, 105, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Prokopowicz, A.; Sobczak, A.; Szula-Chraplewska, M.; Zaciera, M.; Kurek, J.; Szoltysek-Boldys, I. Effect of Occupational Exposure to Lead on New Risk Factors for Cardiovascular Diseases. Occup. Environ. Med. 2017, 74, 366–373. [Google Scholar] [CrossRef]
- Tong, S.; von Schirnding, Y.E.; Prapamontol, T. Environmental lead exposure: A public health problem of global dimensions. Bull. World Health Organ. 2000, 78, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Abadin, H.; Taylor, J.; Buser, M.; Scinicariello, F.; Przybyla, J.; Klotzbach, J.M.; Diamond, G.L.; Citra, M.; Chappell, L.L.; McIlroy, L.A.; et al. Toxicological Profile for Lead. In U.S. Public Health Service, Agency for Toxic Substances and Disease Registry; ATSDR: Atlanta, GA, USA, 2007; p. 582. [Google Scholar] [CrossRef]
- Flora, G.; Gupta, D.; Tiwari, A. Toxicity of lead: A review with recent updates. Interdiscip. Toxicol. 2012, 5, 47–58. [Google Scholar] [CrossRef]
- WHO. Lead Poisoning and Health; Fact Sheet; WHO: New York, NY, USA, 2015; Volume 379. [Google Scholar]
- Vassallo, D.; Simões, M.; Furieri, L.; Fioresi, M.; Fiorim, J.; Almeida, E.; Angeli, J.; Wiggers, G.; Peçanha, F.; Salaices, M. Toxic effects of mercury, lead and gadolinium on vascular reactivity. Braz. J. Med. Biol. Res. 2011, 44, 939–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assi, M.A.; Hezmee, M.N.M.; Haron, A.W.; Sabri, M.Y.M.; Rajion, M.A. The detrimental effects of lead on human and animal health. Veter. World 2016, 9, 660–671. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, M.R.; Madhu, P.; Reddy, K.P.; Reddy, S. The protective effects of zinc in lead-induced testicular and epididymal toxicity in Wistar rats. Toxicol. Ind. Health 2016, 33, 265–276. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, J.A.; Gump, B.B.; Roosa, K.; Bendinskas, K.; Dumas, A.; Morgan, R.; Parsons, P. Lead Exposure and Cardiovascular Dysregulation in Children. FASEB J. 2009, 23, 713–786. [Google Scholar] [CrossRef]
- Navas-Acien, A.; Guallar, E.; Silbergeld, E.K.; Rothenberg, S.J. Lead Exposure and Cardiovascular Disease—A Systematic Review. Environ. Health Perspect. 2007, 115, 472–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, M.R.; Preti, S.C.; Azevedo, B.F.; Fiorim, J.; Freire, D.D.; Covre, E.P.; Vassallo, D.V.; dos Santos, L. Low-level Chronic Lead Exposure Impairs Neural Control of Blood Pressure and Heart Rate in Rats. Cardiovasc. Toxicol. 2017, 17, 190–199. [Google Scholar] [CrossRef]
- Mason, L.H.; Harp, J.P.; Han, D.Y. Pb Neurotoxicity: Neuropsychological Effects of Lead Toxicity. BioMed Res. Int. 2014, 2014, 840547. [Google Scholar] [CrossRef] [Green Version]
- Toscano, C.D.; Guilarte, T.R. Lead neurotoxicity: From exposure to molecular effects. Brain Res. Rev. 2005, 49, 529–554. [Google Scholar] [CrossRef]
- Virgolini, M.B.; Aschner, M. Molecular mechanisms of lead neurotoxicity. Adv. Neurotoxicol. 2021, 5, 159–213. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.-S.; Hao, J.-H.; Zeng, Y.; Dai, F.-C.; Gu, P.-Q. Neurotoxicity and Biomarkers of Lead Exposure: A Review. Chin. Med. Sci. J. 2013, 28, 178–188. [Google Scholar] [CrossRef]
- Lidsky, T.I.; Schneider, J.S. Lead neurotoxicity in children: Basic mechanisms and clinical correlates. Brain 2003, 126, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Waseem, S.; Bahmani, A.; Subaiea, G.M.; Zawia, N.H. Infantile Exposure to Lead and Late-Age Cognitive Decline: Relevance to AD. Alzheimers Dement. 2014, 10, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Eid, A.; Bihaqi, S.W.; Renehan, W.E.; Zawia, N.H. Developmental lead exposure and lifespan alterations in epigenetic regulators and their correspondence to biomarkers of Alzheimer’s disease. Alzheimer’s Dementia Diagn. Assess. Dis. Monit. 2016, 2, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.; Triantafyllidou, S.; Best, D. Elevated Blood Lead in Young Children Due to Lead-Contaminated Drinking Water: Washington, DC, 2001−2004. Environ. Sci. Technol. 2009, 43, 1618–1623. [Google Scholar] [CrossRef] [PubMed]
- Njati, S.Y.; Maguta, M.M. Lead-based paints and children’s PVC toys are potential sources of domestic lead poisoning—A review. Environ. Pollut. 2019, 249, 1091–1105. [Google Scholar] [CrossRef]
- WHO. Childhood lead poisoning prevention. J. Am. Med. 2010, 89, 1129–1130. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. International Lead Poisoning Prevention Week of Action. 2018. Available online: https://www.who.int/ipcs/lead_campaign/en/ (accessed on 24 May 2022).
- Kuhlmann, A.C.; McGlothan, J.L.; Guilarte, T.R. Developmental lead exposure causes spatial learning deficits in adult rats. Neurosci. Lett. 1997, 233, 101–104. [Google Scholar] [CrossRef]
- Driscoll, L.L.; Strupp, B.J. Assessment of attention and inhibitory control in rodent developmental neurotoxicity studies. Neurotoxicol. Teratol. 2015, 52, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Moreira, E.G.; Vassilieff, I.; Vassilieff, V.S. Developmental lead exposure: Behavioral alterations in the short and long term. Neurotoxicol. Teratol. 2001, 23, 489–495. [Google Scholar] [CrossRef]
- Gilbert, M.E.; Mack, C.M.; Lasley, S.M. Chronic developmental lead exposure and hippocampal long-term potentiation: Biphasic dose-response relationship. Neurotoxicology 1999, 20, 71–82. [Google Scholar]
- Hu, F.; Xu, L.; Liu, Z.-H.; Ge, M.-M.; Ruan, D.-Y.; Wang, H.-L. Developmental Lead Exposure Alters Synaptogenesis through Inhibiting Canonical Wnt Pathway In Vivo and In Vitro. PLoS ONE 2014, 9, e101894. [Google Scholar] [CrossRef]
- Zawia, N.; Harry, G. Developmental Exposure to Lead Interferes with Glial and Neuronal Differential Gene Expression in the Rat Cerebellum. Toxicol. Appl. Pharmacol. 1996, 138, 43–47. [Google Scholar] [CrossRef]
- Gilbert, M.E.; Kelly, M.E.; Samsam, T.E.; Goodman, J.H. Chronic Developmental Lead Exposure Reduces Neurogenesis in Adult Rat Hippocampus but Does Not Impair Spatial Learning. Toxicol. Sci. 2005, 86, 365–374. [Google Scholar] [CrossRef] [Green Version]
- CDC. Childhood Lead Poisoning Prevention Program. National Surveillance Data (1997–2011); US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2014. Available online: http://www.cdc.gov/nceh/lead/data/national.htm (accessed on 24 May 2022).
- Mayans, L. Lead Poisoning in Children. Available online: https://www.aafp.org/afp/2019/0701/afp20190701p24.pdf (accessed on 24 May 2022).
- Shvachiy, L.; Geraldes, V.; Amaro-Leal, Â.; Rocha, I. Intermittent low-level lead exposure provokes anxiety, hypertension, autonomic dysfunction and neuroinflammation. NeuroToxicology 2018, 69, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, V.; Carvalho, M.; Goncalves-Rosa, N.; Tavares, C.; Laranjo, S.; Rocha, I. Lead toxicity promotes autonomic dysfunction with increased chemoreceptor sensitivity. NeuroToxicology 2016, 54, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, D.; Carvalho, M.L.; Geraldes, V.; Rocha, I.; Alves, L.C.; Santos, J.P. Lead in liver and kidney of exposed rats: Aging accumulation study. J. Trace Elements Med. Biol. 2012, 26, 285–290. [Google Scholar] [CrossRef]
- Onaolapo, O.J.; Naolapo, A.Y.O.; Josiah, T.; Osaku, M.; Akanji, O.O.; Abiodun, O.R. Elevated Plus Maze and Y-Maze Behavioral Effects of Subchronic, Oral Low Dose Monosodium Glutamate in Swiss Albino Mice. J. Pharm. Biol. Sci. 2012, 3, 21–27. [Google Scholar] [CrossRef]
- dos Santos Alves, R.; de Souza, A.S. The Open Field Test. Igarss 2014, 1–5. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2011, 13, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Buccafusco, J.J. Methods of Behavior Analysis in Neurosciences; CRC: Boca Raton, FL, USA, 2001; Volume 3. [Google Scholar] [CrossRef]
- Schneider, P.; Ho, Y.-J.; Spanagel, R.; Pawlak, C.R. A Novel Elevated Plus-Maze Procedure to Avoid the One-Trial Tolerance Problem. Front. Behav. Neurosci. 2011, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Shvachiy, L.; Geraldes, V.; Amaro-Leal, Â.; Rocha, I. Persistent Effects on Cardiorespiratory and Nervous Systems Induced by Long-Term Lead Exposure: Results from a Longitudinal Study. Neurotox. Res. 2020, 37, 857–870. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A. Animal models of anxiety: Do I need multiple tests? Trends Pharmacol. Sci. 2008, 29, 493–498. [Google Scholar] [CrossRef]
- Mouro, F.M.; Batalha, V.L.; Ferreira, D.G.; Coelho, J.E.; Baqi, Y.; Müller, C.E.; Lopes, L.V.; Ribeiro, J.A.; Sebastião, A.M. Chronic and acute adenosine A2A receptor blockade prevents long-term episodic memory disruption caused by acute cannabinoid CB1 receptor activation. Neuropharmacology 2017, 117, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Sun, Z.; Lee, T.; Fusco, F.R.; Kimble, T.D.; Meade, C.A.; Cuthbertson, S.; Reiner, A. A simple and sensitive antigen retrieval method for free-floating and slide-mounted tissue sections. J. Neurosci. Methods 1999, 93, 149–162. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hol, E.M.; Pekny, M. Glial fibrillary acidic protein (GFAP) and the astrocyte intermediate filament system in diseases of the central nervous system. Curr. Opin. Cell Biol. 2015, 32, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Diz, D.I.; Garcia-Espinosa, M.A.; Gallagher, P.E.; Ganten, D.; Ferrario, C.M.; Averill, D.B. Angiotensin-(1-7) and Baroreflex Function in Nucleus Tractus Solitarii of (mRen2)27 Transgenic Rats. J. Cardiovasc. Pharmacol. 2008, 51, 542. [Google Scholar] [CrossRef] [Green Version]
- Paton, J.F.R.; Wang, S.; Polson, J.W.; Kasparov, S. Signalling across the blood brain barrier by angiotensin II: Novel implications for neurogenic hypertension. J. Mol. Med. 2008, 86, 705–710. [Google Scholar] [CrossRef]
- Wildemann, T.M.; Siciliano, S.D.; Weber, L.P. The mechanisms associated with the development of hypertension after exposure to lead, mercury species or their mixtures differs with the metal and the mixture ratio. Toxicology 2016, 339, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rocha, I.; Rosário, L.B.; de Oliveira, E.I.; Barros, M.A.; Silva-Carvallho, L. Enhancement of carotid chemoreceptor reflex and cardiac chemosensitive reflex in the acute phase of myocardial infarction of the anesthetized rabbit. Basic Res. Cardiol. 2003, 98, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, V.; Goncalves-Rosa, N.; Liu, B.; Paton, J.; Rocha, I. Chronic depression of hypothalamic paraventricular neuronal activity produces sustained hypotension in hypertensive rats. Exp. Physiol. 2013, 99, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, V.; Goncalves-Rosa, N.; Liu, B.; Paton, J.F.; Rocha, I. Essential role of RVL medullary neuronal activity in the long term maintenance of hypertension in conscious SHR. Auton. Neurosci. 2014, 186, 22–31. [Google Scholar] [CrossRef]
- De Almeida Lopes, A.C.B.; Silbergeld, E.K.; Navas-Acien, A.; Zamoiski, R.; da Cunha Martins, A., Jr.; Camargo, A.E.I.; Urbano, M.; Mesas, A.; Paoliello, M.M.B. Association between blood lead and blood pressure: A population-based study in Brazilian adults. Environ. Health 2017, 16, 27. [Google Scholar] [CrossRef] [Green Version]
- Simões, M.R.; Ribeiro Júnior, R.F.; Vescovi, M.V.A.; De Jesus, H.C.; Padilha, A.S.; Stefanon, I.; Vassallo, D.V.; Salaices, M.; Fioresi, M. Acute Lead Exposure Increases Arterial Pressure: Role of the Renin-Angiotensin System. PLoS ONE 2011, 6, e18730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boscolo, P.; Carmignani, M. Neurohumoral blood pressure regulation in lead exposure. Environ. Health Perspect. 1988, 78, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Farmand, F.; Ehdaie, A.; Roberts, C.K.; Sindhu, R.K. Lead-induced dysregulation of superoxide dismutases, catalase, glutathione peroxidase, and guanylate cyclase. Environ. Res. 2005, 98, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Wilbur, S.B. Health effects classification and its role in the derivation of minimal risk levels: Respiratory effects. Clean Technol. Environ. Toxicol. Occup. Med. 1998, 7, 233. [Google Scholar]
- Roncal, C.; Mu, W.; Reungjui, S.; Kim, K.M.; Henderson, G.N.; Ouyang, X.; Nakagawa, T.; Johnson, R.J. Lead, at low levels, accelerates arteriolopathy and tubulointerstitial injury in chronic kidney disease. Am. J. Physiol. Physiol. 2007, 293, F1391–F1396. [Google Scholar] [CrossRef] [PubMed]
- Trzebski, A.; Tafil, M.; Zoltowski, M.; Przybylski, J. Central and peripheral chemosensitivity in early essential hypertension in man. In Central Neurone Environment and the Control Systems of Breathing and Circulation; Springer: Berlin/Heidelberg, Germany, 1983; pp. 204–213. [Google Scholar]
- Kara, T.; Narkiewicz, K.; Somers, V.K. Chemoreflexes—Physiology and clinical implications. Acta Physiol. Scand. 2003, 177, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Prabhakar, N.R.; Prem, K. Peripheral chemoreceptors Function and. Compr. Physiol. 2012, 2, 141–219. [Google Scholar] [CrossRef]
- Prabhakar, N.R.; Peng, Y.-J. Peripheral chemoreceptors in health and disease. J. Appl. Physiol. 2004, 96, 359–366. [Google Scholar] [CrossRef]
- Fisher, J.P.; Paton, J.F.R. The sympathetic nervous system and blood pressure in humans: Implications for hypertension. J. Hum. Hypertens. 2012, 26, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Kasparov, S.; Butcher, J.W.; Paton, J.F. Angiotensin II receptors within the nucleus of the solitary tract mediate the developmental attenuation of the baroreceptor vagal reflex in pre-weaned rats. J. Auton. Nerv. Syst. 1998, 74, 160–168. [Google Scholar] [CrossRef]
- Swenne, C.A. Baroreflex sensitivity: Mechanisms and measurement. Neth. Heart J. 2012, 21, 58–60. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, M.; Parati, G.; Castiglioni, P.; Tordi, R.; Mancia, G.; Pedotti, A. Baroreflex effectiveness index: An additional measure of baroreflex control of heart rate in daily life. Am. J. Physiol. Integr. Comp. Physiol. 2001, 280, R744–R751. [Google Scholar] [CrossRef] [Green Version]
- Albaghdadi, M. Baroreflex Control of Long-Term Arterial Pressure. Rev Bras Hipertens 2007, 14, 212–225. [Google Scholar]
- Obisesan, T.O. Hypertension and Cognitive Function. Clin. Geriatr. Med. 2009, 25, 259–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aletti, F.; Gambarotta, N.; Penn, A.H.; Ferrario, M.; Schmid-Schönbein, G.W. Heart period and blood pressure characteristics in splanchnic arterial occlusion shock-induced collapse. Int. J. Clin. Monit. Comput. 2015, 31, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patwa, J.; Flora, S.J.S. Heavy Metal-Induced Cerebral Small Vessel Disease: Insights into Molecular Mechanisms and Possible Reversal Strategies. Int. J. Mol. Sci. 2020, 21, 3862. [Google Scholar] [CrossRef]
- Selvin-Tesla, A.; Lopez-Costa, J.J.; de Avinon, A.C.N.; Saavedra, J.P. Astroglial alterations in rat hippocampus during chronic lead exposure. Glia 1991, 4, 384–392. [Google Scholar] [CrossRef]
- Liu, J.T.; Dong, M.H.; Zhang, J.Q.; Bai, Y.; Kuang, F.; Chen, L.W. Microglia and astroglia: The role of neuroinflammation in lead toxicity and neuronal injury in the brain. Neuroimmunol. Neuroinflamm. 2015, 2, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, L.S.; Bird, L.; Legare, M.E.; Mikeska, G.; Bratton, G.R.; Tiffany-Castiglioni, E. Differential ability of astroglia and neuronal cells to accumulate lead: Dependence on cell type and on degree of differentiation. Toxicol. Sci. 1999, 50, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghurst, P.A.; McMichael, A.J.; Wigg, N.R.; Vimpani, G.V.; Robertson, E.F.; Roberts, R.J.; Tong, S.L. Environmental exposure to lead and children’s intelligence at the age of seven years: The Port Pirie Cohort Study. N. Engl. J. Med. 1992, 327, 1279–1284. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, Y.; Ni, L.; Zhao, S.; Li, L.; Zhang, J.; Fan, M.; Liang, C.; Cao, J.; Xu, L. Lead exposure through gestation-only caused long-term learning/memory deficits in young adult offspring. Exp. Neurol. 2003, 184, 489–495. [Google Scholar] [CrossRef]
- Lasley, S.M.; Gilbert, M.E. Impairment of Synaptic Function by Exposure to Lead. Vitr. Neurotoxicol. 2004, 217–241. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Blood Lead Levels (μg/dL) | Weight (g) | Food Intake (g) | Liquid Intake (mL) | Produced Feces (g) | Produced Urine (mL) |
|---|---|---|---|---|---|---|
| Ctrl | 0.6 ± 0.1 | 333± 41.0 | 22.8 ± 2.0 | 26.2 ± 1.8 | 10.8 ± 1.5 | 8.0 ± 0.6 |
| DevPb | 3.6 ± 0.4 ns | 445 ± 53.7 ns | 23.2 ± 2.0 ns | 31.2 ± 2.5 ns | 16.8 ± 1.0 ns | 8.7 ± 0.9 ns |
| IntPb | 18.4 ± 1.7 ns | 428 ± 50.2 ns | 24.0 ± 1.6 ns | 25.0 ± 2.2 ns | 10.8 ± 0.4 ns | 9.3 ± 2.2 ns |
| PerPb | 26.9 ± 2.2 ns | 434 ± 50.4 ns | 23.5 ± 2.8 ns | 29.6 ± 2.0 ns | 16.4 ± 2.6 ns | 7.5 ± 1.1 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shvachiy, L.; Amaro-Leal, Â.; Outeiro, T.F.; Rocha, I.; Geraldes, V. From Molecular to Functional Effects of Different Environmental Lead Exposure Paradigms. Biology 2022, 11, 1164. https://doi.org/10.3390/biology11081164
Shvachiy L, Amaro-Leal Â, Outeiro TF, Rocha I, Geraldes V. From Molecular to Functional Effects of Different Environmental Lead Exposure Paradigms. Biology. 2022; 11(8):1164. https://doi.org/10.3390/biology11081164
Chicago/Turabian StyleShvachiy, Liana, Ângela Amaro-Leal, Tiago F. Outeiro, Isabel Rocha, and Vera Geraldes. 2022. "From Molecular to Functional Effects of Different Environmental Lead Exposure Paradigms" Biology 11, no. 8: 1164. https://doi.org/10.3390/biology11081164
APA StyleShvachiy, L., Amaro-Leal, Â., Outeiro, T. F., Rocha, I., & Geraldes, V. (2022). From Molecular to Functional Effects of Different Environmental Lead Exposure Paradigms. Biology, 11(8), 1164. https://doi.org/10.3390/biology11081164