
Comparative Genomics Insights into a Novel Biocontrol Agent Paenibacillus peoriae Strain ZF390 against Bacterial Soft Rot



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation, Growth Conditions, Antagonism Assays, and Biocontrol Assays
2.2. Characteristics, Microscopic Analysis and Genomic DNA Extraction
2.3. Genome Sequencing and Annotation
2.4. Phylogenetic Analysis and Genome Comparisons
2.5. Analysis of Gene Clusters for Biosynthesis of Secondary Metabolites
2.6. Genome Mining for Genes Encoding Plant Beneficial Traits
2.7. Statistical Analysis
3. Results
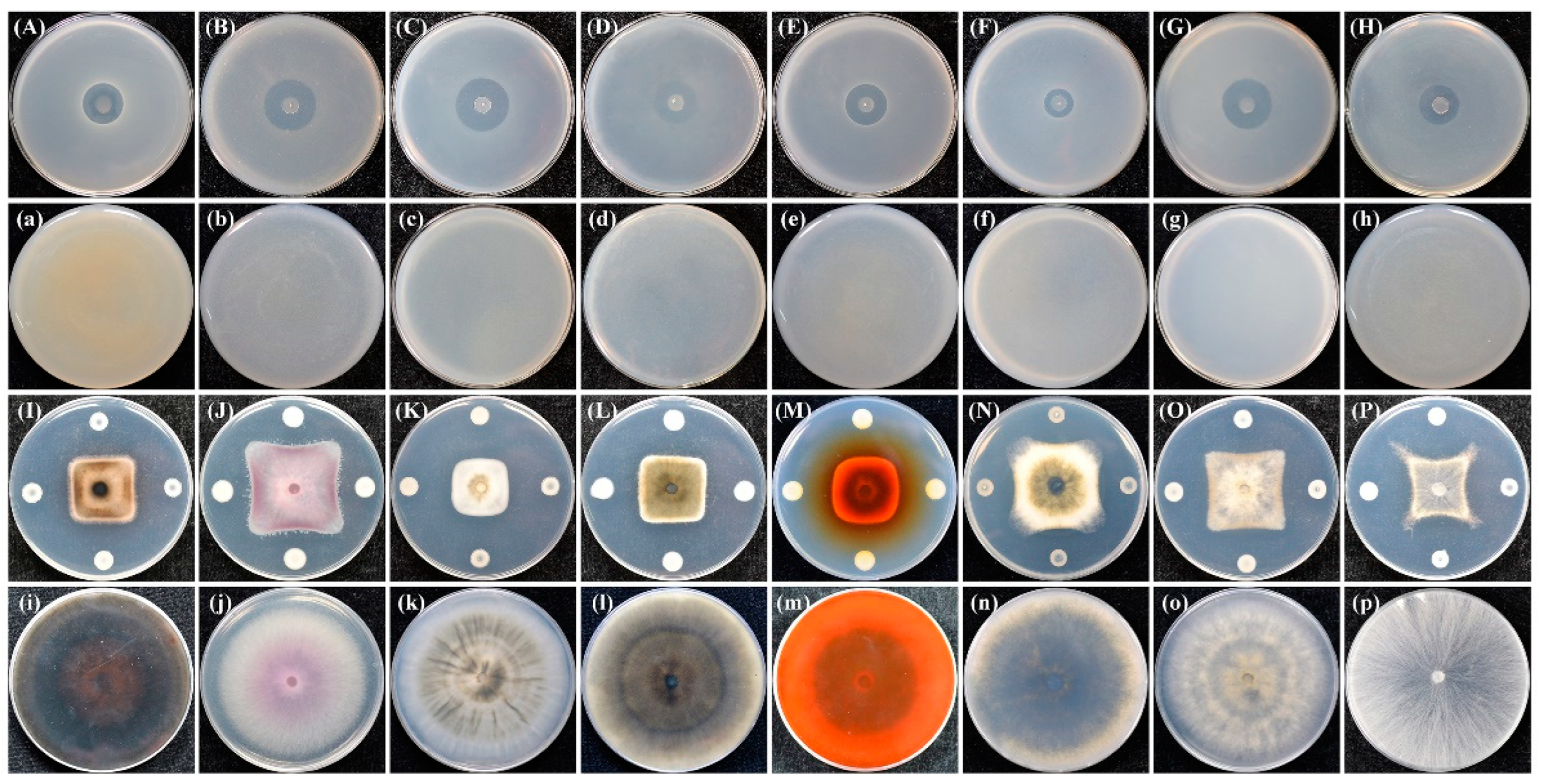
3.1. Biocontrol Effect of Strain ZF390 against Pbr
3.2. Organism Information
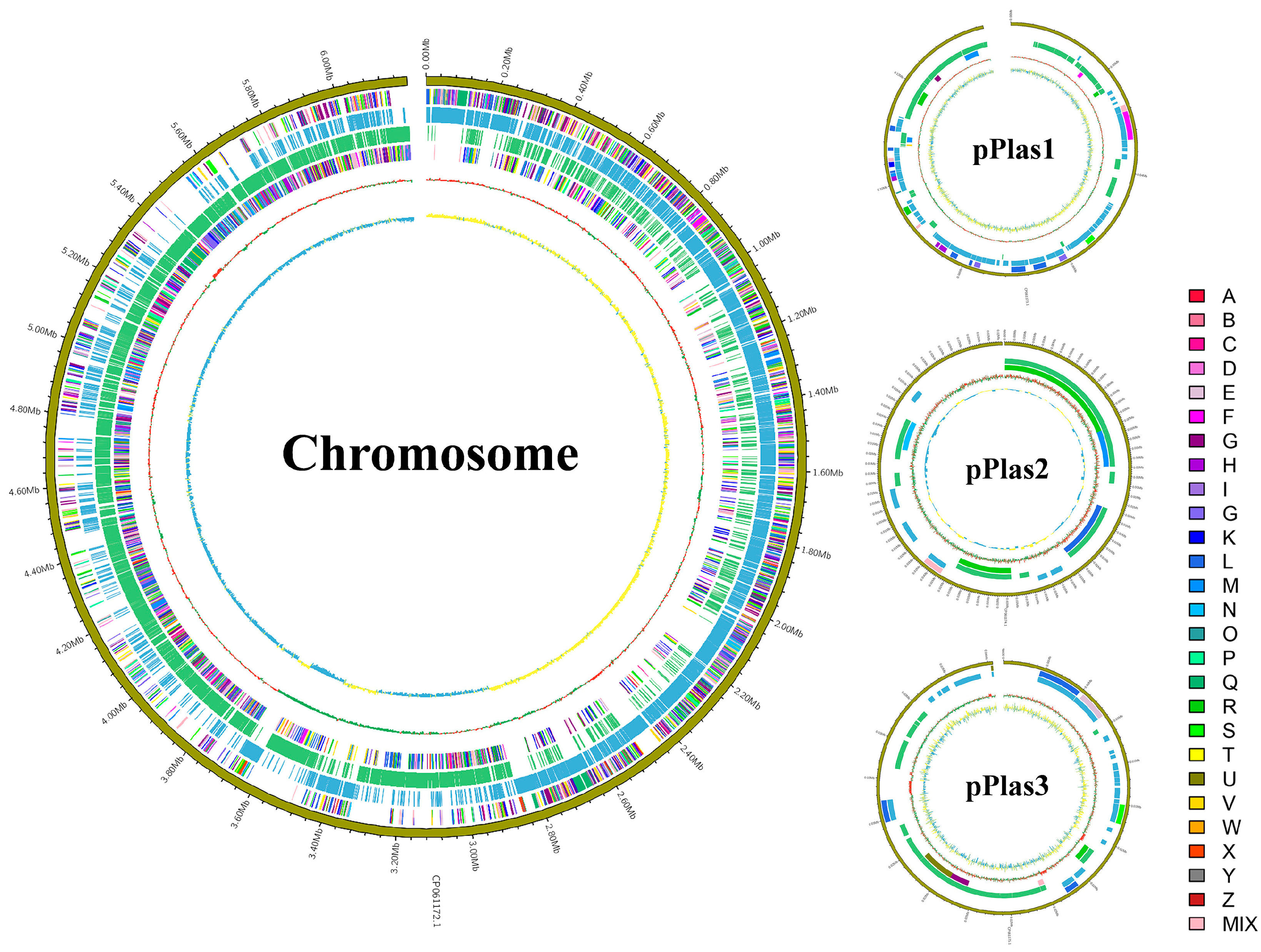
3.3. General Genomic Features of P. peoriae ZF390
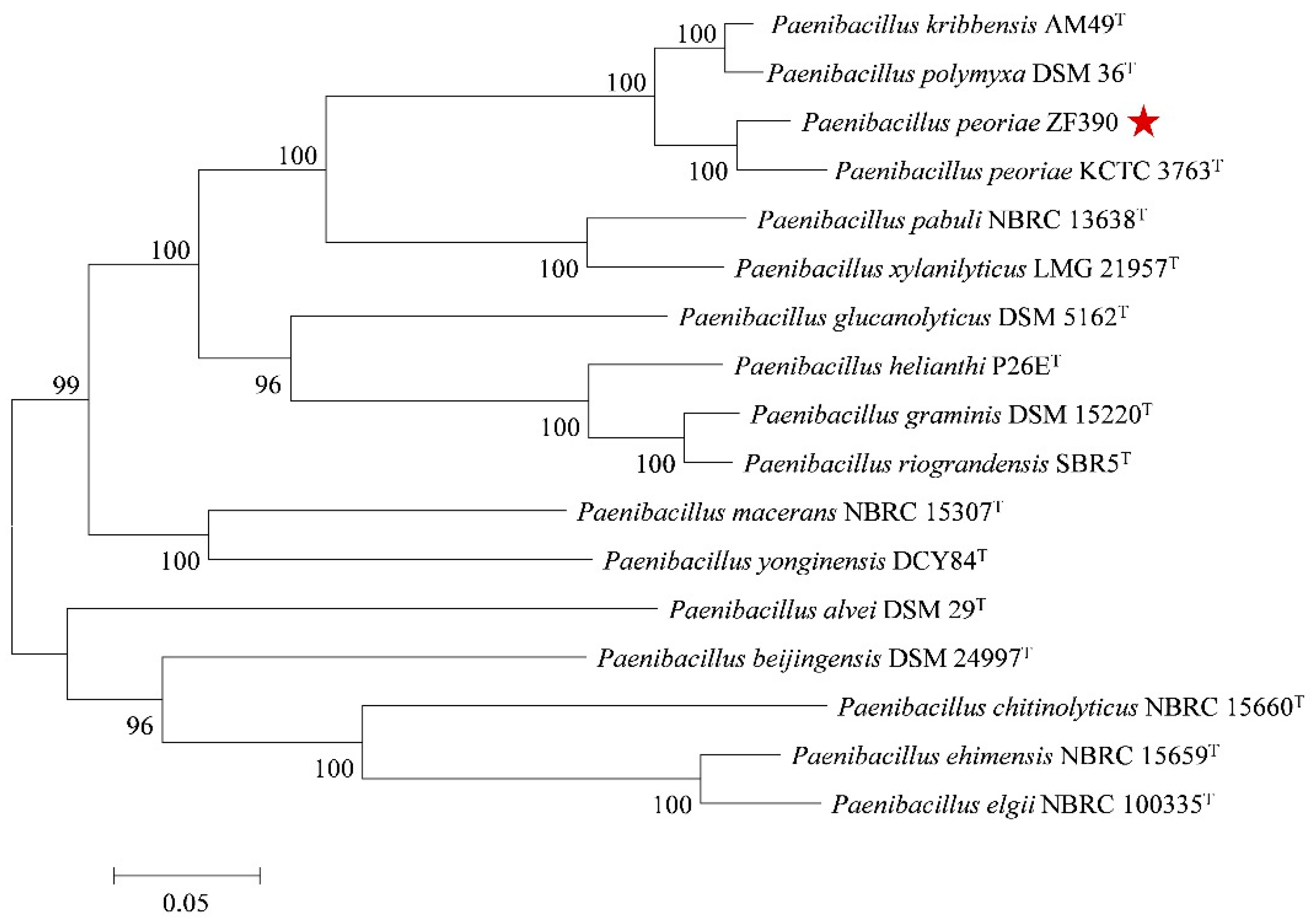
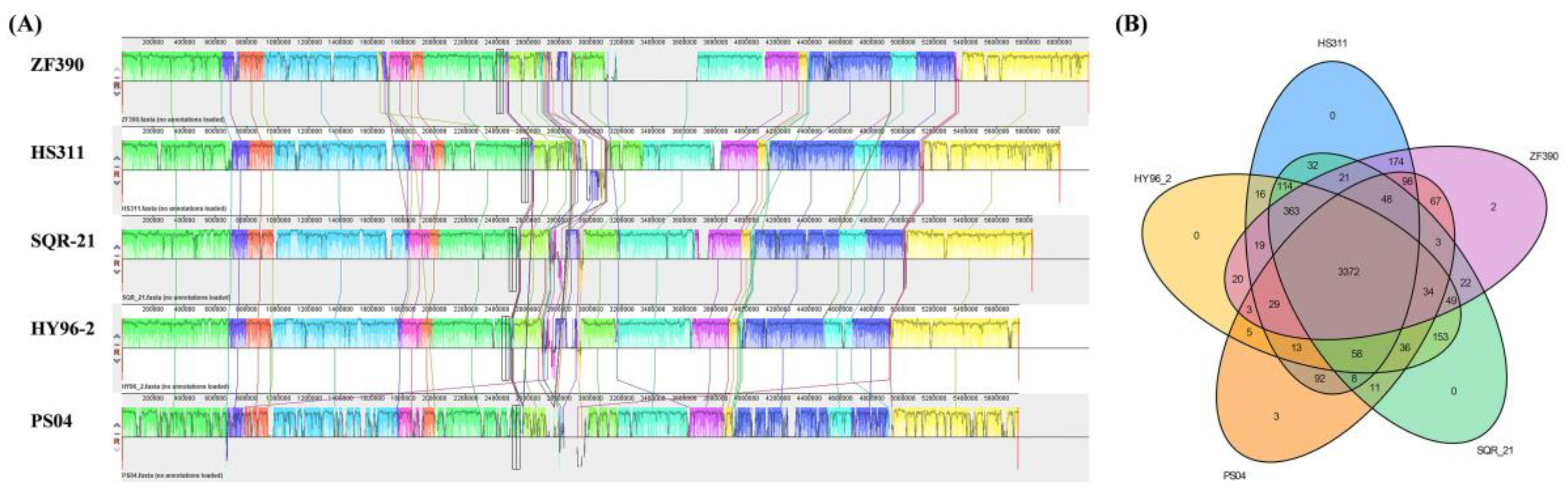
3.4. Comparative Genomics Analysis of P. peoriae ZF390 with Other Paenibacillus Strains
3.5. Gene Clusters Involved in the Synthesis of Secondary Metabolites
3.6. Mining for Functional Genes Potentially Associated with Plant–Bacteria Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portier, P.; Pédron, J.; Taghouti, G.; Fischer-Le Saux, M.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum and description of Pectobacterium versatile sp. nov., isolated from streams and symptoms on diverse plants. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar] [PubMed]
- Oulghazi, S.; Sarfraz, S.; Zaczek-Moczydłowska, M.A.; Khayi, S.; Ed-Dra, A.; Lekbach, Y.; Campbell, K.; Moleleki, L.N.; O’Hanlon, R.; Faure, D. Pectobacterium brasiliense: Genomics, Host Range and Disease Management. Microorganisms 2021, 9, 106. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Chai, A.; Shi, Y.; Xie, X.; Ma, Z.; Li, B. Emergence of Bacterial Soft Rot in Cucumber Caused by Pectobacterium carotovorum subsp. brasiliense in China. Plant Dis. 2017, 101, 279–287. [Google Scholar] [CrossRef] [Green Version]
- van der Wolf, J.M.; de Boer, S.H.; Czajkowski, R.L.; Cahill, G.; van Gijsegem, F.; Davey, T.; Dupuis, B.; Ellicott, J.; Jafra, S.; Kooman, M.; et al. Management of Diseases Caused by Pectobacterium and Dickeya Species. In Plant Diseases Caused by Dickeya and Pectobacterium Species; van Gijsegem, F., van der Wolf, J.M., Toth, I.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 175–214. [Google Scholar]
- Hashem, A.; Tabassum, B.; Fathi Abd_Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.; Zou, C.; Xu, X.; Ke, Z.; Hou, J.; Jiang, G.; Fan, C.; Gong, J.; Wei, J. Complete genome sequence of biocontrol strain Paenibacillus peoriae HJ-2 and further analysis of its biocontrol mechanism. BMC Genom. 2022, 23, 161. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, N.; Cao, M.; Wu, K.; Shen, Q.; Huang, Q. Biological control of tobacco black shank and colonization of tobacco roots by a Paenibacillus polymyxa strain C5. Biol. Fertil. Soils 2012, 48, 613–620. [Google Scholar] [CrossRef]
- Abdallah, Y.; Yang, M.; Zhang, M.; Masum, M.M.I.; Ogunyemi, S.O.; Hossain, A.; An, Q.; Yan, C.; Li, B. Plant growth promotion and suppression of bacterial leaf blight in rice by Paenibacillus polymyxa Sx3. Lett. Appl. Microbiol. 2019, 68, 423–429. [Google Scholar] [CrossRef]
- Chávez-Ramírez, B.; Rodríguez-Velázquez, N.D.; Mondragón-Talonia, C.M.; Avendaño-Arrazate, C.H.; Martínez-Bolaños, M.; Vásquez-Murrieta, M.S.; Estrada de Los Santos, P. Paenibacillus polymyxa NMA1017 as a potential biocontrol agent of Phytophthora tropicalis, causal agent of cacao black pod rot in Chiapas, Mexico. Antonie Van Leeuwenhoek 2021, 114, 55–68. [Google Scholar] [CrossRef]
- Zhai, Y.; Zhu, J.X.; Tan, T.M.; Xu, J.P.; Shen, A.R.; Yang, X.B.; Li, J.L.; Zeng, L.B.; Wei, L. Isolation and characterization of antagonistic Paenibacillus polymyxa HX-140 and its biocontrol potential against Fusarium wilt of cucumber seedlings. BMC Microbiol. 2021, 21, 75. [Google Scholar] [CrossRef]
- Soni, R.; Keharia, H. Phytostimulation and biocontrol potential of Gram-positive endospore-forming Bacilli. Planta 2021, 254, 49. [Google Scholar] [CrossRef] [PubMed]
- Langendries, S.; Goormachtig, S. Paenibacillus polymyxa, a Jack of all trades. Environ. Microbiol. 2021, 23, 5659–5669. [Google Scholar] [CrossRef]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Ramírez, B.; Kerber-Díaz, J.C.; Acoltzi-Conde, M.C.; Ibarra, J.A.; Vásquez-Murrieta, M.S.; Estrada-de Los Santos, P. Inhibition of Rhizoctonia solani RhCh-14 and Pythium ultimum PyFr-14 by Paenibacillus polymyxa NMA1017 and Burkholderia cenocepacia CACua-24: A proposal for biocontrol of phytopathogenic fungi. Microbiol. Res. 2020, 230, 126347. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Wu, H.; Wang, W.; Yang, Y.; Xie, S.; Xie, Y.; Gao, X. Efficient colonization and harpins mediated enhancement in growth and biocontrol of wilt disease in tomato by Bacillus subtilis. Lett. Appl. Microbiol. 2013, 57, 526–533. [Google Scholar] [CrossRef]
- Allard-Massicotte, R.; Tessier, L.; Lécuyer, F.; Lakshmanan, V.; Lucier, J.F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis Early Colonization of Arabidopsis thaliana Roots Involves Multiple Chemotaxis Receptors. mBio 2016, 7, e01664-16. [Google Scholar] [CrossRef] [Green Version]
- Al-Ali, A.; Deravel, J.; Krier, F.; Béchet, M.; Ongena, M.; Jacques, P. Biofilm formation is determinant in tomato rhizosphere colonization by Bacillus velezensis FZB42. Environ. Sci. Pollut. Res. Int. 2018, 25, 29910–29920. [Google Scholar] [CrossRef]
- Zeriouh, H.; de Vicente, A.; Pérez-García, A.; Romero, D. Surfactin triggers biofilm formation of Bacillus subtilis in melon phylloplane and contributes to the biocontrol activity. Environ. Microbiol. 2014, 16, 2196–2211. [Google Scholar] [CrossRef]
- Gao, S.; Wu, H.; Yu, X.; Qian, L.; Gao, X. Swarming motility plays the major role in migration during tomato root colonization by Bacillus subtilis SWR01. Biol. Control 2016, 98, 11–17. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Franco-Sierra, N.D.; Posada, L.F.; Santa-María, G.; Romero-Tabarez, M.; Villegas-Escobar, V.; Álvarez, J.C. Bacillus subtilis EA-CB0575 genome reveals clues for plant growth promotion and potential for sustainable agriculture. Funct. Integr. Genomics 2020, 20, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Sulthana, A.; Lakshmi, S.G.; Madempudi, R.S. Genome Sequencing and Annotation of Bacillus subtilis UBBS-14 to Ensure Probiotic Safety. J. Genomics 2019, 7, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yuan, J.; Zhang, J.; Shen, Z.; Zhang, M.; Li, R.; Ruan, Y.; Shen, Q. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fertil. Soils 2012, 49, 435–446. [Google Scholar] [CrossRef]
- Xu, S.; Xie, X.; Zhao, Y.; Shi, Y.; Chai, A.; Li, L.; Li, B. Whole-genome analysis of bacillus velezensis ZF2, a biocontrol agent that protects cucumis sativus against corynespora leaf spot diseases. 3 Biotech 2020, 10, 186. [Google Scholar] [CrossRef]
- Yuan, L.; Li, L.; Zheng, F.; Shi, Y.; Xie, X.; Chai, A.; Li, B. The complete genome sequence of Rahnella aquatilis ZF7 reveals potential beneficial properties and stress tolerance capabilities. Arch. Microbiol. 2020, 202, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, F.H.; Ambrosini, A.; de Souza, R.; de Carvalho Fernandes, G.; Bach, E.; Balsanelli, E.; Baura, V.; Brito, L.F.; Wendisch, V.F.; de Oliveira Pedrosa, F.; et al. Reclassification of Paenibacillus riograndensis as a Genomovar of Paenibacillus sonchi: Genome-Based Metrics Improve Bacterial Taxonomic Classification. Front. Microbiol. 2017, 8, 1849. [Google Scholar] [CrossRef]
- Heyndrickx, M.; Vandemeulebroecke, K.; Scheldeman, P.; Kersters, K.; de Vos, P.; Logan, N.A.; Aziz, A.M.; Ali, N.; Berkeley, R.C. A Polyphasic Reassessment of the Genus Paenibacillus, Reclassification of Bacillus lautus (Nakamura 1984) as Paenibacillus lautus comb. nov. and of Bacillus peoriae (Montefusco et al. 1993) as Paenibacillus peoriae comb. nov., and Emended Descriptions of P. lautus and of P. peoriae. Int. J. Syst. Bacteriol. 1996, 46, 988–1003. [Google Scholar] [PubMed]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Nawrocki, E.P.; Kolbe, D.L.; Eddy, S.R. Infernal 1.0: Inference of RNA alignments. Bioinformatics 2009, 25, 1335–1337. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Jaroszewski, L.; Godzik, A. Tolerating some redundancy significantly speeds up clustering of large protein databases. Bioinformatics 2002, 18, 77–82. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 2015, 43, D261–D269. [Google Scholar] [CrossRef]
- Saier, M.H., Jr.; Reddy, V.S.; Tsu, B.V.; Ahmed, M.S.; Li, C.; Moreno-Hagelsieb, G. The Transporter Classification Database (TCDB): Recent advances. Nucleic Acids Res. 2016, 44, D372–D379. [Google Scholar] [CrossRef] [Green Version]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- You, Z.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids. Res. 2011, 39, W347–W352. [Google Scholar]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids. Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Y.; Shen, P.; Lin, M.; Ye, X. Exogenous Alanine Reverses the Bacterial Resistance to Zhongshengmycin with the Promotion of the P Cycle in Xanthomonas oryzae. Antibiotics 2022, 11, 245. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Feng, Y.; Zong, Z. Precise Species Identification for Enterobacter: A Genome Sequence-Based Study with Reporting of Two Novel Species, Enterobacter quasiroggenkampii sp. nov. and Enterobacter quasimori sp. nov. mSystems 2020, 5, e00527-20. [Google Scholar] [CrossRef] [PubMed]
- Zong, Z. Genome-based Taxonomy for Bacteria: A Recent Advance. Trends Microbiol. 2020, 28, 871–874. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Hegemann, J.D.; Fage, C.D.; Zimmermann, M.; Xie, X.; Linne, U.; Marahiel, M.A. Insights into the Unique Phosphorylation of the Lasso Peptide Paeninodin. J. Biol. Chem. 2016, 291, 13662–13678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballantine, R.D.; McCallion, C.E.; Nassour, E.; Tokajian, S.; Cochrane, S.A. Tridecaptin-inspired antimicrobial peptides with activity against multidrug-resistant Gram-negative bacteria. Med. Chem. Commun. 2019, 10, 484–487. [Google Scholar] [CrossRef]
- Jangra, M.; Kaur, M.; Tambat, R.; Rana, R.; Maurya, S.K.; Khatri, N.; Ghafur, A.; Nandanwar, H. Tridecaptin M, a New Variant Discovered in Mud Bacterium, Shows Activity against Colistin- and Extremely Drug-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2019, 63, e00338-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Kuipers, O.P. BrevicidineB, a New Member of the Brevicidine Family, Displays an Extended Target Specificity. Front. Microbiol. 2021, 12, 693117. [Google Scholar]
- Park, J.E.; Kim, H.R.; Park, S.Y.; Choi, S.K.; Park, S.H. Identification of the biosynthesis gene cluster for the novel lantibiotic paenilan from Paenibacillus polymyxa E681 and characterization of its product. J. Appl. Microbiol. 2017, 123, 1133–1147. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, Y.; Kaneda, M. Fusaricidins B, C and D, new depsipeptide antibiotics produced by Bacillus polymyxa KT-8: Isolation, structure elucidation and biological activity. J. Antibiot. 1997, 50, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Yang, K.; Yuan, B.; Gong, B.; Wan, L.; Patil, N.A.; Swarbrick, J.D.; Roberts, K.D.; Schreiber, F.; Wang, L.; et al. Simulations of octapeptin–outer membrane interactions reveal conformational flexibility is linked to antimicrobial potency. J. Biol. Chem. 2020, 295, 15902–15912. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Cheng, Y.; Yi, J.; Zhang, Z.; Luo, Q.; Zhang, D.; Li, Y. Complete Genome Sequence of Industrial Biocontrol Strain Paenibacillus polymyxa HY96-2 and Further Analysis of Its Biocontrol Mechanism. Front. Microbiol. 2018, 9, 1520. [Google Scholar] [CrossRef]
- Baz, M.; Lahbabi, D.; Samri, S.; Val, F.; Hamelin, G.; Madore, I.; Bouarab, K.; Beaulieu, C.; Ennaji, M.M.; Barakate, M. Control of potato soft rot caused by Pectobacterium carotovorum and Pectobacterium atrosepticum by Moroccan actinobacteria isolates. World J. Microbiol. Biotechnol. 2012, 28, 303–311. [Google Scholar] [CrossRef]
- Charkowski, A.O. Biology and control of Pectobacterium in potato. Am. J. Potato Res. 2015, 92, 223–229. [Google Scholar] [CrossRef]
- Raoul des Essarts, Y.; Cigna, J.; Quêtu-Laurent, A.; Caron, A.; Munier, E.; Beury-Cirou, A.; Hélias, V.; Faure, D. Biocontrol of the Potato Blackleg and Soft Rot Diseases Caused by Dickeya dianthicola. Appl. Environ. Microbiol. 2015, 82, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; He, P.; Munir, S.; He, P.; He, Y.; Li, X.; Yang, L.; Wang, B.; Wu, Y.; He, P. Biocontrol of Soft Rot of Chinese Cabbage Using an Endophytic Bacterial Strain. Front. Microbiol. 2019, 10, 1471. [Google Scholar]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic Pollution in the Environment: From Microbial Ecology to Public Policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef] [PubMed]
- Von der Weid, I.; Alviano, D.S.; Santos, A.L.S.; Soares, R.M.A.; Alviano, C.S.; Seldin, L. Antimicrobial activity of Paenibacillus peoriae strain NRRL BD-62 against a broad spectrum of phytopathogenic bacteria and fungi. J. Appl. Microbiol. 2003, 95, 1143–1151. [Google Scholar] [CrossRef]
- Lorentz, R.H.; Ártico, S.; da Silveira, A.B.; Einsfeld, A.; Corção, G. Evaluation of antimicrobial activity in Paenibacillus spp. strains isolated from natural environment. Lett. Appl. Microbiol. 2006, 43, 541–547. [Google Scholar] [CrossRef]
- He, H.; Ye, Z.; Yang, D.; Yan, J.; Xiao, L.; Zhong, T.; Yuan, M.; Cai, X.; Fang, Z.; Jiang, Y. Characterization of endophytic Rahnella sp. JN6 from Polygonum pubescens and its potential in promoting growth and Cd, Pb, Zn uptake by Brassica napus. Chemosphere 2013, 90, 1960–1965. [Google Scholar] [CrossRef] [PubMed]
- Montefusco, A.; Nakamura, L.K.; Labeda, D.P. Bacillus peoriae sp. nov. Int. J. Syst. Evol. Microbiol. 1993, 43, 388–390. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.E.; Feil, E.J. Multilocus sequence typing—what is resolved? Trends Microbiol. 2005, 12, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaumeil, P.A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2019, 36, 1925–1927. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Chaumeil, P.A.; Rinke, C.; Mussig, A.J.; Hugenholtz, P. A complete domain-to-species taxonomy for Bacteria and Archaea. Nat. Biotechnol. 2020, 38, 1079–1086. [Google Scholar] [CrossRef]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the Ad Hoc Committee on Reconciliation of Approaches to Bacterial Systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Rosselló-Móra, R.; Amann, R. Past and future species definitions for Bacteria and Archaea. Syst. Appl. Microbiol. 2015, 38, 209–216. [Google Scholar] [CrossRef]
- Ciufo, S.; Kannan, S.; Sharma, S.; Badretdin, A.; Clark, K.; Turner, S.; Brover, S.; Schoch, C.L.; Kimchi, A.; DiCuccio, M. Using average nucleotide identity to improve taxonomic assignments in prokaryotic genomes at the NCBI. Int. J. Syst. Evol. Microbiol. 2018, 68, 2386–2392. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.W.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.X.; Zhong, Z.; Zhang, W.P.; Qian, P.Y. Discovery of cationic nonribosomal peptides as Gram-negative antibiotics through global genome mining. Nat. Commun. 2018, 9, 3273. [Google Scholar] [CrossRef] [PubMed]
- Bann, S.J.; Ballantine, R.D.; Cochrane, S.A. The tridecaptins: Non-ribosomal peptides that selectively target Gram-negative bacteria. RSC Med. Chem. 2021, 12, 538–551. [Google Scholar] [CrossRef]
- Choi, S.K.; Park, S.Y.; Kim, R.; Lee, C.H.; Kim, J.F.; Park, S.H. Identification and functional analysis of the fusaricidin biosynthetic gene of Paenibacillus polymyxa E681. Biochem. Biophys. Res. Commun. 2008, 365, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.B.; Yin, C.Y.; Zhou, Y.; Ye, B.C. Prediction of the Mechanism of Action of Fusaricidin on Bacillus subtilis. PLoS ONE 2012, 7, e50003. [Google Scholar] [CrossRef]
- Hollensteiner, J.; Poehlein, A.; Granzow, S.; Liesegang, H.; Daniel, R.; Vidal, S.; Wemheuer, F. Draft Genome Sequence of the Endophyte Bacillus mycoides Strain GM5LP Isolated from Lolium perenne. Genome Announc. 2018, 6, e01517-17. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.; Choi, S.K.; Ryu, C.M.; Park, S.H. Chronicle of a Soil Bacterium: Paenibacillus polymyxa E681 as a Tiny Guardian of Plant and Human Health. Front. Microbiol. 2019, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef] [PubMed]
- Elnahal, A.S.M.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.S.M.; El-Tahan, A.M.; Rady, M.M.; AbuQamar, S.F.; El-Tarabily, K.A. The use of microbial inoculants for biological control, plant growth promotion, and sustainable agriculture: A review. Eur. J. Plant Pathol. 2022, 162, 759–792. [Google Scholar] [CrossRef]
- Attallah, M.F.; Metwally, S.S.; Moussa, S.I.; Soliman, M.A. Environmental impact assessment of phosphate fertilizers and phosphogypsum waste: Elemental and radiological effects. Microchem. J. 2019, 146, 789–797. [Google Scholar] [CrossRef]
- Lee, E.K.; Zhang, X.; Adler, P.R.; Kleppel, G.S.; Romeiko, X.X. Spatially and temporally explicit life cycle global warming, eutrophication, and acidification impacts from corn production in the U.S. Midwest. J. Clean. Prod. 2020, 242, 118465. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Fact. 2016, 15, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, L.F.; López, M.G.; Straube, L.; Passaglia, L.M.P.; Wendisch, V.F. Inorganic Phosphate Solubilization by Rhizosphere Bacterium Paenibacillus sonchi: Gene Expression and Physiological Functions. Front. Microbiol. 2020, 11, 588605. [Google Scholar] [CrossRef] [PubMed]
- Weselowski, B.; Nathoo, N.; Eastman, A.W.; MacDonald, J.; Yuan, Z.C. Isolation, identification and characterization of Paenibacillus polymyxa CR1 with potentials for biopesticide, biofertilization, biomass degradation and biofuel production. BMC Microbiol. 2016, 16, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gao, T.; Wang, Q. Comparative and Functional Analyses of Two Sequenced Paenibacillus polymyxa Genomes Provides Insights Into Their Potential Genes Related to Plant Growth-Promoting Features and Biocontrol Mechanisms. Front. Genet. 2020, 11, 564939. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Grantcharova, N.; Wagner, E.G.H. Paenibacillus polymyxa Invades Plant Roots and Forms Biofilms. Appl. Environ. Microbiol. 2005, 71, 7292–7300. [Google Scholar] [CrossRef] [Green Version]
- Poudel, S.; Giannone, R.J.; Farmer, A.T.; Campagna, S.R.; Bible, A.N.; Morrell-Falvey, J.L.; Elkins, J.G.; Hettich, R.L. Integrated Proteomics and Lipidomics Reveal That the Swarming Motility of Paenibacillus polymyxa Is Characterized by Phospholipid Modification, Surfactant Deployment, and Flagellar Specialization Relative to Swimming Motility. Front. Microbiol. 2019, 10, 2594. [Google Scholar] [CrossRef] [PubMed]
- Terashima, H.; Kojima, S.; Homma, M. Chapter 2 Flagellar Motility in Bacteria: Structure and Function of Flagellar Motor. Int. Rev. Cell Mol. Biol. 2008, 270, 39–85. [Google Scholar]
- Guttenplan, S.B.; Kearns, D.B. Regulation of flagellar motility during biofilm formation. FEMS Microbiol. Rev. 2013, 37, 849–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julkowska, D.; Obuchowski, M.; Holland, I.B.; Séror, S.J. Comparative Analysis of the Development of Swarming Communities of Bacillus subtilis 168 and a Natural Wild Type: Critical Effects of Surfactin and the Composition of the Medium. J. Bacteriol. 2005, 187, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Bree, A.C.; Liu, J.; Patrick, J.E.; Chien, P.; Kearns, D.B. Adaptor-mediated Lon proteolysis restricts Bacillus subtilis hyperflagellation. Proc. Natl. Acad. Sci. USA 2015, 112, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Kanesaki, Y.; Yoshikawa, H. Genetic Analysis of Collective Motility of Paenibacillus sp. NAIST15-1. PLoS Genet. 2016, 12, e1006387. [Google Scholar] [CrossRef] [PubMed]
- Colin, R.; Ni, B.; Laganenka, L.; Sourjik, V. Multiple functions of flagellar motility and chemotaxis in bacterial physiology. FEMS Microbiol. Rev. 2021, 45, fuab038. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, Y.H.; Zhu, H.Z.; Andrianova, E.P.; Jiang, C.Y.; Li, D.; Ma, L.; Feng, J.; Liu, Z.P.; Xiang, H.; et al. Cross Talk between Chemosensory Pathways That Modulate Chemotaxis and Biofilm Formation. mBio 2019, 10, e02876-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadhams, G.H.; Armitage, J.P. Making sense of it all: Bacterial chemotaxis. Nat. Rev. Mol. Cell Biol. 2004, 5, 1024–1037. [Google Scholar] [CrossRef]
- Liu, W.; Sun, Y.; Shen, R.; Dang, X.; Liu, X.; Sui, F.; Li, Y.; Zhang, Z.; Alexandre, G.; Elmerich, C.; et al. A Chemotaxis-Like Pathway of Azorhizobium caulinodans Controls Flagella-Driven Motility, Which Regulates Biofilm Formation, Exopolysaccharide Biosynthesis, and Competitive Nodulation. Mol. Plant Microbe Interact. 2018, 31, 737–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharf, B.E.; Hynes, M.F.; Alexandre, G.M. Chemotaxis signaling systems in model beneficial plant–bacteria associations. Plant Mol. Biol. 2016, 90, 549–559. [Google Scholar] [CrossRef]
- Mangwani, N.; Kumari, S.; Das, S. Taxonomy and Characterization of Biofilm Forming Polycyclic Aromatic Hydrocarbon Degrading Bacteria from Marine Environments. Polycycl. Aromat. Compd. 2019, 41, 1249–1262. [Google Scholar] [CrossRef]
- Branda, S.S.; Vik, Å.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Zhang, D.; Cheng, Y.; Tan, J.; Luo, Y. The impact of Paenibacillus polymyxa HY96-2 luxS on biofilm formation and control of tomato bacterial wilt. Appl. Microbiol. Biotechnol. 2019, 103, 9643–9657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voichek, M.; Maaß, S.; Kroniger, T.; Becher, D.; Sorek, R. Peptide-based quorum sensing systems in Paenibacillus polymyxa. Life Sci. Alliance. 2020, 3, e202000847. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Farag, M.A.; Park, H.B.; Kloepper, J.W.; Lee, S.H.; Ryu, C. Induced Resistance by a Long-Chain Bacterial Volatile: Elicitation of Plant Systemic Defense by a C13 Volatile Produced by Paenibacillus polymyxa. PLoS ONE 2012, 7, e48744. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Disease Index | Control Effect (%) |
|---|---|---|
| ZF115 | 76.00 ± 6.93 bcd | 19.72 ± 7.32 defg |
| ZF119 | 77.33 ± 4.62 abc | 18.31 ± 4.88 efg |
| ZF185 | 88.00 ± 6.93 ab | 7.05 ± 7.32 g |
| ZF194 | 34.67 ± 4.62 fg | 63.38 ± 4.88 ab |
| ZF278 | 81.33 ± 12.86 ab | 14.09 ± 13.58 g |
| ZF390 | 21.33 ± 11.55 g | 77.47 ± 12.20 a |
| ZF402 | 58.67 ± 14.05 e | 38.03 ± 14.84 cd |
| ZF405 | 78.67 ± 6.11 abc | 16.90 ± 6.45 fg |
| ZF428 | 40.00 ± 8.00 f | 57.75 ± 8.45 b |
| ZF429 | 85.33 ± 6.11 ab | 9.86 ± 6.45 g |
| ZF436 | 60.00 ± 12.00 de | 36.62 ± 12.68 cde |
| ZF448 | 78.67 ± 4.62 abc | 16.90 ± 4.88 fg |
| ZF450 | 76.00 ± 12.00 bcd | 19.72 ± 12.68 defg |
| ZF453 | 62.67 ± 16.17 cde | 33.81 ± 17.08 cdef |
| Zhongshengmycin | 49.33 ± 2.31 ef | 47.89 ± 2.44 bc |
| Control | 94.67 ± 6.11 a | - |
| Antibiotic Name | Type | Core Gene Clusters | Size | Position | Bioactive Spectrum | ZF390 | HS311 | SQR-21 | HY96-2 | PS04 |
|---|---|---|---|---|---|---|---|---|---|---|
| Brevicidine | T1PKS, NRPS | fenA, ftdB | 761,14 bp | 2792116–2868229 | G-bacteria | IAQ67_12465- IAQ67_12650 | ABE82_12870-ABE82_13050 | PPSQR21_024840-PPSQR21_025180 | C1A50_2551-C1A50_2581 | NA |
| Fusaricidin B | NRPS | fadD, bacC | 68,481 bp | 62571– 131051 | Fungi, G+ bacteria | IAQ67_00290- IAQ67_00510 | ABE82_00305-ABE82_00500 | PPSQR21_000590-PPSQR21_001000 | C1A50_0064-C1A50_0108 | FOA15_RS06155- FOA15_RS06360 |
| Octapeptin | NRPS | fenAB, tycC | 115,828 bp | 5324980–5440807 | Fungi, bacteria | IAQ67_24570- IAQ67_24795 | ABE82_22040-ABE82_22245 | PPSQR21_044420-PPSQR21_044780 | C1A50_4551-C1A50_4603 | NA |
| Paenilan | Lanthipeptide- class-i | epiBC | 27,007 bp | 1620879–1647885 | G+ bacteria | IAQ67_07435- IAQ67_07530 | NA | PPSQR21_015740-PPSQR21_015950 | C1A50_1599-C1A50_1620 | NA |
| Paenilipoheptin | TransAT-PKS, NRPS | fabD, ituA, fenC, fusAA | 82,343 bp | 2634231–2716573 | NA | IAQ67_11935- IAQ67_12110 | ABE82_12325-ABE82_12500 | PPSQR21_009520-PPSQR21_009940 | C1A50_0993-C1A50_1036 | FOA15_RS17665- FOA15_RS17835 |
| Paeninodin | Lassopeptide | asnB, pqqD | 24,119 bp | 1301862–1325980 | Fungi, bacteria, virus | IAQ67_05860- IAQ67_05975 | ABE82_06065-ABE82_06175 | PPSQR21_012140-PPSQR21_012370 | C1A50_1276-C1A50_1299 | FOA15_RS11780- FOA15_RS11890 |
| Tridecaptin | NRPS | fusAA, bacC | 92,515 bp | 2419321–2511835 | G-bacteria | IAQ67_11205- IAQ67_11360 | ABE82_11520-ABE82_11675 | PPSQR21_022460-PPSQR21_022870 | C1A50_2315-C1A50_2359 | FOA15_RS17145- FOA15_RS17325 |
| Unknown | Siderophore | sbnCEF | 17,402 bp | 999368– 1016769 | NA | IAQ67_04460- IAQ67_04525 | ABE82_04615-ABE82_04685 | NA | NA | FOA15_RS16945- FOA15_RS17000 |
| Unknown | RiPP-like | NA | 10,237 bp | 1167935–1178171 | NA | IAQ67_05235- IAQ67_05290 | ABE82_05390-ABE82_05445 | PPSQR21_010820-PPSQR21_010940 | C1A50_1133-C1A50_1145 | FOA15_RS11190- FOA15_RS11225 |
| Unknown | NRPS-like | NA | 43,119 bp | 1991107–2034225 | NA | IAQ67_09200- IAQ67_09365 | ABE82_09545-ABE82_09710 | PPSQR21_018700-PPSQR21_019060 | C1A50_1915-C1A50_1953 | FOA15_RS15165- FOA15_RS15330 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Xie, X.; Li, J.; Shi, Y.; Chai, A.; Fan, T.; Li, B.; Li, L. Comparative Genomics Insights into a Novel Biocontrol Agent Paenibacillus peoriae Strain ZF390 against Bacterial Soft Rot. Biology 2022, 11, 1172. https://doi.org/10.3390/biology11081172
Zhao Y, Xie X, Li J, Shi Y, Chai A, Fan T, Li B, Li L. Comparative Genomics Insights into a Novel Biocontrol Agent Paenibacillus peoriae Strain ZF390 against Bacterial Soft Rot. Biology. 2022; 11(8):1172. https://doi.org/10.3390/biology11081172
Chicago/Turabian StyleZhao, Yurong, Xuewen Xie, Junhui Li, Yanxia Shi, Ali Chai, Tengfei Fan, Baoju Li, and Lei Li. 2022. "Comparative Genomics Insights into a Novel Biocontrol Agent Paenibacillus peoriae Strain ZF390 against Bacterial Soft Rot" Biology 11, no. 8: 1172. https://doi.org/10.3390/biology11081172
APA StyleZhao, Y., Xie, X., Li, J., Shi, Y., Chai, A., Fan, T., Li, B., & Li, L. (2022). Comparative Genomics Insights into a Novel Biocontrol Agent Paenibacillus peoriae Strain ZF390 against Bacterial Soft Rot. Biology, 11(8), 1172. https://doi.org/10.3390/biology11081172