


Distribution and Prevalence of Plant-Parasitic Nematodes of Turfgrass at Golf Courses in China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
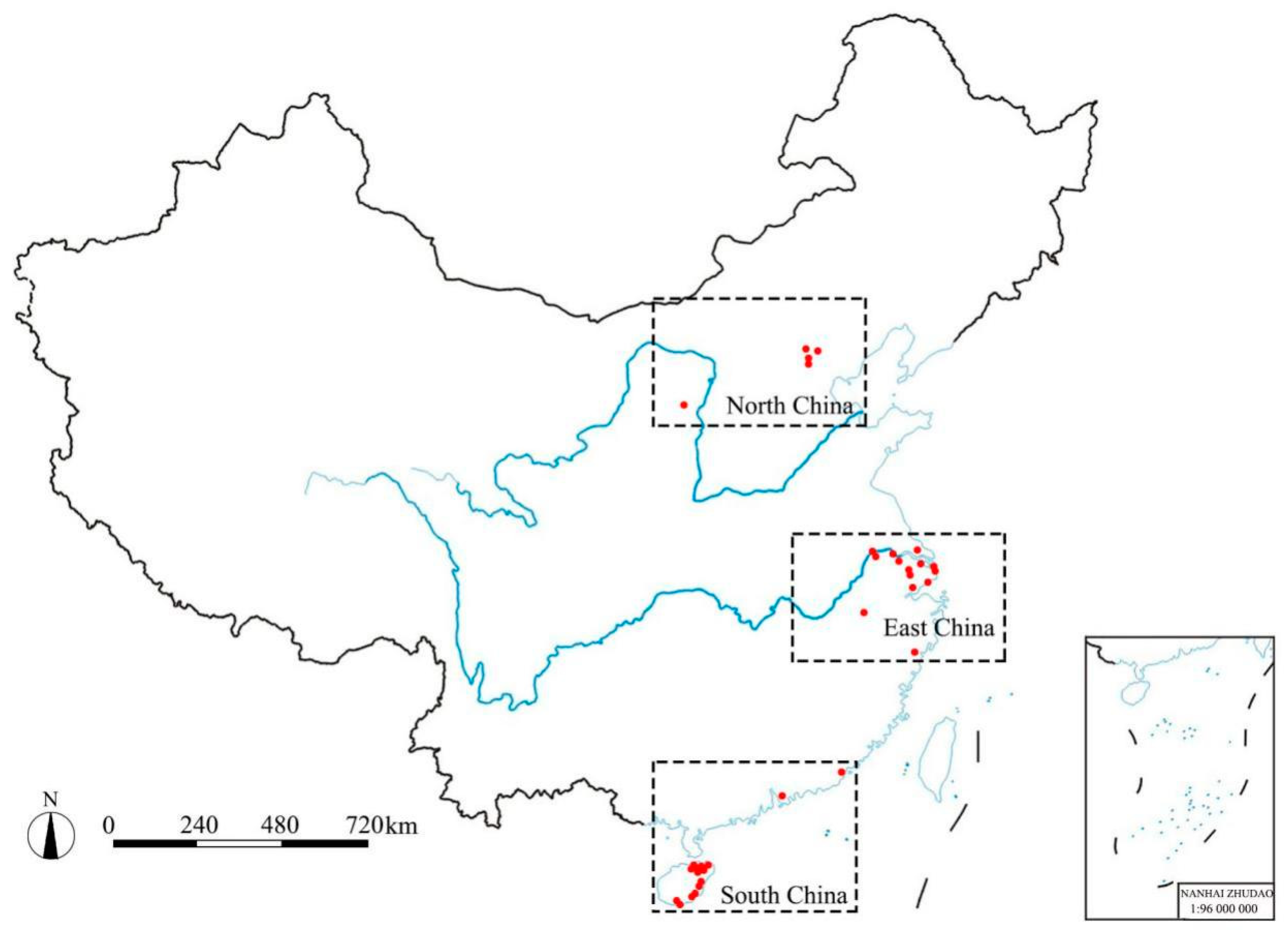
2.1. Sample Collection
2.2. Nematode Extraction and Identification
2.3. Data Analysis
3. Results
3.1. Classification and Distribution of Plant-Parasitic Nematodes
3.2. Prevalence of the Main PPNs
3.3. PPNs and Soil Health
3.4. Nematode Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uddin, M.K.; Juraimi, A.S. Salinity Tolerance Turfgrass: History and Prospects. Sci. World J. 2013, 2013, 409413. [Google Scholar] [CrossRef]
- Stowell, L.J.; Gelernter, W.D. Diagnosis of turfgrass diseases. Annu. Rev. Phytopathol. 2001, 39, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Smiley, R.W.; Dernoeden, P.H.; Clarke, B.B. Compendium of Turfgrass Diseases, 2nd ed.; American Phytopathological Society: St. Paul, MN, USA, 1992. [Google Scholar]
- Stackhouse, T.; Martinez-Espinoza, A.D.; Ali, M.E. Turfgrass disease diagnosis: Past, present, and future. Plants 2020, 9, 1544. [Google Scholar] [CrossRef] [PubMed]
- Jordan, K.; Mitkowski, N. Population dynamics of plant-parasitic nematodes in golf course greens turf in southern New England. Plant Dis. 2006, 90, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Stirling, G.; Stirling, A.; Eden, L. Plant-parasitic nematodes on turfgrass in Queensland, Australia, and biosecurity issues associated with the interstate transfer and eradication of southern sting nematode (Ibipora lolii). Australas. Plant Pathol. 2021, 50, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.S.; Ye, W.; Martin, S.B.; Tredway, L. Diversity and occurrence of plant-parasitic nematodes associated with golf course turfgrasses in North and South Carolina, USA. Nematology 2012, 44, 337–347. [Google Scholar]
- Chen, X.G. Identification of Parasitic Nematodes on Golf Course Turfgrasses in Guangdong. Ph.D. Thesis, Zhongkai University of Agriculture and Engineering, Guangzhou, China, 2017. [Google Scholar]
- Zeng, Y.S.; Ye, W.; Tredway, L.; Martin, S.; Martin, M. Taxonomy and morphology of plant-parasitic nematodes associated with turfgrasses in North and South Carolina, USA. Zootaxa 2012, 3452, 1–46. [Google Scholar] [CrossRef]
- Vandenbossche, B.; Viaene, N.; Sutter, N.; Maes, M.; Karssen, G.; Bert, W. Diversity and incidence of plant-parasitic nematodes in Belgian turfgrass. Nematology 2011, 13, 245–256. [Google Scholar] [CrossRef]
- Mwamula, A.O.; Lee, D.W. Occurrence of plant-parasitic nematodes of turfgrass in Korea. Plant Pathol. J. 2021, 37, 446–454. [Google Scholar] [CrossRef]
- Mcleod, R.W.; Reay, F.; Smyth, J. Plant Nematodes of Australia Listed by Plant and by Genes; NSW Agriculture: Orange, Australia, 1995.
- LaMondia, J.; Wick, R.L.; Mitkowski, N. Plant parasitic nematodes of New England: Connecticut, Massachusetts and Rhode Island. Sustainability in plant and crop protection. In Plant Parasitic Nematodes in Sustainable Agriculture of North America; Subbotin, S., Chitambar, J., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Crow, W.T.; Habteweld, A.W.; Bean, T. Mist chamber extraction for improved diagnosis of Meloidogyne spp. from golf course bermudagrass. J. Nematol. 2020, 52, 1–12. [Google Scholar] [CrossRef]
- Lucas, L.T. Population dynamics of Belonolaimus longicaudatus and Criconemella ornata and growth response of bermudagrass and overseeded grasses on golf greens following treatment with nematicides. J. Nematol. 1982, 14, 358–363. [Google Scholar] [PubMed]
- Buckley, R.J.; Koppenhöfer, A.M.; Crow, W.T. Integrated approach to nematode management in turfgrass. Plant Pest Advis. 2008, 14, 4–7. [Google Scholar]
- Crow, W.T. Nematode management for golf courses in Florida. EDIS 2018. [Google Scholar] [CrossRef]
- Ruscoe, P.E.; Stirling, G.R. Southern sting nematode (Ibipora lolii), a serious pest of turf grasses in Australia. A review of what can be learnt from research on Belonolaimus longicaudatus, a closely related pest of turfgrass and many crops in the United States. Australas. Plant Pathol. 2020, 49, 493–504. [Google Scholar] [CrossRef]
- Barker, K.R.; Nusbaum, C.J.; Nelson, L.A. Effects of storage temperature and extraction procedure on recovery of plant-parasitic nematodes from field soils. J. Nematol. 1969, 1, 240–247. [Google Scholar] [PubMed]
- Whitehead, A.G.; Hemming, J. A comparison of some quantitative methods of extracting small vermiform nematodes from soil. Ann. Appl. Biol. 1965, 55, 25–38. [Google Scholar] [CrossRef]
- Mao, X.F.; Li, H.X.; Chen, X.Y. Extraction efficiency of soil nematodes by different methods. Chin. J. Ecol. 2004, 23, 149–151. [Google Scholar]
- Chitwood, B.G. Soil and freshwater nematodes. Science 1963, 144, 721. [Google Scholar] [CrossRef]
- Perry, R.N.; Moens, M. Plant Nematology; CABI Publishing: Wallingford, UK, 2006. [Google Scholar]
- Xie, H. Taxonomy of Plant Nematodes, 2nd ed.; Higher Education Press: Beijing, China, 2005. [Google Scholar]
- Antoinette, S.; Mariette, M.; Denberg, E. Nematode Morphology and Classification; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Rong, W.T.; Wang, J.C.; Liu, P.; Lin, Y.; Guo, J.Z.; Huang, G.M.; Sun, J.H. Research on DNA extraction methods from a single nematode. Acta Agric. Boreali-Sin. 2014, 29, 127–132. [Google Scholar]
- Nunn, G.B. Nematode molecular evolution: An Investigation of Evolutionary Patterns Among Nematodes Based upon DNA Sequences. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1992. [Google Scholar]
- Mullin, P.; Harris, T.; Powers, T. Phylogenetic relationships of Nygolaimina and Dorylaimina (Nematoda: Dorylaimida) inferred from small subunit ribosomal DNA sequences. J. Nematol. 2005, 7, 59–79. [Google Scholar] [CrossRef]
- Sawadogo, A.; Thio, B.; Kiemde, S.; Drabo, I.; Dabire, C.L.; Ouedraogo, J.T.; Mullens, T.R.; Ehlers, J.D.; Roberts, P.A. Distribution and prevalence of parasitic nematodes of cowpea (Vigna unguiculata) in Burkina Faso. J. Nematol. 2009, 41, 120–127. [Google Scholar] [PubMed]
- Nelson, E. Nematode disorders of turfgrasses: How important are they? Turf Grass Trends 1995, 4, 1–16. [Google Scholar]
- Yeates, G.W.; Bird, A.F. Some observations on the influence of agricultural practices on the nematode faunae of some South Australian soils. Fundam. Appl. Nematol. 1994, 17, 133–145. [Google Scholar]
- Ekschmitt, K.; Bakonyi, G.; Bongers, M.; Bongers, T.; Boström, S.; Dogan, H.; Harrison, A.; Nagy, P.; O’Donnell, A.G.; Papatheodorou, E.M.; et al. Nematode community structure as indicator of soil functioning in European grassland soils. Eur. J. Soil Biol. 2001, 37, 263–268. [Google Scholar] [CrossRef]
- Gautier, M. Nematodes in fruit tree nurseries. Arboric. Fruit. 1975, 252, 17–25. [Google Scholar]
- Fang, Y.S.; Zhuo, K.; Gao, J.S.; Wang, X.Y. Preliminary survey and identification of grass carpey nematode in Guangzhou region. Pratacultural Sci. 2003, 20, 37–42. [Google Scholar]
- Wei, Y.J. Investigation and identification of species of parasitic nematodes in grass in Huaian region. J. Anhui Agric. Sci. 2006, 34, 3413. [Google Scholar]
- Bond, J.P.; Mcgawley, E.C.; Hoy, J.W. Distribution of plant-parasitic nematodes on sugarcane in Louisiana and efficacy of nematicides. J. Nematol. 2000, 32, 493–501. [Google Scholar]
- Simard, L.; Bélair, G.; Powers, T.O.; Tremblay, N.; Dionne, J. Incidence and population density of plant-parasitic nematodes on golf courses in Ontario and Québec, Canada. J. Nematol. 2008, 40, 241–251. [Google Scholar]
- Liang, L.L.; Liu, Q.Z.; Xie, N.; Li, X.Y.; Yang, D.; Bian, Y.; Xie, D.Y. Nematode trophic group structure in wilted turf soil and its regulation factors. Acta Agric. Zhejiangensis 2011, 23, 948–954. [Google Scholar]
- Liu, Q.Z.; Liang, L.L.; Yang, D.; Bian, Y.; Zhou, H.Y.; Xie, D.Y. Analysis of community characteristics of nematodes in turf soil of Yongfeng virescence region in Beijing. Acta Pratacultural Sci. 2009, 18, 136–141. [Google Scholar]
- Lucas, L.T.; Blake, C.T.; Barker, K.R. Nematodes associated with bentgrass and bermudagrass golf greens in North Carolina. Plant Dis. Report. 1974, 58, 822–824. [Google Scholar]
- Sikora, E.J.; Guertal, E.A.; Bowen, K.L. Plant-parasitic nematodes associated with hybrid bermudagrass and creeping bentgrass putting greens in Alabama. Nematropica 2001, 31, 301–306. [Google Scholar]
- Walker, N.R.; Goad, C.L.; Zhang, H.; Martin, D.L. Factors associated with populations of plant-parasitic nematodes in bentgrass putting greens in Oklahoma. Plant Dis. 2002, 86, 764–768. [Google Scholar] [CrossRef] [PubMed]
- Janati, S.; Houari, A.; Wifaya, A.; Essarioui, A.; Mimouni, A.; Hormatallah, A.; Sbaghi, M.; Dababat, A.; Mokrini, F. Occurrence of the root-knot nematode species in vegetable crops in Souss region of Morocco. Plant Pathol. J. 2018, 34, 308–315. [Google Scholar] [CrossRef]
- Ye, W.; Zeng, Y.; Kerns, J.P. Molecular characterisation and diagnosis of root-knot nematodes (Meloidogyne spp.) from turfgrasses in North Carolina, USA. PLoS ONE 2015, 10, e0143556. [Google Scholar] [CrossRef]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161–168. [Google Scholar] [PubMed]
- Popovici, I.; Ciobanu, M. Diversity and distribution of nematode communities in grasslands from Romania in relation to vegetation and soil characteristics. Appl. Soil Ecol. 2000, 14, 27–36. [Google Scholar] [CrossRef]
- Zhang, W.; Nan, Z.B.; Tian, P.; Hu, M.J.; Gao, Z.Y.; Li, M.; Liu, G.D. Microdochium paspali, a new species causing seashore paspalum disease in southern China. Mycologia 2015, 107, 80–89. [Google Scholar] [CrossRef]
- Allan-Perkins, E.; Manter, D.K.; Wick, R.L.; Ebdon, S.; Jung, G. Nematode communities on putting greens, fairways, and roughs of organic and conventional cool-season golf courses. Appl. Soil Ecol. 2017, 121, 161–171. [Google Scholar] [CrossRef]
- Kostenko, O.; Duyts, H.; Grootemaat, S.; De Deyn, G.B.; Bezemer, T.M. Plant diversity and identity effects on predatory nematodes and their prey. Ecol. Evol. 2015, 5, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Neher, D.A. Ecology of plant and free-living nematodes in natural and agricultural soil. Annu. Rev. Phytopathol. 2010, 48, 371–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Region | Sampling Date | Golf Course | Turfgrass Species | No. of Samples |
|---|---|---|---|---|
| NC | July 2021 | Beijing Qinghewan | Creeping bentgrass | 2 |
| July 2021 | Beijing Laffitte | Creeping bentgrass | 2 | |
| August 2021 | Beijing Yuyang | Creeping bentgrass | 4 | |
| October 2021 | Beijing CBD | Creeping bentgrass | 2 | |
| October 2021 | Shaanxi Yulin Desert | Kentucky bluegrass | 2 | |
| EC | May 2021 | Nantong Xiao Yang Kou | Creeping bentgrass, Seashore paspalum | 2 |
| July 2021 | Nanjing Sun Island | Creeping bentgrass | 6 | |
| August 2021 | Nanjing Zhongshan | Creeping bentgrass | 2 | |
| July 2021 | Suzhou Taihu | Creeping bentgrass | 8 | |
| July 2021 | Taicang East Sea | Creeping bentgrass | 2 | |
| July 2021 | Suzhou Jinji Lake | Creeping bentgrass | 6 | |
| September 2021 | Changzhou Kingswan | Creeping bentgrass | 5 | |
| September 2021 | Suzhou Shuangshan | Creeping bentgrass | 4 | |
| May 2021 | Huzhou Hotspring | Creeping bentgrass | 3 | |
| July 2021 | Haining Lake Hill | Creeping bentgrass | 2 | |
| July 2021 | Shanghai Sino-Bay | Creeping bentgrass | 4 | |
| August 2021 | Shanghai Links | Creeping bentgrass | 2 | |
| August 2021 | Shanghai Tomson | Creeping bentgrass | 2 | |
| September 2021 | Huangshan Pine | Hybrid bermudagrass | 4 | |
| SC | November 2020 | Hainan Meilan | Hybrid bermudagrass | 4 |
| November 2020 | Hainan Meilangwan | Seashore paspalum | 5 | |
| November 2020 | Hainan West Coast | Seashore paspalum | 6 | |
| November 2020 | Hainan Meishi | Hybrid bermudagrass | 4 | |
| November 2020 | Hainan Mission Hills | Hybrid bermudagrass | 4 | |
| November 2020 | Haikou Three Miles | Hybrid bermudagrass | 7 | |
| November 2020 | Boao Asia BBS | Hybrid bermudagrass, Seashore paspalum | 4 | |
| November 2020 | Qionghai White Stone Hot Spring | Seashore paspalum | 2 | |
| July 2021 | Shenzhou Peninsula | Seashore paspalum | 4 | |
| November 2020 | Sanya Blue Bay | Seashore paspalum | 6 | |
| July 2021 | Wanning Shanqin Bay | Seashore paspalum | 4 | |
| September 2021 | Sanya Luhuitou | Hybrid bermudagrass | 3 | |
| September 2021 | Guangdong Chaoshan | Creeping bentgrass | 2 | |
| October 2021 | Shenzhen Sand River | Hybrid bermudagrass, Seashore paspalum | 8 |
| Order | Nematode Classification | Region | Golf Course | Turfgrass Species | No. of Courses |
|---|---|---|---|---|---|
| Order Tylenchida, family Anguinidae | - | ||||
| 1 | Ditylenchus spp. | EC | Nanjing Zhongshan | Creeping bentgrass | 1 |
| 2 | Ditylenchus destructor | EC | Suzhou Taihu | Creeping bentgrass | 1 |
| 3 | Ditylenchus dipsaci | EC | Huzhou Hotspring, Shanghai Sino-Bay, Shanghai Links | Creeping bentgrass | 3 |
| SC | Wanning Shanqin Bay, Hainan Mission Hills | Hybrid bermudagrass, Seashore Paspalum | 2 | ||
| 4 | Ditylenchus cf. microdens | NC | Shaanxi Yulin Desert | Kentucky bluegrass | 1 |
| Order Tylenchida, family Belonolaimidae | - | ||||
| 5 | Tylenchorhynchus annulatus | NC | Beijing Laffitte | Creeping bentgrass | 1 |
| Order Tylenchida, family Criconematidae | - | ||||
| 6 | Criconemoides annulatus | NC | Beijing Laffitte | Creeping bentgrass | 1 |
| 7 | Mesocriconema spp. | NC | Shaanxi Yulin Desert | Kentucky bluegrass | 1 |
| SC | Haikou Three Miles, Guangdong Chaoshan, Shenzhen Sand River | Creeping bentgrass, Seashore Paspalum, Hybrid bermudagrass | 3 | ||
| 8 | Mesocriconema curvatum | NC | Beijing Laffitte, Beijing Yuyang | Creeping bentgrass | 2 |
| 9 | Hemicycliophoraconida | NC | Shaanxi Yulin Desert | Kentucky bluegrass | 1 |
| 10 | Hemicriconemoides spp. | EC | Nantong Xiao Yang Kou | Creeping bentgrass | 1 |
| SC | Hainan West Coast, Wanning Shanqin Bay, Shenzhou Peninsula, Sanya Luhuitou, Haikou Meilangwan, Hainan Mission Hills | Seashore Paspalum, Hybrid bermudagrass | 6 | ||
| Order Tylenchida, family Heteroderidae | - | ||||
| 11 | Heterodera avenae | NC | Beijing CBD | Creeping bentgrass | 1 |
| 12 | Meloidogyne graminis | EC | Changzhou Kingswan, Nanjing Zhongshan | Creeping bentgrass | 2 |
| SC | Shenzhen Sand River | Hybrid bermudagrass, Seashore Paspalum | 1 | ||
| 13 | Meloidogyne incognita | EC | Suzhou Jinji Lake | Creeping bentgrass | 1 |
| Order Tylenchida, family Hoplolaimidae | - | ||||
| 14 | Helicotylenchus spp. | EC | Suzhou Jinji Lake, Suzhou Shuangshan, Shanghai Sino-Bay, Huangshan Pine | Creeping bentgrass, Hybrid bermudagrass | 4 |
| SC | Haikou Meishi, Haikou Meilangwan, Boao Asia BBS, Qionghai White Stone Hot Spring | Hybrid bermudagrass, Seashore Paspalum | 4 | ||
| 15 | Helicotylenchus microlobus | NC | Beijing Qinghewan, Beijing Laffitte | Creeping bentgrass | 2 |
| SC | Hainan Mission Hills, Haikou Three Miles, Haikou Meilan, Wanning Shanqin Bay | Hybrid bermudagrass, Seashore Paspalum | 4 | ||
| 16 | Helicotylenchus digitiformis | EC | Nantong Xiao Yang Kou | Creeping bentgrass | 1 |
| 17 | Helicotylenchus dihystera | NC | Beijing Yuyang | Creeping bentgrass | 1 |
| SC | Shenzhou Peninsula, Hainan West Coast, Guangdong Chaoshan | Hybrid bermudagrass, Seashore Paspalum | 3 | ||
| 18 | Helicotylenchus paraplatyurus | SC | Hainan Mission Hills | Hybrid bermudagrass | 1 |
| 19 | Helicotylenchus pseudorobustus | NC | Beijing CBD | Creeping bentgrass | 1 |
| SC | Hainan West Coast | Seashore Paspalum | 1 | ||
| 20 | Helicotylenchus rotundicauda | EC | Nantong Xiao Yang Kou | Creeping bentgrass | 1 |
| 21 | Helicotylenchus indicus | SC | Shenzhen Sand River | Hybrid bermudagrass, Seashore Paspalum | 1 |
| 22 | Hoplolaimus columbus | EC | Suzhou Jinji Lake, Nanjing Zhongshan, Shanghai Sino-Bay, Links, Shanghai Tomson | Creeping bentgrass | 5 |
| SC | Hainan West Coast, Guangdong Chaoshan | Seashore Paspalum, Creeping bentgrass | 2 | ||
| 23 | Rotylenchulus spp. | SC | Haikou Three Miles | Hybrid bermudagrass | 1 |
| Order Tylenchida, family Pratylenchidae | - | ||||
| 24 | Hirschmanniellaoryzae | EC | Huzhou Hotspring | Creeping bentgrass | 1 |
| Order Tylenchida, family Tylenchidae | - | ||||
| 25 | Filenchus spp. | NC | Beijing Qinghewan, Beijing Yuyang | Creeping bentgrass | 2 |
| EC | Suzhou Jinji Lake, Suzhou Shuangshan | Creeping bentgrass | 2 | ||
| SC | Hainan Meilan, Haikou Meilangwan, Haikou Meishi, Hainan West Coast, Haikou Three Miles, Qionghai White Stone Hot Spring | Hybrid bermudagrass, Seashore Paspalum | 6 | ||
| 26 | Filenchus discrepans | EC | Suzhou Taihu, Taicang East Sea | Creeping bentgrass | 2 |
| 27 | Filenchus misellus | EC | Huangshan Pine | Hybrid bermudagrass | 1 |
| 28 | Discotylenchus spp. | NC | Beijing CBD | Creeping bentgrass | 1 |
| EC | Shanghai Tomson | Creeping bentgrass | 1 | ||
| 29 | Ecphyadophora quadralata | EC | Suzhou Taihu | Creeping bentgrass | 1 |
| 30 | Labryskhuzestanensis | SC | Shenzhou Peninsula | Seashore Paspalum | 1 |
| 31 | Labrys chinensis | SC | Wanning Shanqin Bay | Seashore Paspalum | 1 |
| Order Triplonchida, family Trichodoridae | |||||
| 32 | Paratrichodorus porosus | NC | Beijing Qinghewan | Creeping bentgrass | 1 |
| 33 | Paratrichodorus minor | EC | Suzhou Taihu, Suzhou Jinji Lake, Suzhou Shuangshan, Changzhou Kingswan, Nanjing Sun Island, Nantong Xiao Yang Kou, Shanghai Sino-Bay, Huangshan Pine | Creeping bentgrass, Seashore Paspalum, Hybrid bermudagrass | 8 |
| Order Dorylaimida, family Longidoridae | - | ||||
| 34 | Xiphinema spp. | EC | Huangshan Pine | Hybrid bermudagrass | 1 |
| SC | Boao Asia BBS | Hybrid bermudagrass, Seashore Paspalum | 1 | ||
| 35 | Xiphinema nuragicum | EC | Suzhou Jinji Lake, Nanjing Sun Island, Nanjing Zhongshan, Shanghai Tomson | Creeping bentgrass | 4 |
| 36 | Xiphinema hunaniense | EC | Nanjing Sun Island | Creeping bentgrass | 1 |
| 37 | Xiphinema insigne | EC | Shanghai Sino-Bay | Creeping bentgrass | 1 |
| 38 | Xiphinema ifacolum | EC | Shanghai Sino-Bay, Nanjing Zhongshan | Creeping bentgrass | 2 |
| SC | Shenzhen Sand River | Hybrid bermudagrass, Seashore Paspalum | 1 | ||
| 39 | Paralongidorus koreanensis | SC | Shenzhen Sand River | Hybrid bermudagrass | 1 |
| Order Aphelenchida, family Aphelenchoididae | - | ||||
| 40 | Aphelenchoides pseudogoodeyi | SC | Hainan Mission Hills, Boao Asia BBS | Hybrid bermudagrass, Seashore Paspalum | 2 |
| 41 | Aphelenchoides bicaudatus | EC | Shanghai Sino-Bay, Shanghai Tomson | Creeping bentgrass | 2 |
| SC | Guangdong Chaoshan | Creeping bentgrass | 1 |
| Region 1 | Genus 2 | Infestation (%) 3 | Frequency (%) | Abundance | Max (Individuals/100 cm3 of Soil) | Damage Threshold 4 | Samples above Threshold (%) 5 |
|---|---|---|---|---|---|---|---|
| NC | Helicotylenchus | 80.00 | 75.00 | 1.89 | 215 | 600 | - |
| Mesocriconema | 60.00 | 41.67 | 1.16 | 48 | - | - | |
| EC | Filenchus | 35.71 | 21.15 | 1.65 | 392 | - | - |
| Paratrichodorus | 64.29 | 59.62 | 1.36 | 103 | 80 | 9.68 | |
| Xiphinema | 42.86 | 23.08 | 1.86 | 328 | 200 | 8.33 | |
| Hoplolaimus | 35.71 | 23.08 | 1.47 | 135 | 150 | - | |
| Ditylenchus | 35.71 | 15.38 | 1.88 | 315 | - | - | |
| Meloidogyne | 21.43 | 13.46 | 1.61 | 103 | 100 | 28.57 | |
| SC | Helicotylenchus | 83.33 | 78.26 | 2.55 | 2900 | 600 | 33.33 |
| Hemicriconemoides | 66.67 | 47.83 | 1.05 | 25 | - | - | |
| Labrys | 33.33 | 17.39 | 1.20 | 45 | - | - | |
| Mesocriconema | 33.33 | 30.43 | 1.29 | 81 | - | - | |
| Meloidogyne | 16.67 | 34.78 | 1.37 | 105 | 100 | 12.5 |
| Golf Course | Health Status 1 | Plant-Parasitic Nematodes Density (Individuals/100 cm3 of Soil) | Dominance (%) 2 |
|---|---|---|---|
| Beijing Qinghewan | H | 18 | 2.98 |
| P | 33 | 2.78 | |
| Beijing Laffitte | H | 57 | 5.55 |
| P | 114 | 6.04 | |
| Beijing Yuyang | H | 98 | 14.16 |
| P | 89 | 17.87 | |
| Beijing CBD | H | 217 | 27.78 |
| P | 112 | 28.43 | |
| Suzhou Jinji Lake | H | 19 | 2.64 |
| P | 66 | 10.87 | |
| Suzhou Taihu | H | 400 | 27.61 |
| P | 322 | 28.10 | |
| Changzhou Kingswan | H | 112 | 8.85 |
| P | 67 | 6.73 | |
| Nanjing Sun Island | H | 65 | 5.31 |
| P | 333 | 41.73 | |
| Nanjing Zhongshan | H | 241 | 24.69 |
| P | 122 | 15.31 | |
| Shaanxi Yulin Desert | H | 27 | 6.05 |
| P | 91 | 18.80 | |
| Shanghai Sino-Bay | H | 115 | 5.67 |
| P | 127 | 12.10 | |
| Shanghai Links | H | 137 | 10.83 |
| P | 59 | 3.86 | |
| Shanghai Tomson | H | 26 | 6.88 |
| P | 127 | 31.91 | |
| Anhui Huangshan Pine | H | 67 | 9.33 |
| P | 32 | 4.18 | |
| Shenzhou Peninsula | H | 82 | 14.86 |
| P | 22 | 3.50 | |
| Wanning Shanqin Bay | H | 2900 | 61.70 |
| P | 423 | 50.12 | |
| Sanya Luhuitou | H | 5 | 2.76 |
| P | 19 | 10.86 | |
| Shenzhen Sand River | H | 8 | 22.22 |
| P | 108 | 65.85 |
| Region | Number of Courses | Diversity H’ 1 | Richness SR 2 | Evenness J’ 3 | Dominance λ 4 | Diversity H2 5 |
|---|---|---|---|---|---|---|
| NC | 5 | 0.93 ± 0.01 b 6 | 60.98 ± 0.16 c | 0.41 ± 0.04 b | 0.62 ± 0.02 b | 0.48 ± 0.02 b |
| EC | 14 | 1.92 ± 0.01 a | 89.56 ± 0.31 a | 0.77 ± 0.05 a | 0.17 ± 0.01 c | 1.77 ± 0.05 a |
| SC | 6 | 0.42 ± 0.02 c | 88.36 ± 0.19 b | 0.18 ± 0.03 c | 0.85 ± 0.03 a | 0.17 ± 0.03 c |
| Turfgrass Species | Number of Samples | Diversity H’ | Richness SR | Evenness J’ | Dominance λ | Diversity H2 |
|---|---|---|---|---|---|---|
| Creeping bentgrass | 57 | 2.13 ± 0.02 a 2 | 124.72 ± 2.15 a | 0.77 ± 0.04 a | 0.14 ± 0.02 c | 1.97 ± 0.03 a |
| Seashore paspalum | 17 | 0.21 ± 0.02 c | 79.06 ± 0.97 b | 0.09 ± 0.01 c | 0.93 ± 0.02 a | 0.07 ± 0.01 c |
| Hybrid bermudagrass | 13 | 1.36 ± 0.03 b | 42.89 ± 1.16 c | 0.65 ± 0.04 b | 0.33 ± 0.04 b | 1.11 ± 0.05 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Jin, P.; Zhang, H.; Hu, J.; Lamour, K.; Yang, Z. Distribution and Prevalence of Plant-Parasitic Nematodes of Turfgrass at Golf Courses in China. Biology 2022, 11, 1322. https://doi.org/10.3390/biology11091322
Dong Y, Jin P, Zhang H, Hu J, Lamour K, Yang Z. Distribution and Prevalence of Plant-Parasitic Nematodes of Turfgrass at Golf Courses in China. Biology. 2022; 11(9):1322. https://doi.org/10.3390/biology11091322
Chicago/Turabian StyleDong, Yinglu, Peiyuan Jin, Huangwei Zhang, Jian Hu, Kurt Lamour, and Zhimin Yang. 2022. "Distribution and Prevalence of Plant-Parasitic Nematodes of Turfgrass at Golf Courses in China" Biology 11, no. 9: 1322. https://doi.org/10.3390/biology11091322
APA StyleDong, Y., Jin, P., Zhang, H., Hu, J., Lamour, K., & Yang, Z. (2022). Distribution and Prevalence of Plant-Parasitic Nematodes of Turfgrass at Golf Courses in China. Biology, 11(9), 1322. https://doi.org/10.3390/biology11091322