

Identification of Putative Neuropeptides That Alter the Behaviour of Schistosoma mansoni Cercariae

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biomphalaria glabrata Maintenance Conditions and Ethics Guidelines
2.2. Schistosoma mansoni Cercaria Collection and Peptide Isolation
2.3. uHPLC Tandem QTOF MS/MS Analyses
2.4. Protein Identification
2.5. Prediction of Putative Neuropeptides, Gene Ontology, and KEGG Pathway Analysis
2.6. Protein–Protein Interaction (PPI) Network
2.7. Comparative Sequence Analysis of Putative Neuropeptides
2.8. Peptide Synthesis and Preparation for Bioassay
2.9. Schistosoma mansoni Cercarial Behavioural Bioassays
2.10. Behavioural Bioassay Data Analysis
2.10.1. Analysis of the Characteristics of Cercarial Movement
2.10.2. Prolonged Exposure Analysis
2.11. Secondary Structure Analysis
3. Results
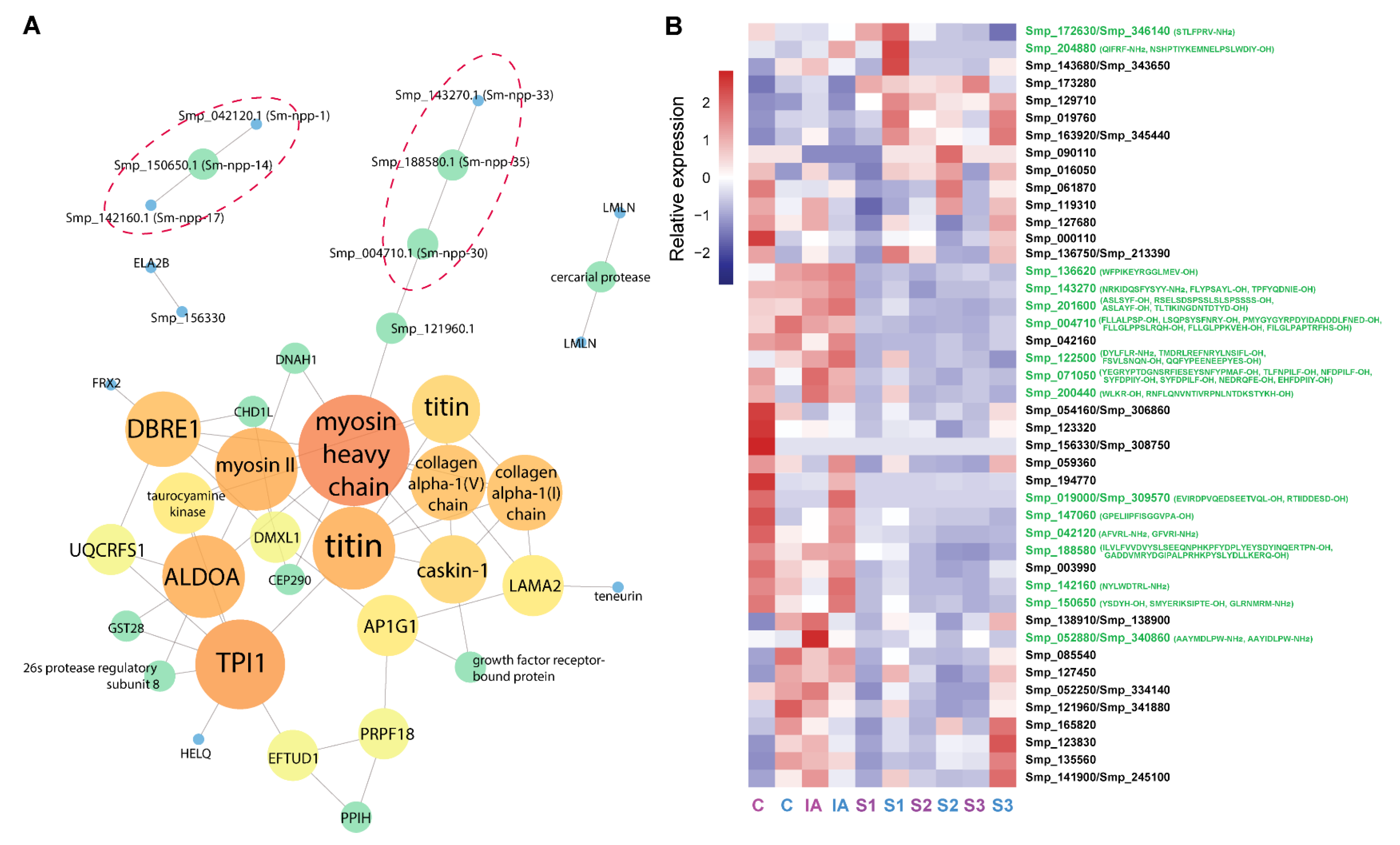
3.1. Putative Neuropeptide Precursors Identified in Cercariae
3.2. Protein–Protein Interaction Analysis
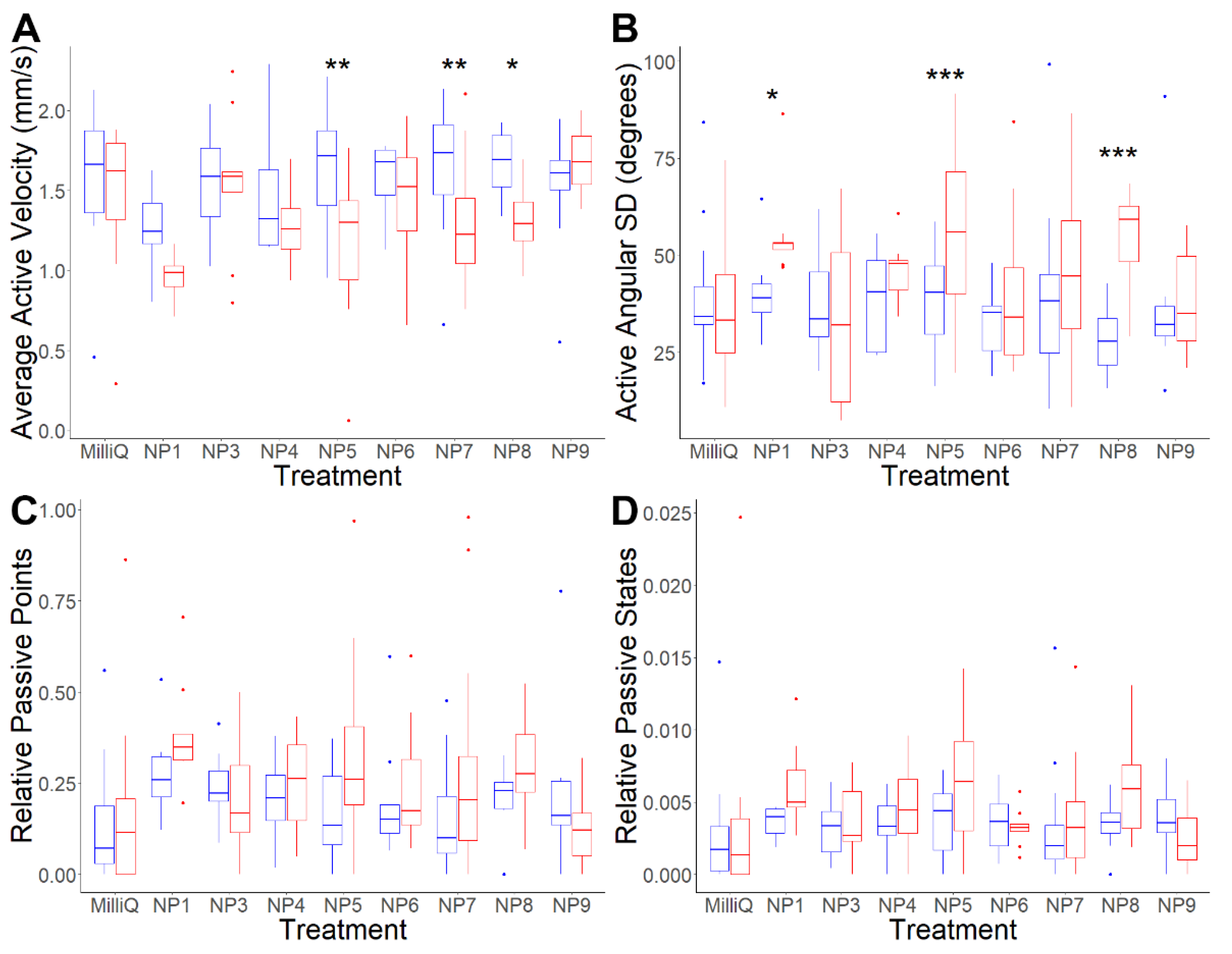
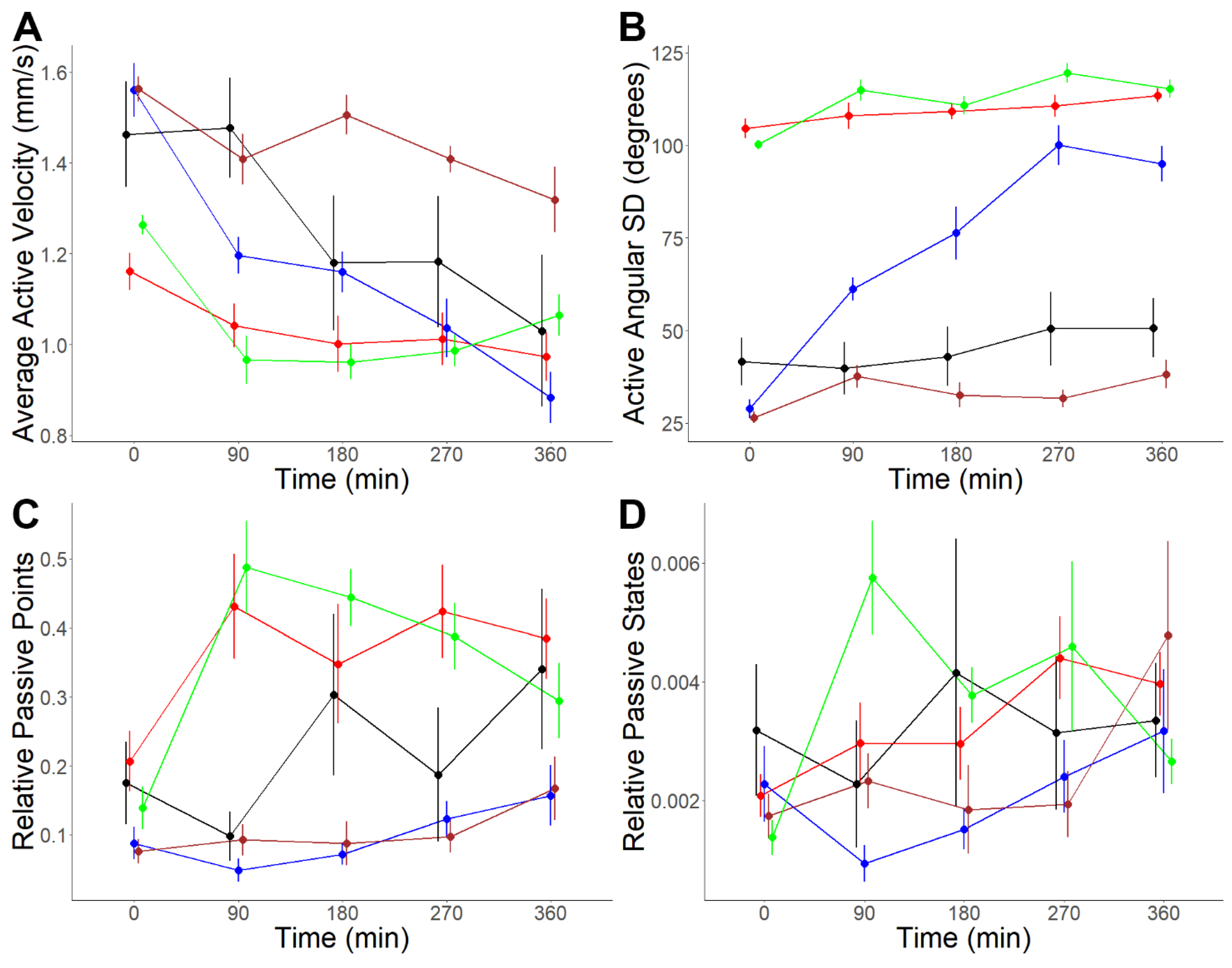
3.3. Analysis of Putative Neuropeptides on Schistosoma mansoni Cercaria Behaviour
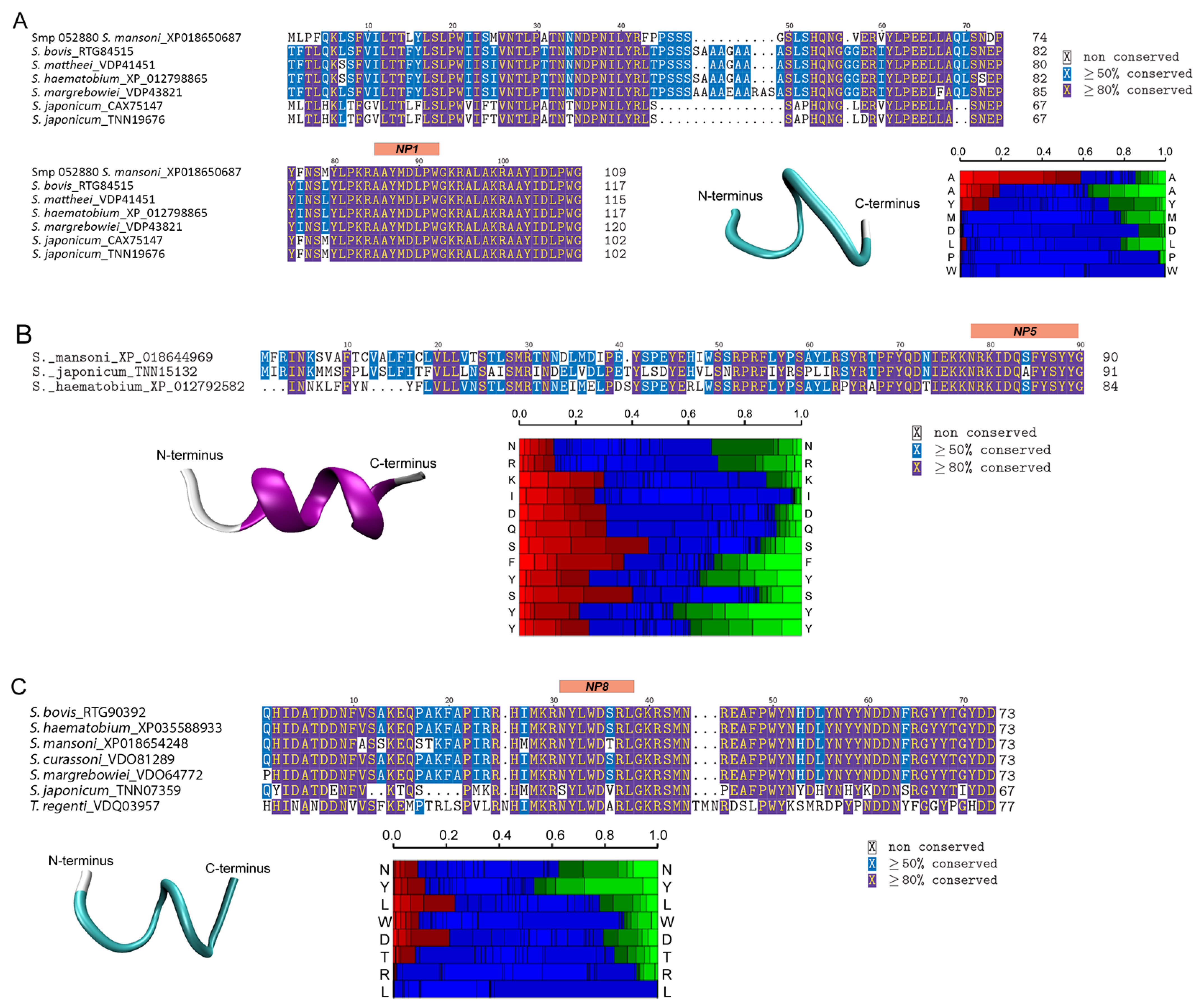
3.4. Structural Analysis of Bioactive Putative Neuropeptides That Modify Schistosoma mansoni Cercaria Behaviour
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ribeiro, A.R.; Luis, C.; Fernandes, R.; Botelho, M.C. Schistosomiasis and Infertility: What Do We Know? Trends Parasitol. 2019, 35, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Naser, M.B.; Altenburg, A.; Zouboulis, C.C.; Wollina, U. Schistosomiasis (bilharziasis) and male infertility. Andrologia 2019, 51, e13165. [Google Scholar] [CrossRef] [PubMed]
- Brodish, P.H.; Singh, K. Association between Schistosoma haematobium exposure and human immunodeficiency virus infection among females in Mozambique. Am. J. Trop. Med. Hyg. 2016, 94, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.P.; Dunne, D.W.; Sacko, M.; Utzinger, J.; Vennervald, B.J.; Zhou, X.N. Schistosomiasis. Nat. Rev. Dis. Prim. 2018, 4. [Google Scholar] [CrossRef]
- Knowles, S.C.L. The effect of helminth co-infection on malaria in mice: A meta-analysis. Int. J. Parasitol. 2011, 41, 1041–1051. [Google Scholar] [CrossRef]
- Clark, E.H.; Serpa, J.A. Tissue Parasites in HIV Infection. Curr. Infect. Dis. Rep. 2019, 21, 49. [Google Scholar] [CrossRef]
- Mello-Silva, C.C.; João, R.C.; Augusto, R.C.; Santos, C.P. A rapid diagnostic test for Schistosomiasis mansoni. Mem. Inst. Oswaldo Cruz 2013, 108, 1078–1080. [Google Scholar] [CrossRef]
- Olveda, D.U.; McManus, D.P.; Ross, A.G.P. Mass drug administration and the global control of schistosomiasis: Successes, limitations and clinical outcomes. Curr. Opin. Infect. Dis. 2016, 29, 595–608. [Google Scholar] [CrossRef]
- da Silva, V.B.R.; Campos, B.R.K.L.; de Oliveira, J.F.; Decout, J.L.; do Carmo Alves de Lima, M. Medicinal chemistry of antischistosomal drugs: Praziquantel and oxamniquine. Bioorg. Med. Chem. 2017, 25, 3259–3277. [Google Scholar] [CrossRef]
- Gbalégba, N.G.C.; Silué, K.D.; Ba, O.; Ba, H.; Tian-Bi, N.T.Y.; Yapi, G.Y.; Kaba, A.; Koné, B.; Utzinger, J.; Koudou, B.G. Prevalence and seasonal transmission of Schistosoma haematobium infection among school-aged children in Kaedi town, southern Mauritania. Parasites Vectors 2017, 10, 353. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Dong, H.F.; Guo, Y.; Zhao, Q.P.; Jiang, M.S. Efficacy of praziquantel and artemisinin derivatives for the treatment and prevention of human schistosomiasis: A systematic review and meta-analysis. Parasites Vectors 2011, 4, 201. [Google Scholar] [CrossRef] [PubMed]
- Gurarie, D.; Lo, N.C.; Ndeffo-Mbah, M.L.; Durham, D.P.; King, C.H. The human-snail transmission environment shapes long term schistosomiasis control outcomes: Implications for improving the accuracy of predictive modeling. PLoS Negl. Trop. Dis. 2018, 12, e0006514. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.M.; Neimark, H.; Eveland, L. Schistosoma mansoni: Response of cercariae to a thermal gradient. J. Parasitol. 1980, 66, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Hansell, E.; Braschi, S.; Medzihradszky, K.F.; Sajid, M.; Debnath, M.; Ingram, J.; Lim, K.C.; McKerrow, J.H. Proteomic analysis of skin invasion by blood fluke larvae. PLoS Negl. Trop. Dis. 2008, 2, e0000262. [Google Scholar] [CrossRef]
- Haas, W.; Grabe, K.; Geis, C.; Päch, T.; Stoll, K.; Fuchs, M.; Haberl, B.; Loy, C. Recognition and invasion of human skin by Schistosoma mansoni cercariae: The key-role of L-arginine. Parasitology 2002, 124, 153–167. [Google Scholar] [CrossRef]
- Berriman, M.; Haas, B.J.; LoVerde, P.T.; Wilson, R.A.; Dillon, G.P.; Cerqueira, G.C.; Mashiyama, S.T.; Al-Lazikani, B.; Andrade, L.F.; Ashton, P.D.; et al. The genome of the blood fluke Schistosoma mansoni. Nature 2009, 460, 352–358. [Google Scholar] [CrossRef]
- Koziol, U.; Koziol, M.; Preza, M.; Costábile, A.; Brehm, K.; Castillo, E. De novo discovery of neuropeptides in the genomes of parasitic flatworms using a novel comparative approach. Int. J. Parasitol. 2016, 46, 709–721. [Google Scholar] [CrossRef]
- Hökfelt, T.; Broberger, C.; Xu, Z.Q.; Sergeyev, V.; Ubink, R.; Diez, M. Neuropeptides—An overview. Neuropharmacology 2000, 39, 1337–1356. [Google Scholar] [CrossRef]
- Buddenborg, S.K.; Kamel, B.; Hanelt, B.; Bu, L.; Zhang, S.M.; Mkoji, G.M.; Loker, E.S. The in vivo transcriptome of Schistosoma mansoni in the prominent vector species Biomphalaria pfeifferi with supporting observations from Biomphalaria glabrata. PLoS Negl. Trop. Dis. 2019, 13, e0007013. [Google Scholar] [CrossRef]
- Wang, T.; Zhao, M.; Rotgans, B.A.; Strong, A.; Liang, D.; Ni, G.; Limpanont, Y.; Ramasoota, P.; McManus, D.P.; Cummins, S.F. Proteomic analysis of the schistosoma mansoni miracidium. PLoS ONE 2016, 11, e0147247. [Google Scholar] [CrossRef] [Green Version]
- McVeigh, P.; Kimber, M.J.; Novozhilova, E.; Day, T.A. Neuropeptide signalling systems in flatworms. Parasitology 2005, 131 (Suppl. S1), S41–S55. [Google Scholar] [CrossRef] [PubMed]
- McVeigh, P.; Atkinson, L.; Marks, N.J.; Mousley, A.; Dalzell, J.J.; Sluder, A.; Hammerland, L.; Maule, A.G. Parasite neuropeptide biology: Seeding rational drug target selection? Int. J. Parasitol. Drugs Drug Resist. 2012, 2, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yang, N.; Lewis, F.A.; Yau, P.; Collins, J.J., 3rd; Sweedler, J.V.; Newmark, P.A. A rotifer-derived paralytic compound prevents transmission of schistosomiasis to a mammalian host. PLoS Biol. 2019, 17, e3000485. [Google Scholar] [CrossRef] [PubMed]
- McVeigh, P.; Mair, G.R.; Novozhilova, E.; Day, A.; Zamanian, M.; Marks, N.J.; Kimber, M.J.; Day, T.A.; Maule, A.G. Schistosome I/Lamides--a new family of bioactive helminth neuropeptides. Int. J. Parasitol. 2011, 41, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Chieu, H.D.; Turner, L.; Smith, M.K.; Wang, T.; Nocillado, J.; Palma, P.; Suwansa-Ard, S.; Elizur, A.; Cummins, S.F. Aquaculture Breeding Enhancement: Maturation and Spawning in Sea Cucumbers Using a Recombinant Relaxin-Like Gonad-Stimulating Peptide. Front. Genet. 2019, 10, 77. [Google Scholar] [CrossRef]
- Chambers, M.C.; MacLean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Hiller, K.; Grote, A.; Scheer, M.; Munch, R.; Jahn, D. PrediSi: Prediction of signal peptides and their cleavage positions. Nucleic Acids Res. 2004, 32, W375–W379. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Southey, B.R.; Amare, A.; Zimmerman, T.A.; Rodriguez-Zas, S.L.; Sweedler, J.V. NeuroPred: A tool to predict cleavage sites in neuropeptide precursors and provide the masses of the resulting peptides. Nucleic Acids Res. 2006, 34, W267–W272. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, C.E.; Zhao, M.; McManus, D.P.; Duke, M.G.; Cummins, S.F.; Wang, T. Comparative study of excretory-secretory proteins released by Schistosoma mansoni-resistant, susceptible and naïve Biomphalaria glabrata. Parasites Vectors 2019, 12, 452. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Picard, M.A.; Boissier, J.; Roquis, D.; Grunau, C.; Allienne, J.F.; Duval, D.; Toulza, E.; Arancibia, N.; Caffrey, C.R.; Long, T.; et al. Sex-Biased Transcriptome of Schistosoma mansoni: Host-Parasite Interaction, Genetic Determinants and Epigenetic Regulators Are Associated with Sexual Differentiation. PLoS Negl. Trop. Dis. 2016, 10, e0004930. [Google Scholar] [CrossRef]
- Wyeth, R.C.; Braubach, O.R.; Fine, A.; Croll, R.P. Videograms: A method for repeatable unbiased quantitative behavioral analysis without scoring or tracking. Neuromethods 2011, 51, 15–33. [Google Scholar] [CrossRef]
- Tinevez, J.Y.; Perry, N.; Schindelin, J.; Hoopes, G.M.; Reynolds, G.D.; Laplantine, E.; Bednarek, S.Y.; Shorte, S.L.; Eliceiri, K.W. TrackMate: An open and extensible platform for single-particle tracking. Methods 2017, 115, 80–90. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Meijering, E.; Dzyubachyk, O.; Smal, I. Methods for cell and particle tracking. Methods Enzymol. 2012, 504, 183–200. [Google Scholar] [CrossRef]
- Brachs, S.; Haas, W. Swimming behaviour of Schistosoma mansoni cercariae: Responses to irradiance changes and skin attractants. Parasitol. Res. 2008, 102, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The aligned rank transform for nonparametric factorial analyses using only anova procedures. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Vancouver, BC, Canada, 7–12 May 2011; pp. 143–146. [Google Scholar]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. In Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology, Virtual Event, USA, 10–14 October 2021; pp. 754–768. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio: Boston, MA, USA, 2021. [Google Scholar]
- Wickham, H.; Bryan, J. readxl: Read Excel Files. 2019. Available online: https://CRAN.R-project.org/package=readxl (accessed on 21 January 2021).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; D’Agostino, L. Welcome to the {tidyverse}. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Bache, S.M.; Wickham, H. magrittr: A Forward-Pipe Operator for R. 2020. Available online: https://CRAN.R-project.org/package=magrittr (accessed on 21 January 2021).
- Wickham, H. forcats: Tools for Working with Categorical Variables (Factors). 2021. Available online: https://CRAN.R-project.org/package=forcats (accessed on 21 January 2021).
- Bates, D.; Machler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using {lme4}. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). 2020. Available online: https://CRAN.R-project.org/package=AICcmodavg (accessed on 21 January 2021).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Lemon, J. Plotrix: A package in the red light district of R. R-News 2006, 6, 8–12. [Google Scholar]
- Kay, M.; Elkin, L.A.; Higgins, J.J.; Wobbrock, J.O. ARTool: Aligned Rank Transform for Nonparametric Factorial ANOVAs. Available online: https://github.com/mjskay/ARTool (accessed on 21 January 2021).
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 21 January 2021).
- Shen, Y.; Maupetit, J.; Derreumaux, P.; Tufféry, P. Improved PEP-FOLD Approach for Peptide and Miniprotein Structure Prediction. J. Chem. Theory Comput. 2014, 10, 4745–4758. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Collins, J.J., 3rd; Hou, X.; Romanova, E.V.; Lambrus, B.G.; Miller, C.M.; Saberi, A.; Sweedler, J.V.; Newmark, P.A. Genome-wide analyses reveal a role for peptide hormones in planarian germline development. PLoS Biol. 2010, 8, e1000509. [Google Scholar] [CrossRef]
- Krishnamurthy, D.; Katsikis, G.; Bhargava, A.; Prakash, M. Schistosoma mansoni cercariae swim efficiently by exploiting an elastohydrodynamic coupling. Nat. Phys. 2017, 13, 266–271. [Google Scholar] [CrossRef]
- McCoy, C.J.; Atkinson, L.E.; Zamanian, M.; McVeigh, P.; Day, T.A.; Kimber, M.J.; Marks, N.J.; Maule, A.G.; Mousley, A. New insights into the FLPergic complements of parasitic nematodes: Informing deorphanisation approaches. EuPA Open Proteom. 2014, 3, 262–272. [Google Scholar] [CrossRef]
- Cabrele, C.; Beck-Sickinger, A.G. Molecular characterization of the ligand-receptor interaction of the neuropeptide Y family. J. Pept. Sci. Off. Publ. Eur. Pept. Soc. 2000, 6, 97–122. [Google Scholar] [CrossRef]
- Delgado, M.; Anderson, P.; Garcia-Salcedo, J.A.; Caro, M.; Gonzalez-Rey, E. Neuropeptides kill African trypanosomes by targeting intracellular compartments and inducing autophagic-like cell death. Cell Death Differ. 2009, 16, 406–416. [Google Scholar] [CrossRef] [PubMed]
- van Grondelle, W.; Iglesias, C.L.; Coll, E.; Artzner, F.; Paternostre, M.; Lacombe, F.; Cardus, M.; Martinez, G.; Montes, M.; Cherif-Cheikh, R.; et al. Spontaneous fibrillation of the native neuropeptide hormone Somatostatin-14. J. Struct. Biol. 2007, 160, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Haas, W.; Haberl, B.; Schmalfuss, G.; Khayyal, M.T. Schistosoma haematobium cercarial host-finding and host-recognition differs from that of S. mansoni. J. Parasitol. 1994, 80, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, A.; de Wit, J. Leucine-rich repeat-containing synaptic adhesion molecules as organizers of synaptic specificity and diversity. Exp. Mol. Med. 2018, 50, 10. [Google Scholar] [CrossRef]
- Howard, S.R.; Guasti, L.; Ruiz-Babot, G.; Mancini, A.; David, A.; Storr, H.L.; Metherell, L.A.; Sternberg, M.J.; Cabrera, C.P.; Warren, H.R.; et al. IGSF10 mutations dysregulate gonadotropin-releasing hormone neuronal migration resulting in delayed puberty. EMBO Mol. Med. 2016, 8, 626–642. [Google Scholar] [CrossRef]
- Sulbarán, G.; Alamo, L.; Pinto, A.; Márquez, G.; Méndez, F.; Padrón, R.; Craig, R. An invertebrate smooth muscle with striated muscle myosin filaments. Proc. Natl. Acad. Sci. USA 2015, 112, E5660–E5668. [Google Scholar] [CrossRef]
- Zhang, Y.; Taylor, M.G.; Bickle, Q.D. Schistosoma japonicum myosin: Cloning, expression and vaccination studies with the homologue of the S. mansoni myosin fragment IrV-5. Parasite Immunol. 1998, 20, 583–594. [Google Scholar] [CrossRef]
- Benian, G.M.; Ayme-Southgate, A.; Tinley, T.L. The genetics and molecular biology of the titin/connectin-like proteins of invertebrates. Rev. Physiol. Biochem. Pharmacol. 1999, 138, 235–268. [Google Scholar] [CrossRef]
- Labeit, S.; Kolmerer, B. Titins: Giant proteins in charge of muscle ultrastructure and elasticity. Science 1995, 270, 293–296. [Google Scholar] [CrossRef]
- Camacho, M.; Alsford, S.; Agnew, A. Molecular forms of tegumental and muscle acetylcholinesterases of Schistosoma. Parasitology 1996, 112 Pt 2, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Zhao, M.; Wang, T.; McManus, D.P.; Cummins, S.F. GPCR and IR genes in Schistosoma mansoni miracidia. Parasit Vectors 2016, 9, 563. [Google Scholar] [CrossRef]
- You, J.; Pan, F.; Wang, S.; Wang, Y.; Hu, Y. FMRFamide-Like Peptide 22 Influences the Head Movement, Host Finding, and Infection of Heterodera glycines. Front. Plant Sci. 2021, 12, 673354. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sequence | Molecular Weight (Da) | Accession of Precursor |
|---|---|---|---|
| NP1 | AAYMDLPW-NH2 | 965.131 | Smp_052880.1 (Sm-npp-5) |
| NP2 | SYFDPILF-OH | 1001.137 | Smp_071050.1 (Sm-npp-26) |
| NP3 | STLFPRV-NH2 | 817.979 | Smp_172630.1 (Sm-npp-38) |
| NP4 | GLRNMRM-NH2 | 876.110 | Smp_150650.1 (Sm-npp-14) |
| NP5 | NRKIDQSFYSYY-NH2 | 1582.721 | Smp_143270.1 (Sm-npp-33) |
| NP6 | ASLSYF-OH | 686.756 | Smp_201600.1 (Sm-npp-36) |
| NP7 | FLLALPSP-OH | 857.052 | Smp_004710.1 (Sm-npp-30) |
| NP8 | NYLWDTRL-NH2 | 1079.214 | Smp_142160.1 (Sm-npp-17) |
| NP9 | YIRF-NH2 | 596.724 | Smp_188580.1 (Sm-npp-35) |
| Metrics | Effect | Statistic | df | p-Value | MQ: NP1 | MQ: NP3 | MQ: NP4 | MQ: NP5 | MQ: NP6 | MQ: NP7 | MQ: NP8 | MQ: NP9 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Active velocity | Neuropeptide | 2.98 | 8 | 0.0049 | ||||||||
| Period | 53.69 | 1 | <0.0001 | |||||||||
| Neuropeptide:Period | 4.79 | 8 | <0.0001 | 0.0793 | 0.4597 | 0.6763 | 0.0011 | 0.9736 | 0.0025 | 0.0315 | 0.1182 | |
| Active angular SD | Neuropeptide | 2.05 | 8 | 0.0480 | ||||||||
| Period | 22.35 | 1 | <0.0001 | |||||||||
| Neuropeptide:Period | 3.67 | 8 | 0.0009 | 0.0120 | 0.9753 | 0.1447 | 0.0009 | 0.3849 | 0.0959 | <0.0001 | 0.2574 | |
| Relative passive points | Neuropeptide | 2.91 | 8 | 0.0058 | ||||||||
| Period | 12.50 | 1 | 0.0006 | |||||||||
| Neuropeptide:Period | 1.55 | 8 | 0.1496 | - | - | - | - | - | - | - | - | |
| Relative passive states | Neuropeptide | 3.33 | 8 | 0.0020 | ||||||||
| Period | 7.13 | 1 | 0.0088 | |||||||||
| Neuropeptide:Period | 1.21 | 8 | 0.3017 | - | - | - | - | - | - | - | - |
| Metrics | Effect | Statistic | df | p-Value | Interaction | 0–90 | 0–180 | 0–270 | 0–360 |
|---|---|---|---|---|---|---|---|---|---|
| Active velocity | Neuropeptide | 30.64 | 4 | <0.0001 | |||||
| Period | 21.84 | 4 | <0.0001 | ||||||
| Neuropeptide:Period | 2.93 | 16 | 0.0003 | NP1:MilliQ | 0.5295 | 0.7231 | 0.5637 | 0.3432 | |
| NP5:MilliQ | 0.0055 | 0.0694 | 0.0065 | 0.0014 | |||||
| NP7:MilliQ | 0.2462 | 0.1611 | 0.4193 | 0.7179 | |||||
| NP8:MilliQ | 0.0198 | 0.2809 | 0.5734 | 0.4158 | |||||
| Active angular SD | Neuropeptide | 181.59 | 4 | <0.0001 | |||||
| Period | 29.43 | 4 | <0.0001 | ||||||
| Neuropeptide:Period | 7.67 | 16 | <0.0001 | NP1:MilliQ | 0.6446 | 0.8506 | 0.3824 | 0.4818 | |
| NP5:MilliQ | 0.0008 | <0.0001 | <0.0001 | <0.0001 | |||||
| NP7:MilliQ | 0.1627 | 0.9820 | 0.2415 | 0.6917 | |||||
| NP8:MilliQ | 0.0384 | 0.4700 | 0.3075 | 0.9527 | |||||
| Relative passive points | Neuropeptide | 20.60 | 4 | <0.0001 | |||||
| Period | 6.47 | 4 | <0.0001 | ||||||
| Neuropeptide:Period | 3.00 | 16 | 0.0002 | NP1:MilliQ | 0.0038 | 0.3417 | 0.0144 | 0.3564 | |
| NP5:MilliQ | 0.8690 | 0.2362 | 0.7278 | 0.4844 | |||||
| NP7:MilliQ | 0.5104 | 0.3731 | 0.9642 | 0.6251 | |||||
| NP8:MilliQ | 0.0001 | 0.0031 | 0.0020 | 0.5326 | |||||
| Relative passive states | Neuropeptide | 3.68 | 4 | 0.0121 | |||||
| Period | 2.44 | 4 | 0.0492 | ||||||
| Neuropeptide:Period | 2.06 | 16 | 0.0127 | NP1:MilliQ | 0.2428 | 0.2149 | 0.1153 | 0.4699 | |
| NP5:MilliQ | 0.6458 | 0.9770 | 0.9498 | 0.6564 | |||||
| NP7:MilliQ | 0.2899 | 0.8596 | 0.8572 | 0.6859 | |||||
| NP8:MilliQ | 0.0004 | 0.0076 | 0.0702 | 0.7113 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fogarty, C.E.; Suwansa-ard, S.; Phan, P.; McManus, D.P.; Duke, M.G.; Wyeth, R.C.; Cummins, S.F.; Wang, T. Identification of Putative Neuropeptides That Alter the Behaviour of Schistosoma mansoni Cercariae. Biology 2022, 11, 1344. https://doi.org/10.3390/biology11091344
Fogarty CE, Suwansa-ard S, Phan P, McManus DP, Duke MG, Wyeth RC, Cummins SF, Wang T. Identification of Putative Neuropeptides That Alter the Behaviour of Schistosoma mansoni Cercariae. Biology. 2022; 11(9):1344. https://doi.org/10.3390/biology11091344
Chicago/Turabian StyleFogarty, Conor E., Saowaros Suwansa-ard, Phong Phan, Donald P. McManus, Mary G. Duke, Russell C. Wyeth, Scott F. Cummins, and Tianfang Wang. 2022. "Identification of Putative Neuropeptides That Alter the Behaviour of Schistosoma mansoni Cercariae" Biology 11, no. 9: 1344. https://doi.org/10.3390/biology11091344
APA StyleFogarty, C. E., Suwansa-ard, S., Phan, P., McManus, D. P., Duke, M. G., Wyeth, R. C., Cummins, S. F., & Wang, T. (2022). Identification of Putative Neuropeptides That Alter the Behaviour of Schistosoma mansoni Cercariae. Biology, 11(9), 1344. https://doi.org/10.3390/biology11091344