
Identification and Functional Analysis of the Cell Proliferation Regulator, Insulin-like Growth Factor 1 (IGF1) in Freshwater Pearl Mussel (Hyriopsis cumingii)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Tissue Samples
2.2. RNA Isolation and cDNA Synthesis
2.3. Cloning of Full-Length cDNA
2.4. Bioinformatics Analysis
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.6. In Situ Hybridization
2.7. Synthesis of dsRNA In Vitro and In Vivo Interference
2.8. Prokaryotic Expression and Activity Analysis of HcIGF1
2.9. Statistical Analysis
3. Results
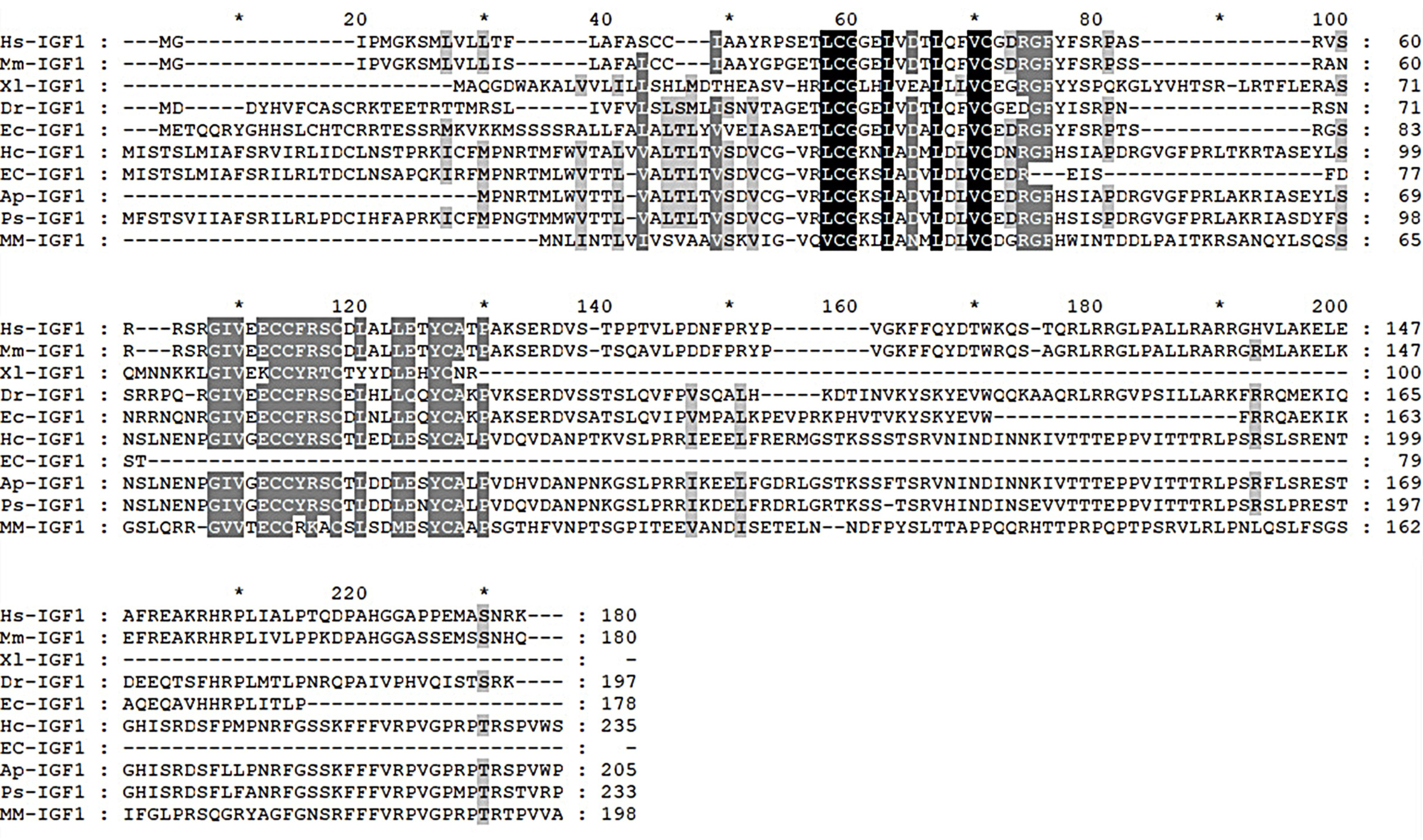
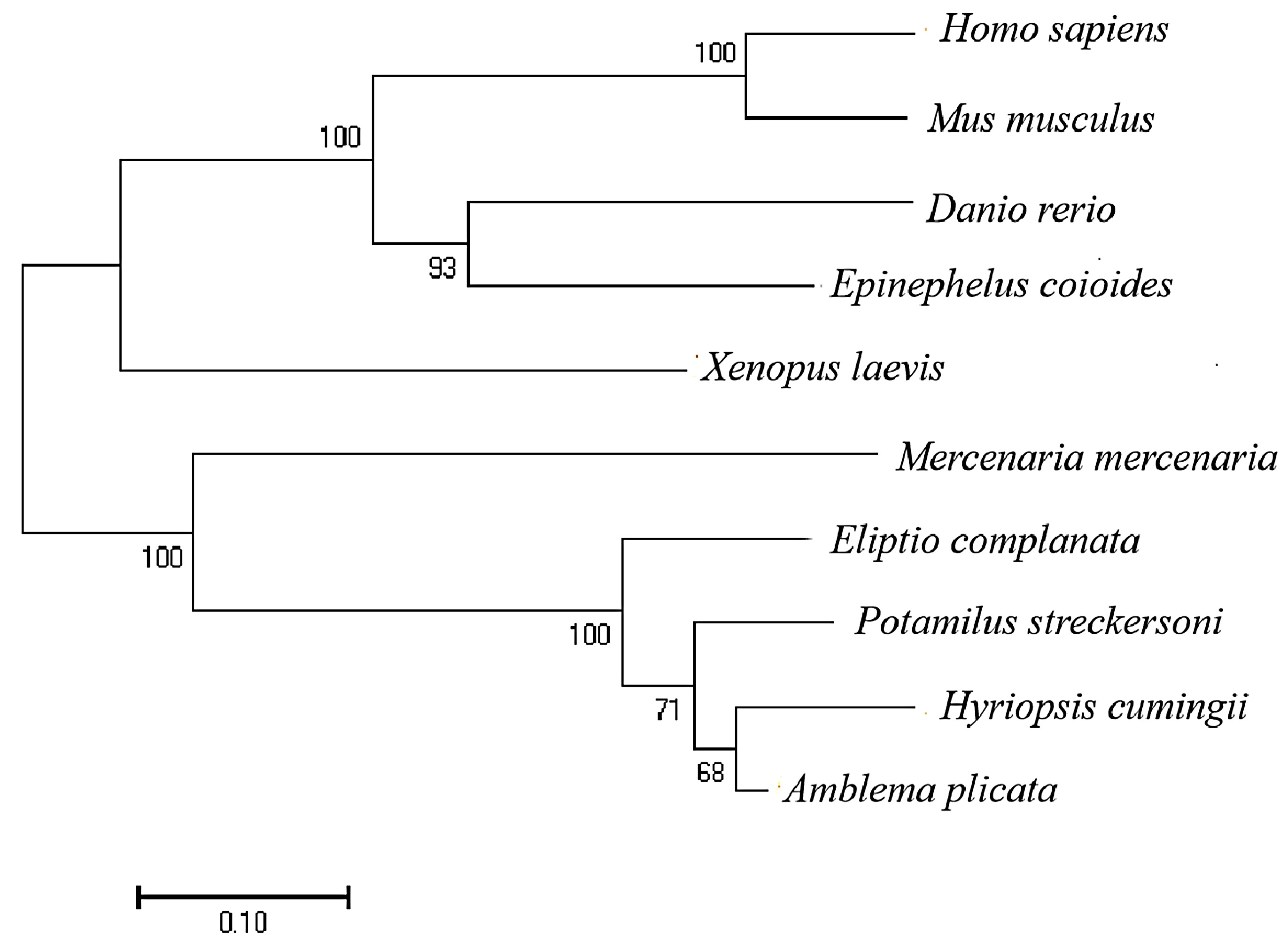
3.1. Bioinformatics Analysis of HcIGF1
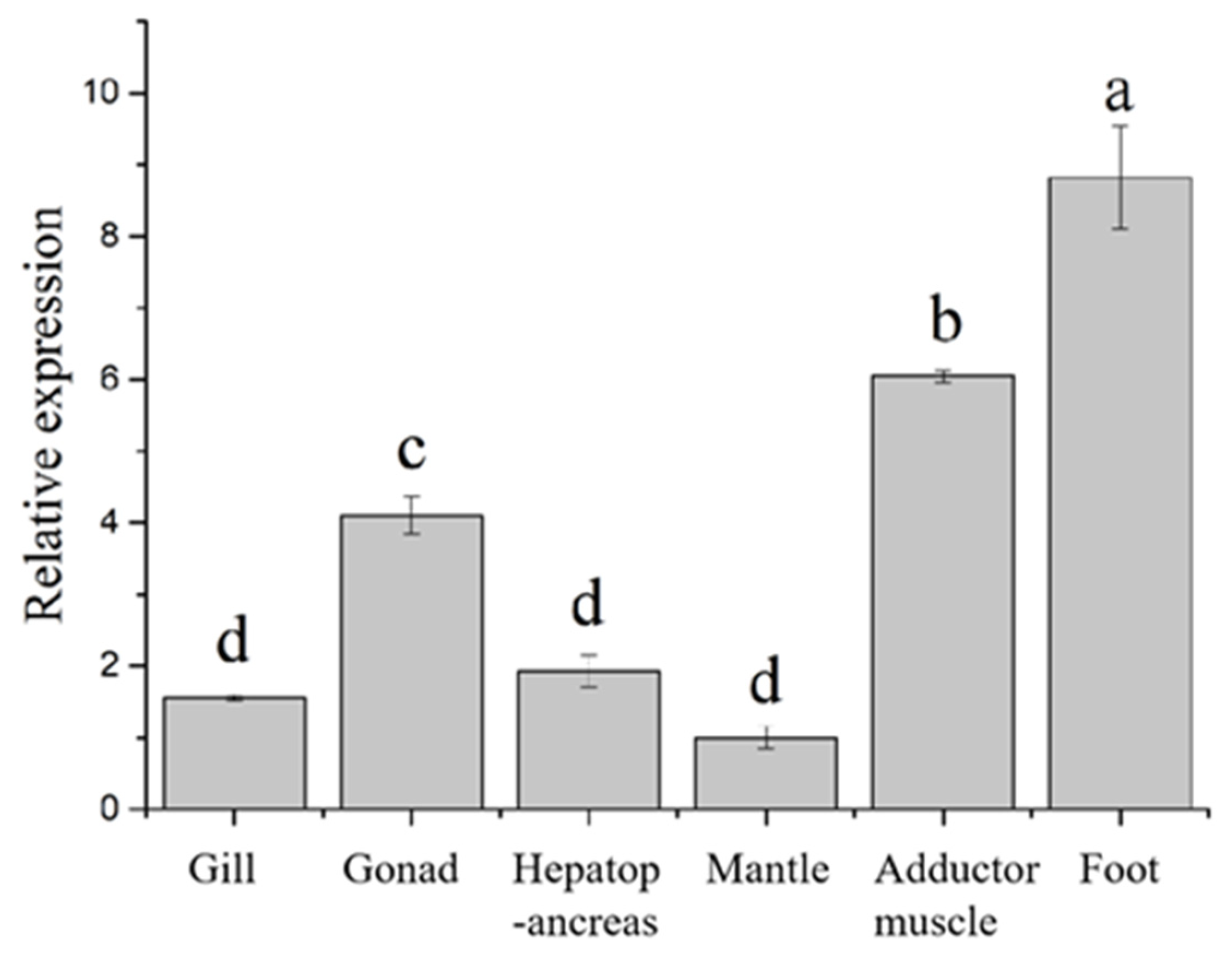
3.2. HcIGF1 Expression in Different Tissues
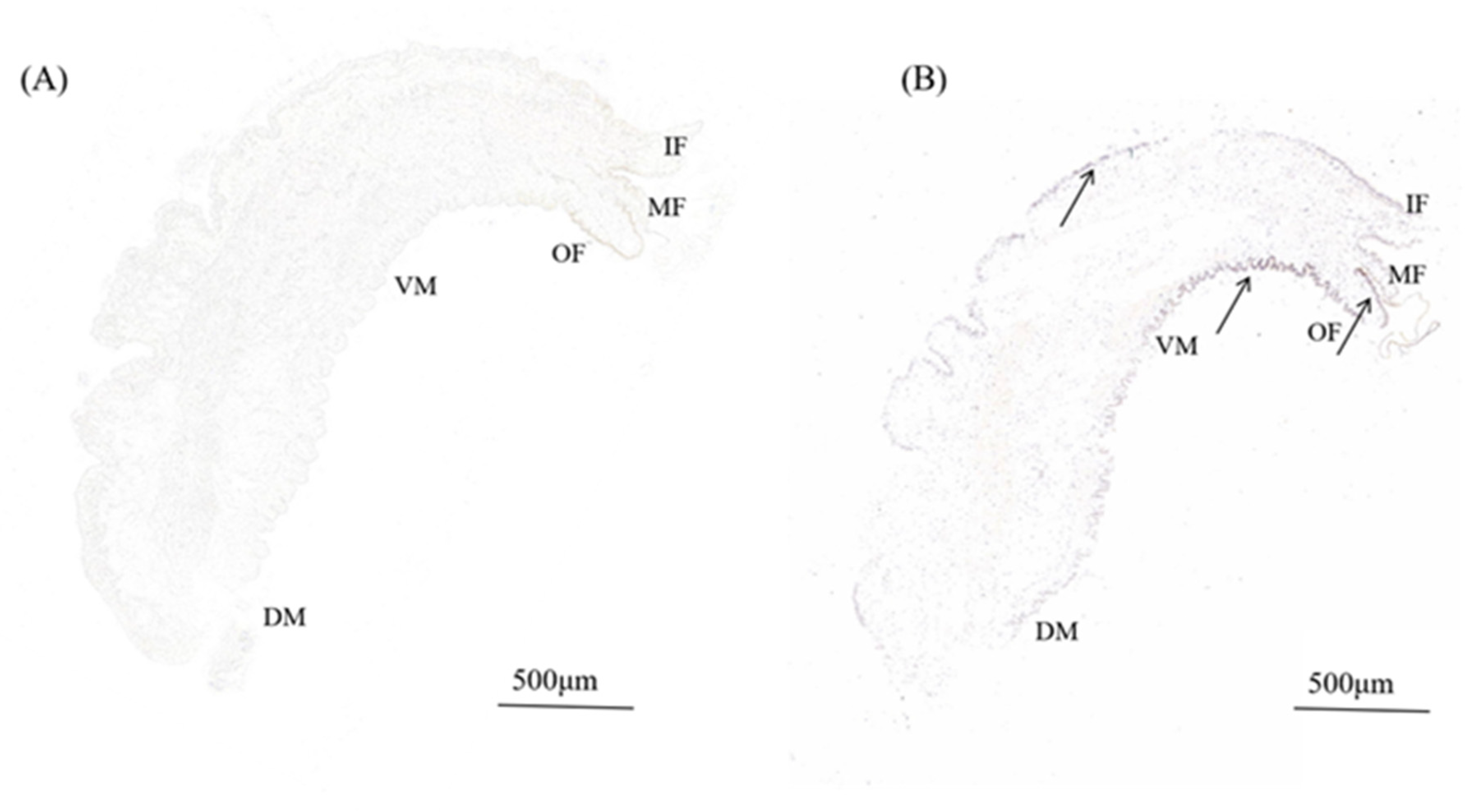
3.3. In Situ Hybridization
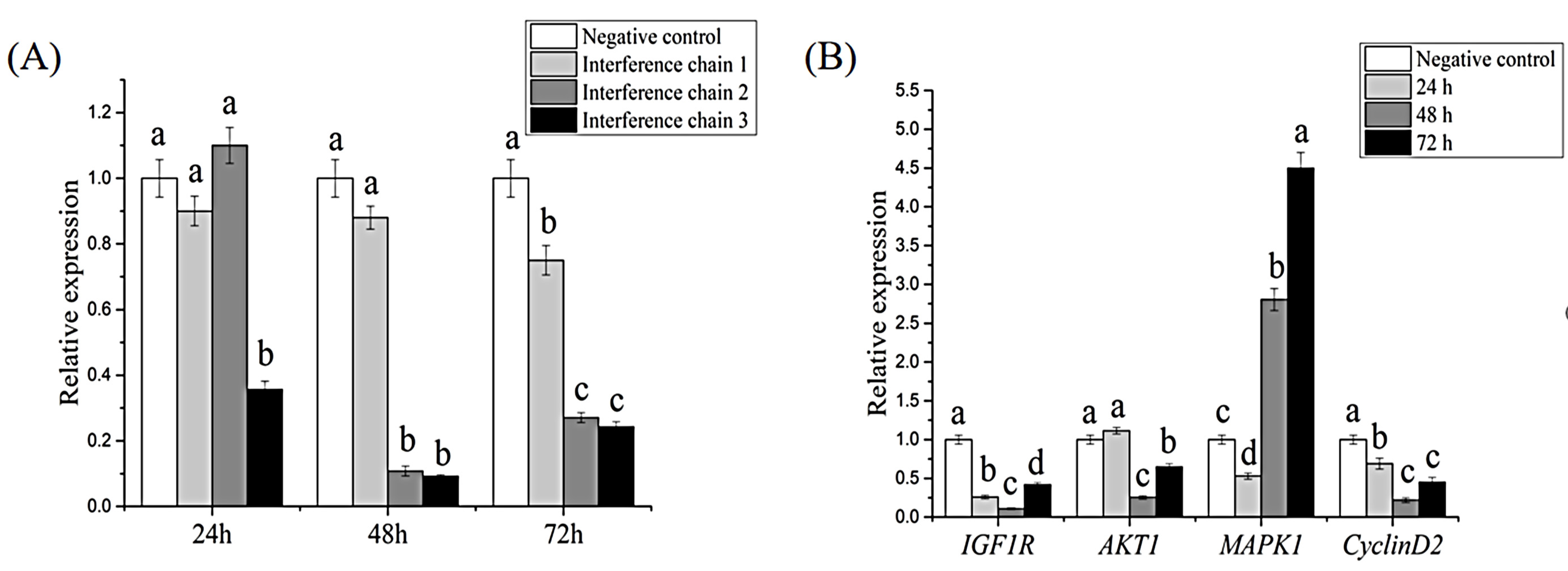
3.4. RNAi and Effects on Downstream Genes
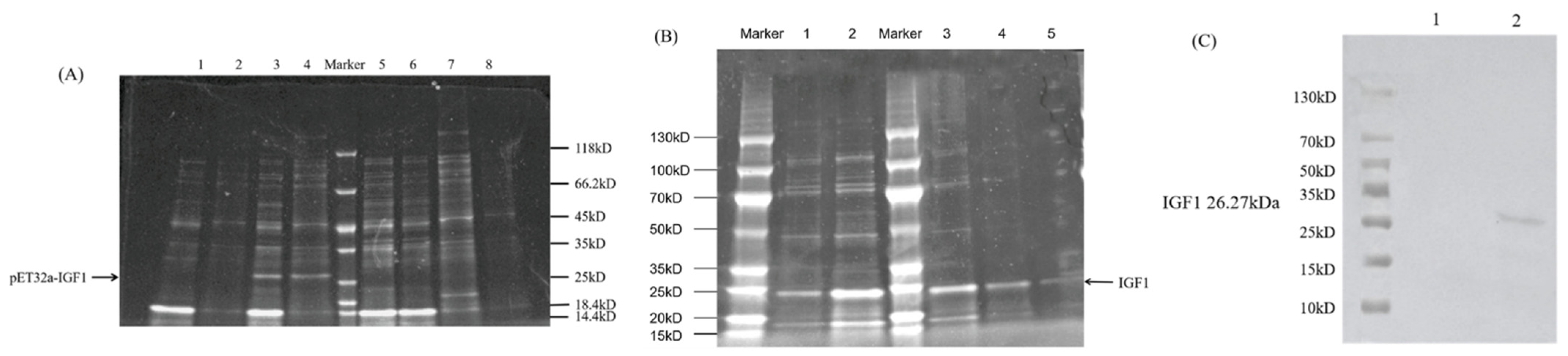
3.5. Prokaryotic Expression and Purification of HcIGF1 Fusion Protein
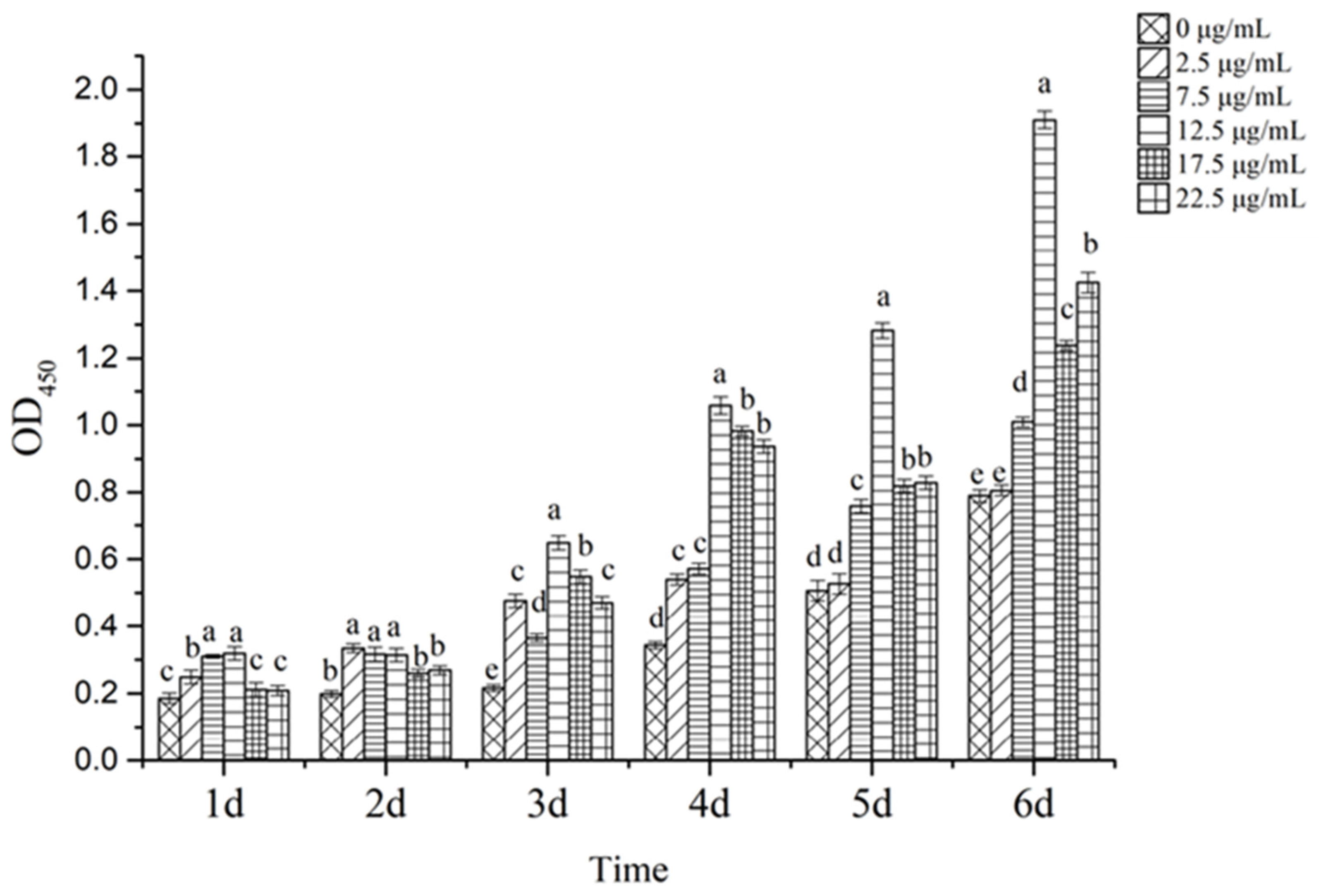
3.6. Identification of Biological Activity of Recombinantly Expressed Proteins
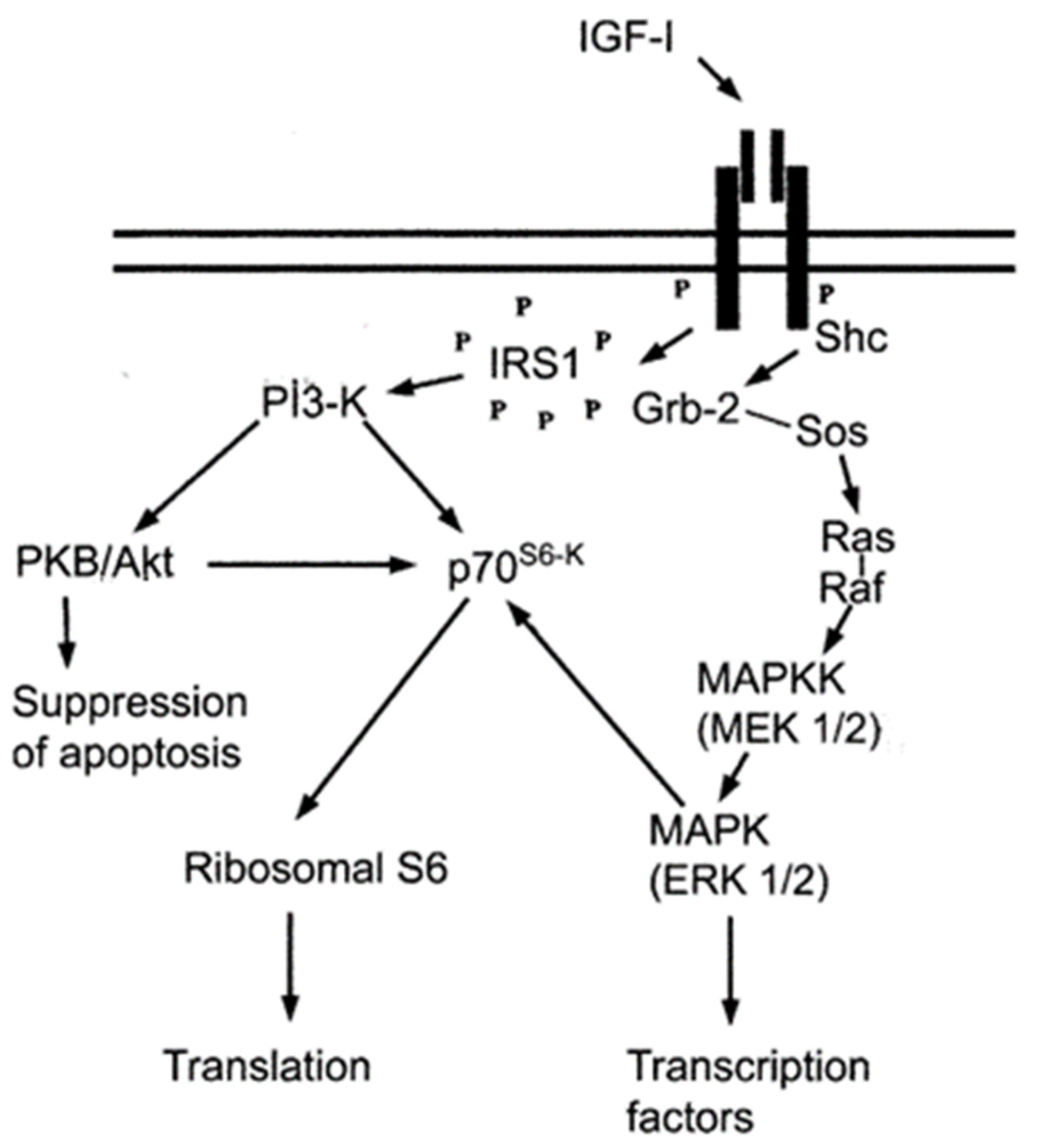
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Mahoney, J.V.; Adams, T.E. Nucleotide sequence of an ovine insulin-like growth factor-II cDNA. Nucleic Acids Res. 1989, 17, 5392. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar] [PubMed]
- Hua, Y.M.; Lin, H.R. Progress of studies on insulin like growth factors. Acta Sci. Nat. Univ. Sunyatseni 1996, S2, 109–114. [Google Scholar]
- Galvin, C.D.; Hardiman, O.; Nolan, C.M. IGF-1 receptor mediates differentiation of primary cultures of mouse skeletal myoblasts. Mol. Cell. Endocrinol. 2003, 200, 19–29. [Google Scholar] [CrossRef]
- Daughaday, W.H.; Rotwein, P. Insulin-like growth factors I and II. Peptide, messenger ribonucleic acid and gene structures, serum, and tissue concentrations. Endocr. Rev. 1989, 10, 68–91. [Google Scholar] [CrossRef]
- Duan, C. Nutritional and developmental regulation of insulin-like growth factors in fish. J. Nutr. 1998, 128, 306S–314S. [Google Scholar] [CrossRef]
- Crane, J.L.; Cao, X. Function of matrix IGF-1 in coupling bone resorption and formation. J. Mol. Med. 2014, 92, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Liu, Q.; He, H.B.; Luo, W. The possible role of insulin-like growth factor-1 in osteosarcoma. Curr. Probl. Cancer 2019, 43, 228–235. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Z.X. Research Progress on the Influence Mechanism of Insulin-like Growth Factors System on Growth Restriction. Acta Acad. Med. Sin. 2011, 33, 18–21. [Google Scholar]
- Lu, J.Y.; Lu, W.; Liu, J.; Lu, J.; Zhai, X.H.; Shen, Z.M. Mechanism of IGF-1 on proliferation of rumen epithelium of goats in vitro. J. Nanjing Agric. Univ. 2014, 37, 106–110. [Google Scholar]
- Cao, Q.P.; Duguay, S.J.; Plisetskaya, E.; Steiner, D.F.; Chan, S.J. Nucleotide sequence and growth hormone-regulated expression of salmon insulin-like growth factor I mRNA. Mol. Endocrinol. 1989, 3, 2005–2010. [Google Scholar] [CrossRef] [PubMed]
- Stahlbom, A.K.; Sara, V.R.; Hoeben, P. Insulin-like growth factor mRNA in Barramundi (Lates calcarifer): Alternative splicing and nonresponsiveness to growth hormone. Biochem. Genet. 1999, 37, 69–93. [Google Scholar] [CrossRef]
- Won, E.T.; Douros, J.D.; Hurt, D.A.; Borski, R.J. Leptin stimulates hepatic growth hormone receptor and insulin-like growth factor gene expression in a teleost fish, the hybrid striped bass. Gen. Comp. Endocrinol. 2016, 229, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Azizi, S.; Nematollahi, M.A.; Mojazi Amiri, B.; Velez, E.J.; Lutfi, E.; Navarro, I.; Capilla, E.; Gutierrez, J. Lysine and Leucine Deficiencies Affect Myocytes Development and IGF Signaling in Gilthead Sea Bream (Sparus aurata). PLoS ONE 2016, 11, e0147618. [Google Scholar] [CrossRef] [PubMed]
- Nornberg, B.F.; Figueiredo, M.A.; Marins, L.F. Expression profile of IGF paralog genes in liver and muscle of a GH-transgenic zebrafish. Gen. Comp. Endocrinol. 2016, 226, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Shi, X.B.; Chen, J.L. The Temporal and Tissue Expression Patterns of IGF1/2 Gene in Goldfish (Carassius Auratus). J. Nat. Sci. Hunan Norm. Univ. 2017, 40, 44–48. [Google Scholar]
- Yu, Y.; Mu, J.; Fan, Z.; Lei, G.; Yan, M.; Wang, S.; Tang, C.; Wang, Z.; Yu, J.; Zhang, G. Insulin-like growth factor 1 enhances the proliferation and osteogenic differentiation of human periodontal ligament stem cells via ERK and JNK MAPK pathways. Histochem. Cell Biol. 2012, 137, 513–525. [Google Scholar] [CrossRef]
- Pozios, K.C.; Ding, J.; Degger, B.; Upton, Z.; Duan, C. IGFs stimulate zebrafish cell proliferation by activating MAP kinase and PI3-kinase-signaling pathways. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1230–R1239. [Google Scholar] [CrossRef]
- Garrett, S.M.; Hsu, E.; Thomas, J.M.; Pilewski, J.M.; Feghali-Bostwick, C. Insulin-like growth factor (IGF)-II- mediated fibrosis in pathogenic lung conditions. PLoS ONE 2019, 14, e0225422. [Google Scholar] [CrossRef]
- Shen, G.; Wu, R.; Liu, B.; Dong, W.; Tu, Z.; Yang, J.; Xu, Z.; Pan, T. Upstream and downstream mechanisms for the promoting effects of IGF-1 on differentiation of spermatogonia to primary spermatocytes. Life Sci. 2014, 101, 49–55. [Google Scholar] [CrossRef]
- Xue, Y.M.; Li, K.; Lv, J.Q. Inhibition regulation of apoptosis by insulin-like growth factor (IGF)-1. Chin. Bull. Life Sci. 2007, 19, 68–72. [Google Scholar]
- Li, J.; Li, Y. Aquaculture in China--Freshwater pearl culture. World Aquac. 2009, 40, 60. [Google Scholar]
- Li, Y.J.; Sheng, J.Q.; Wang, J.H.; Wang, F.; Dai, Y.G.; Hong, Y.J. Observation on the mantle of the freshwater pearl shell, Hyriopsis Schlegeli. J. Nanchang Univ. (Nat. Sci.) 2009, 33, 279–284. [Google Scholar]
- Jin, Y.L.; Shi, Z.Y.; Li, W.J.; Hao, Y.Y.; Qiang, G. Improvement on mantle cell culture and technique for large nucleated pearl producing in Hyriopsis cumingii. J. Shanghai Ocean. Univ. 2011, 20, 705–711. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wang, G.; Dong, S.; Guo, P.; Cui, X.; Duan, S.; Li, J. Identification of Foxl2 in freshwater mussel Hyriopsis cumingii and its involvement in sex differentiation. Gene 2020, 754, 144853. [Google Scholar] [CrossRef]
- Han, X.J.; Duan, T.T.; Pang, Y.; Xu, Y.D.; Wang, Y.; Yang, Y.Q.; Yi, L.M. Optimization research on the westem blot experimental conditions for detecting phosphorylated protein. Prog. Mod. Biomed. 2017, 17, 4047–4050+4037. [Google Scholar]
- Shavlakadze, T.; Chai, J.; Maley, K.; Cozens, G.; Grounds, G.; Winn, N.; Rosenthal, N.; Grounds, M.D. A growth stimulus is needed for IGF-1 to induce skeletal muscle hypertrophy in vivo. J. Cell Sci. 2010, 123, 960–971. [Google Scholar] [CrossRef]
- El-Shewy, H.M.; Lee, M.H.; Obeid, L.M.; Jaffa, A.A.; Luttrell, L.M. The insulin-like growth factor type 1 and insulin-like growth factor type 2/mannose-6-phosphate receptors independently regulate ERK1/2 activity in HEK293 cells. J. Biol. Chem. 2007, 282, 26150–26157. [Google Scholar] [CrossRef]
- Menetrey, J.; Kasemkijwattana, C.; Day, C.S.; Bosch, P.; Vogt, M.; Fu, F.H.; Moreland, M.S.; Huard, J. Growth factors improve muscle healing in vivo. J. Bone Joint. Surg. Br. 2000, 82, 131–137. [Google Scholar] [CrossRef]
- Florini, J.R.; Ewton, D.Z.; Coolican, S.A. Growth hormone and the insulin-like growth factor system in myogenesis. Endocr. Rev. 1996, 17, 481–517. [Google Scholar] [PubMed]
- Li, W.J.; Li, Q.; Qi, X.X.; Shang, C.; Zhou, Z.R.; Shi, Z.Y. Sequence Clone of cDNA and Gene Expression of Calmodulin in Hyriopsis cumingii. J. Anhui Agric. Sci. 2014, 42, 7310–7314. [Google Scholar]
- Zhang, M.; Xuan, S.; Bouxsein, M.L.; Stechow, D.V.; Akeno, N.; Faugere, M.C.; Malluche, H.; Zhao, G.; Rosen, C.J.; Efstratiadis, A.; et al. Osteoblast-specific knockout of the insulin-like growth factor (IGF) receptor gene reveals an essential role of IGF signaling in bone matrix mineralization. J. Biol. Chem. 2002, 277, 44005–44012. [Google Scholar] [CrossRef] [PubMed]
- Massoner, P.; Ladurner-Rennau, M.; Eder, I.E.; Klocker, H. Insulin-like growth factors and insulin control a multifunctional signalling network of significant importance in cancer. Br. J. Cancer 2010, 103, 1479–1484. [Google Scholar] [CrossRef]
- Bibollet-Bahena, O.; Almazan, G. IGF-1-stimulated protein synthesis in oligodendrocyte progenitors requires PI3K/mTOR/Akt and MEK/ERK pathways. J. Neurochem. 2009, 109, 1440–1451. [Google Scholar] [CrossRef]
- Matsuzaki, H.; Tamatani, M.; Mitsuda, N.; Namikawa, K.; Kiyama, H.; Miyake, S.; Tohyama, M. Activation of Akt kinase inhibits apoptosis and changes in Bcl-2 and Bax expression induced by nitric oxide in primary hippocampal neurons. J. Neurochem. 1999, 73, 2037–2046. [Google Scholar]
- Koroleva, O.A.; Tomlinson, M.; Parinyapong, P.; Sakvarelidze, L.; Leader, D.; Shaw, P.; Doonan, J.H. CycD1, a putative G1 cyclin from Antirrhinum majus, accelerates the cell cycle in cultured tobacco BY-2 cells by enhancing both G1/S entry and progression through S and G2 phases. Plant Cell 2004, 16, 2364–2379. [Google Scholar] [CrossRef]
- Ren, M.; Zhong, X.; Ma, C.Y.; Sun, Y.; Guan, Q.B.; Cui, B.; Guo, J.; Wang, H.; Gao, L.; Zhao, J.J. Insulin-like growth factor-1 promotes cell cycle progression via upregulation of cyclin D1 expression through the phosphatidylinositol 3-kinase/nuclear factor-kappaB signaling pathway in FRTL thyroid cells. Acta. Pharmacol. Sin. 2009, 30, 113–119. [Google Scholar] [CrossRef]
- Lv, T.; Wu, Y.; Mu, C.; Liu, G.; Yan, M.; Xu, X.; Wu, H.; Du, J.; Yu, J.; Mu, J. Insulin-like growth factor 1 promotes the proliferation and committed differentiation of human dental pulp stem cells through MAPK pathways. Arch. Oral Biol. 2016, 72, 116–123. [Google Scholar] [CrossRef]
- Porowinska, D.; Wujak, M.; Roszek, K.; Komoszynski, M. Prokaryotic expression systems. Postepy Hig. Med. Dosw. 2013, 67, 119–129. [Google Scholar] [CrossRef]
- Kaar, W.; Ahrer, K.; Durauer, A.; Greinstetter, S.; Sprinzl, W.; Wechner, P.; Clementschitsch, F.; Bayer, K.; Achmuller, C.; Auer, B.; et al. Refolding of Npro fusion proteins. Biotechnol. Bioeng. 2009, 104, 774–784. [Google Scholar] [PubMed]
- Zhang, Z.C.; Liu, X.H.; Niu, J.Z.; Huang, Y.; Wang, B.; Jian, J.C. Rokaryotic expression and optimization of galectin4 gene from Nile Tilapia (Oreochromis niloticus). J. Guangdong Ocean. Univ. 2020, 40, 118–123. [Google Scholar]
- Cao, C.P.; Chen, X.H.; Wu, J.G.; Shang, A.L.; Yu, H.H.; Kang, W.P.; Zheng, L.L.; Zhang, J.; Ju, T.W. Construction of prokaryotic expression vector for human insulin-like growth factor-1 gene and biological activity of expressed protein. Chin. J. Biol. 2008, 21, 848–851. [Google Scholar]
- Fu, P.; Thompson, J.A.; Leeding, K.S.; Bach, L.A. Insulin-like growth factors induce apoptosis as well as proliferation in LIM 1215 colon cancer cells. J. Cell Biochem. 2007, 100, 58–68. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) | Purpose |
|---|---|---|
| IGF1 | F: TTTGGGTGACAGCCTTAG | partial sequence PCR |
| R: TCCTTCTCGGCAGTGATA | ||
| IGF1-5′ | Outer: ACGGGCAAGGCACAGTAACTCTCCA | 5′RACE |
| Inner: TTTTTCCACACAGGCGAACGCCACA | ||
| IGF1-3′ | Outer: AGTGGTGGCTCTGACGCTGACAGTT | 3′RACE |
| Inner: TGCCTTGCCCGTGGACCAAGTAGAT | ||
| qRT-PCR | F: AGAGTTACTGTGCCTTGCC | qRT-PCR |
| R: ACTTTGTGCTTCCCATCC | ||
| EF1α | F: AGGGTCCTTCAAGTATGCC | Reference gene |
| R: CCAGTTTCCACTCTGCCTA | ||
| IGF1R | F: ATTCCTCAATCGTCGCTCAA | Downstream Genes |
| R: TGCGGAGTTCCAATGCTG | ||
| cyclin D2 | F: GATAGCAGCAGGGAGTGT | |
| R: TTTGTGGATTCCGTTTCG | ||
| MAPK1 | F: AGGTTGGTCCTCGCTACT | |
| R: TGGTGCCCGTATTATGTC | ||
| AKT1 | F: GTGAACAGAAGGCGAAAG | |
| R: ATCCAGGGTCTCAGCATC | ||
| G1 | F: CAAGGTATCACTGCCGAGAA | DsRNA |
| R: TGGATTGCGAGGTGGTTT | ||
| T7-G1 | F: GGATCCTAATACGACTCACTATAGGCAAGGTATCACTGCCGAGAA | |
| R: GGATCCTAATACGACTCACTATAGGTGGATTGCGAGGTGGTTT | ||
| G2 | F: GTTATACGCCTAATAGACTGC | DsRNA |
| R: ACTTTGTGCTTCCCATCC | ||
| T7-G2 | F: GGATCCTAATACGACTCACTATAGGGTTATACGCCTAATAGACTGC | |
| R: GGATCCTAATACGACTCACTATAGGACTTTGTGCTTCCCATCC | ||
| G3 | F: GCCTTCTCAAGAGTTATACGC | DsRNA |
| R: ACTTTGTGCTTCCCATCC | ||
| T7-G3 | F: GGATCCTAATACGACTCACTATAGGGCCTTCTCAAGAGTTATACGC | |
| R: GGATCCTAATACGACTCACTATAGGACTTTGTGCTTCCCATCC | ||
| IGF1-ISH | F: AAAGTCCTCCTCCACAAG | In situ hybridization probe |
| R: GGATCCTAATACGACTCACTATACCACAGGTCGTACAAAGAA | ||
| P-IGF1 | F: TTGTCGACGGAGCTCGAATTCATGATCTCCACCTCACTTATGATTGC | Plasmid construction |
| R: GCTGATATCGGATCCGAATTCTCAGGACCAGACTGGACTTCGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Feng, S.; Wang, H.; Shen, X.; Chen, Y.; Xuan, X.; Fu, Y.; Bai, Z.; Li, W. Identification and Functional Analysis of the Cell Proliferation Regulator, Insulin-like Growth Factor 1 (IGF1) in Freshwater Pearl Mussel (Hyriopsis cumingii). Biology 2022, 11, 1369. https://doi.org/10.3390/biology11091369
Li X, Feng S, Wang H, Shen X, Chen Y, Xuan X, Fu Y, Bai Z, Li W. Identification and Functional Analysis of the Cell Proliferation Regulator, Insulin-like Growth Factor 1 (IGF1) in Freshwater Pearl Mussel (Hyriopsis cumingii). Biology. 2022; 11(9):1369. https://doi.org/10.3390/biology11091369
Chicago/Turabian StyleLi, Xuenan, Shangle Feng, He Wang, Xiaoya Shen, Yige Chen, Xingrong Xuan, Yuanshuai Fu, Zhiyi Bai, and Wenjuan Li. 2022. "Identification and Functional Analysis of the Cell Proliferation Regulator, Insulin-like Growth Factor 1 (IGF1) in Freshwater Pearl Mussel (Hyriopsis cumingii)" Biology 11, no. 9: 1369. https://doi.org/10.3390/biology11091369
APA StyleLi, X., Feng, S., Wang, H., Shen, X., Chen, Y., Xuan, X., Fu, Y., Bai, Z., & Li, W. (2022). Identification and Functional Analysis of the Cell Proliferation Regulator, Insulin-like Growth Factor 1 (IGF1) in Freshwater Pearl Mussel (Hyriopsis cumingii). Biology, 11(9), 1369. https://doi.org/10.3390/biology11091369