


Effects of Climate Change on Chlorophyll a in the Barents Sea: A Long-Term Assessment




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
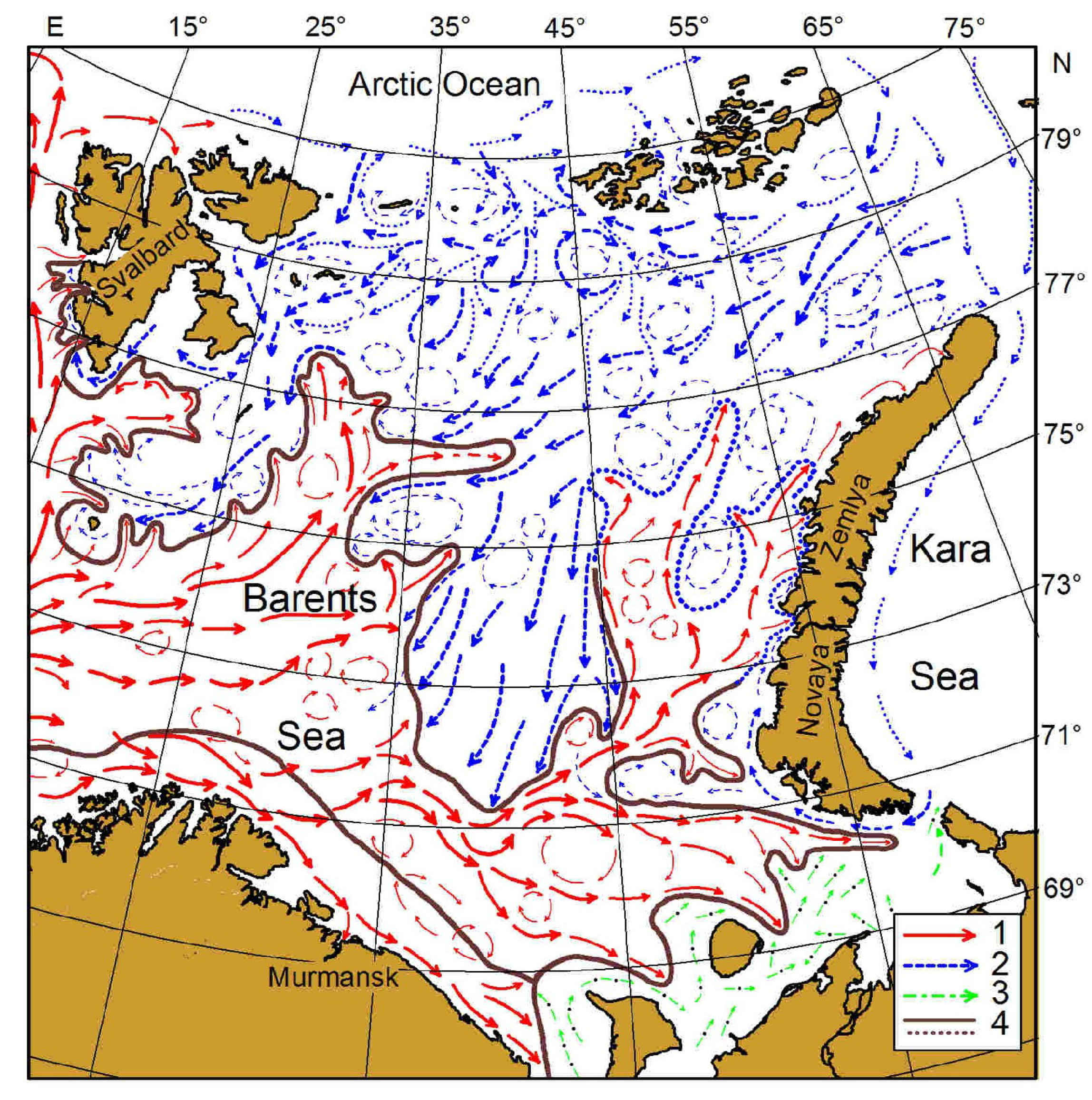
2.1. Sampling
2.2. Environmental and Climatic Indicators
2.3. Statistical Analyses
3. Results
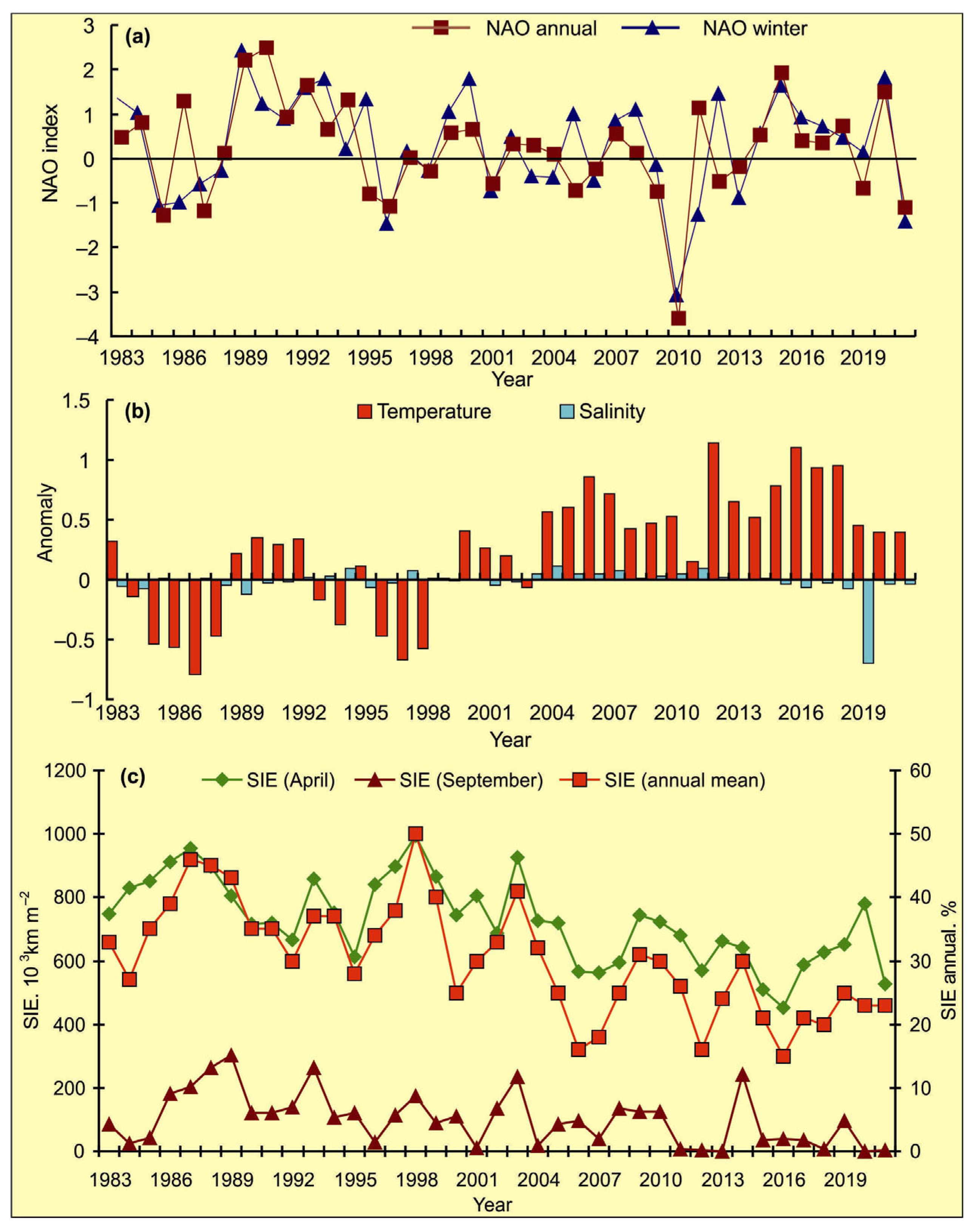
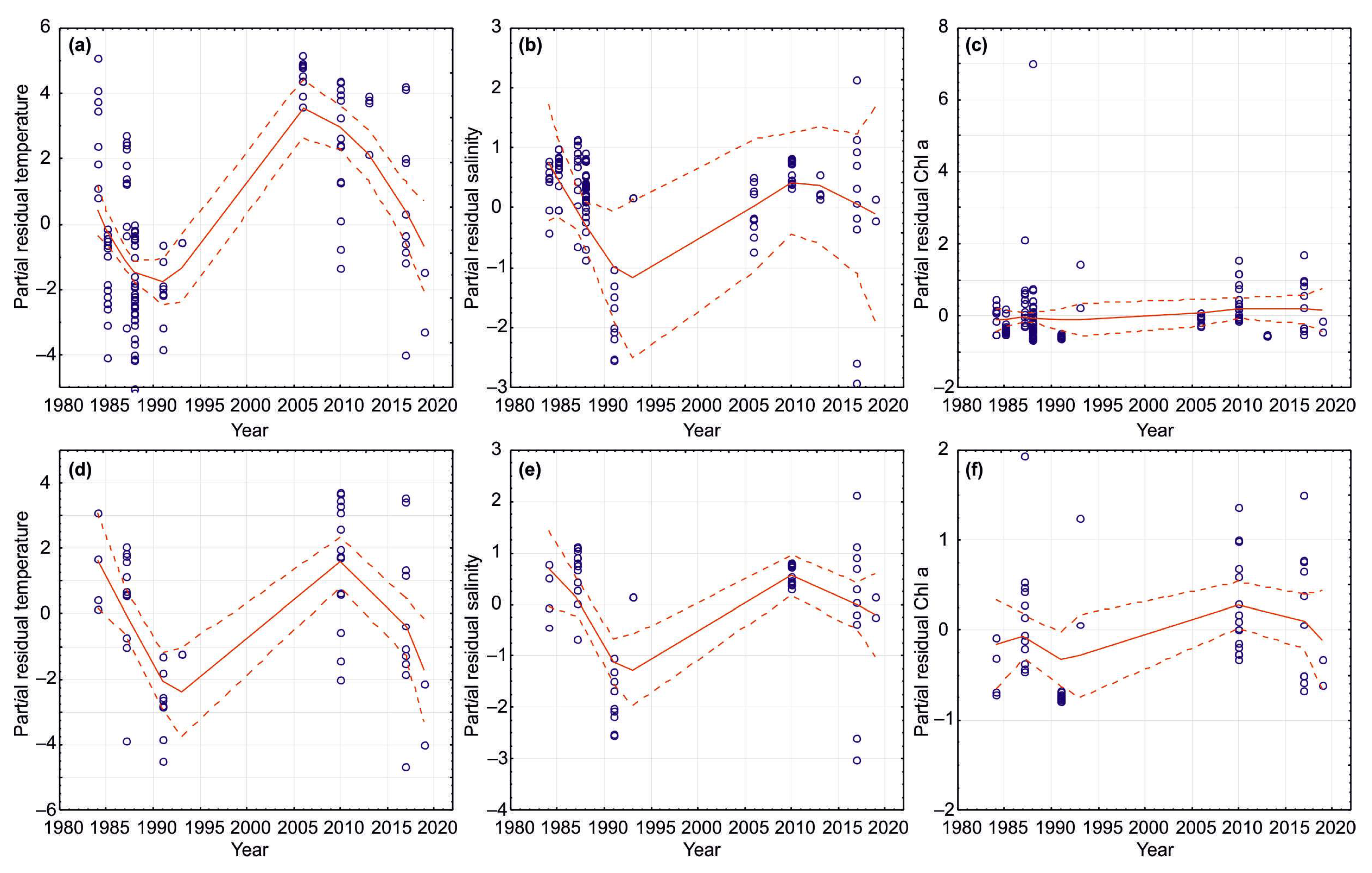
3.1. Temporal Variations in Climatic Indicators
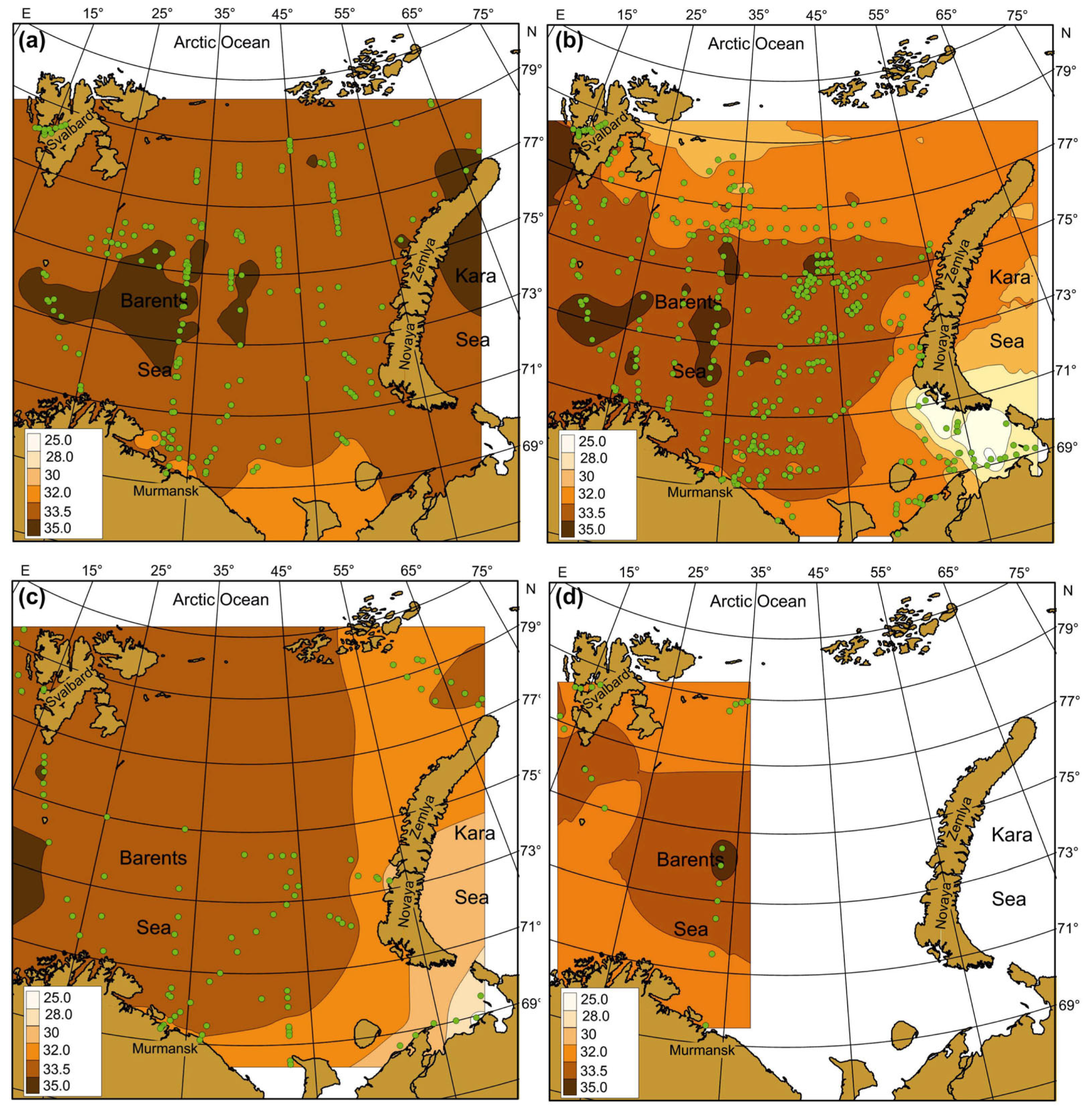
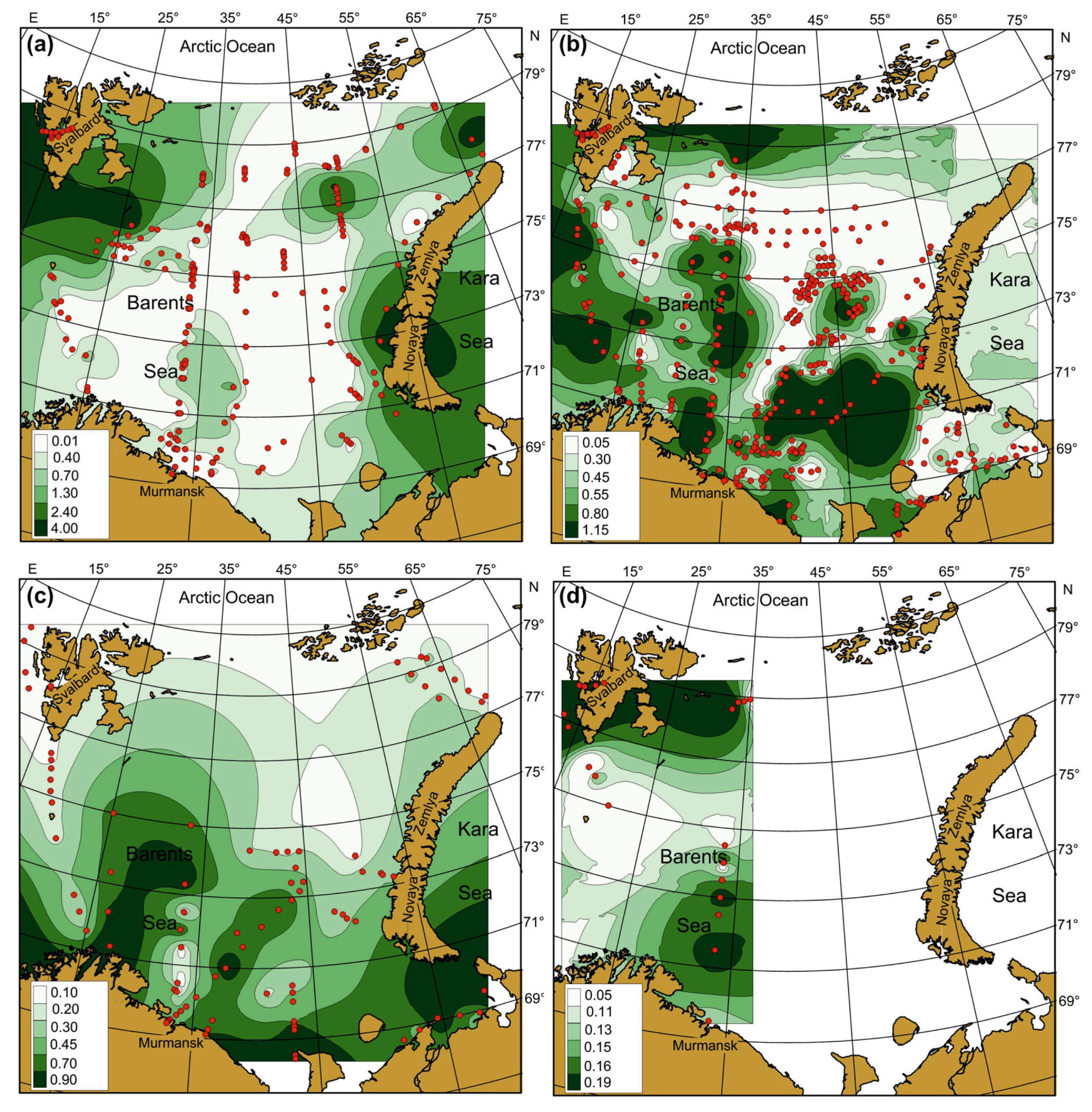
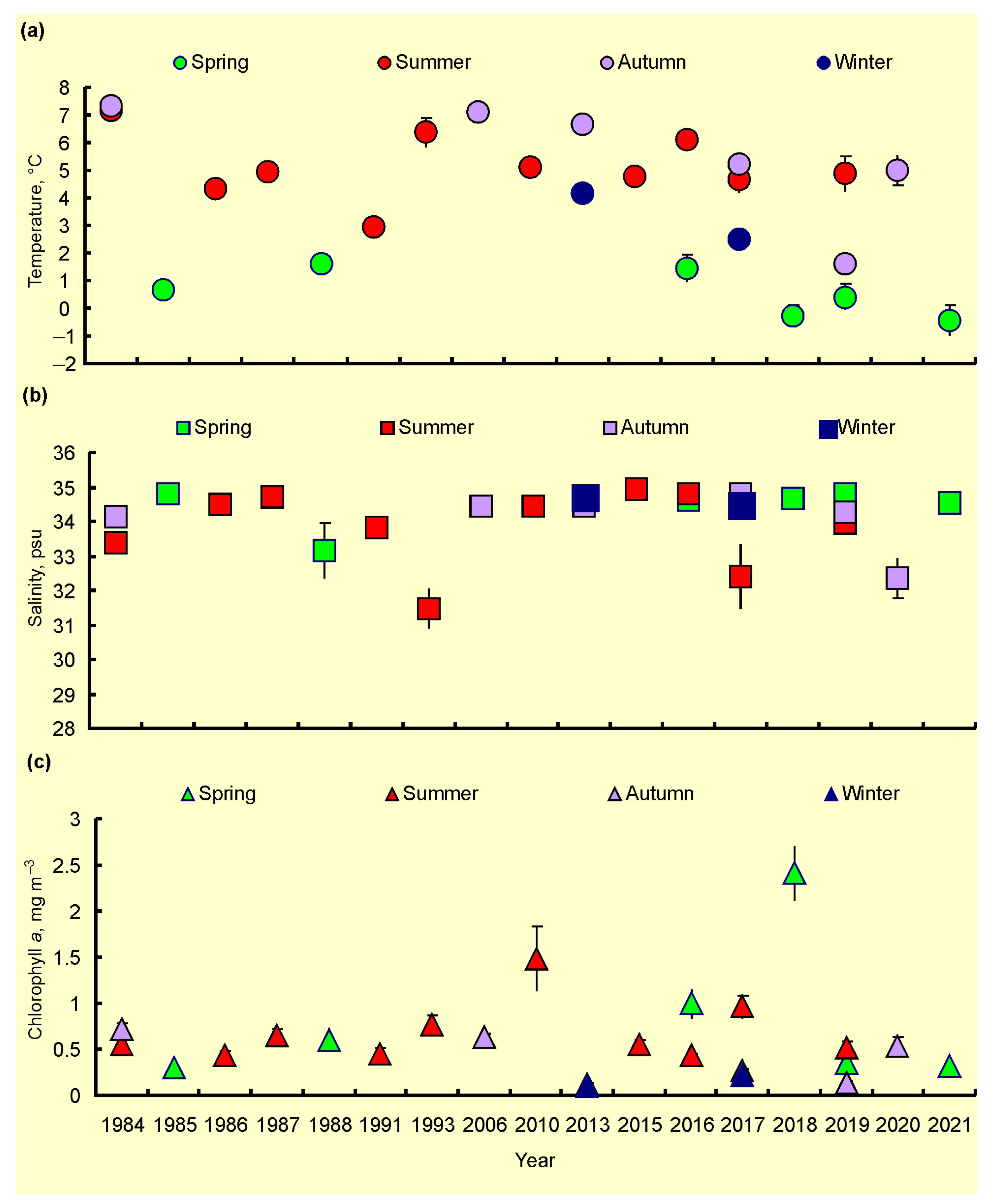
3.2. Spatial, Seasonal, and Temporal Variations of SST, SSS and Chl-a
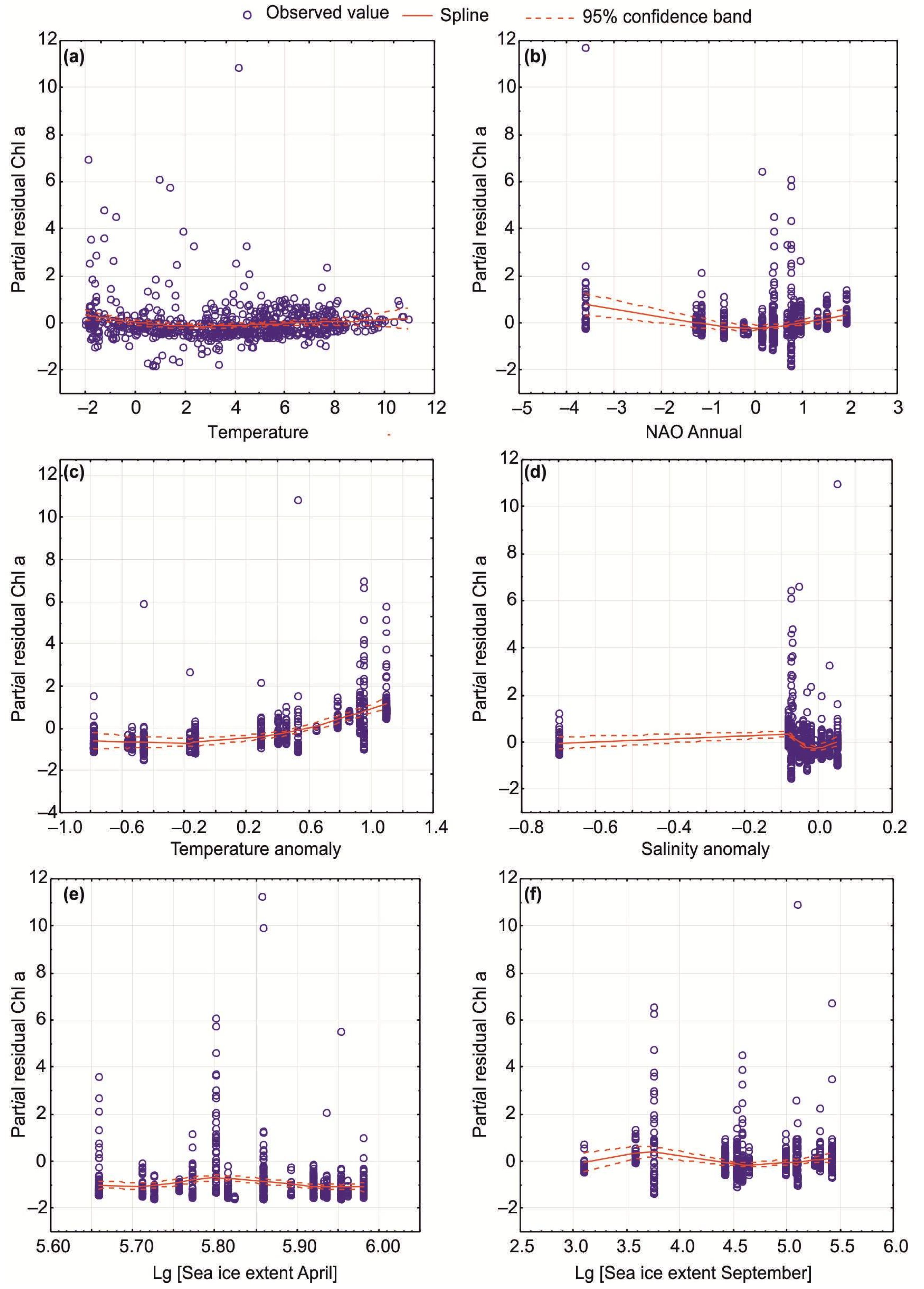
3.3. The Impact of Climatic Factors on Chl-a
4. Discussion
4.1. Temporal Variations in Climatic Indicators
4.2. Spatial, Seasonal, and Temporal Variations in SST, SSS, and Chl-a
4.3. Environmental Impact of Climatic Indicators on SST, SSS, and Chl-a
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhurst, A. Ecological Geography of the Sea; Academic Press: Burlington, ON, Canada, 2007. [Google Scholar]
- ACIA. Arctic Climate Impact Assessment; Cambridge Univ. Press: Cambridge, UK, 2005. [Google Scholar]
- Meier, W.N.; Hovelsrud, G.K.; van Oort, B.E.H.; Key, J.R.; Kovacs, K.M.; Michel, C.; Haas, C.; Granskog, M.A.; Gerland, S.; Perovich, D.K.; et al. Arctic sea ice in transformation: A review of recent observed changes and impacts on biology and human activity. Rev. Geophys. 2014, 51, 185–217. [Google Scholar] [CrossRef]
- Carmack, E.; Polyakov, I.; Padman, L.; Fer, I.; Hunke, E.; Hutchings, J.; Jackson, J.; Kelley, D.; Kwok, R.; Layton, C.; et al. Towards quantifying the increasing role of oceanic heat flux in sea ice loss in the new Arctic. Bull. Am. Meteorol. Soc. 2015, 96, 2079–2105. [Google Scholar] [CrossRef]
- Polyakov, I.V.; Alkire, M.B.; Bluhm, B.A.; Brown, K.A.; Carmack, E.C.; Chierici, M.; Danielson, S.L.; Ellingsen, I.; Ershova, E.A.; Gårdfeldt, K.; et al. Borealization of the Arctic Ocean in response to anomalous advection from sub-arctic seas. Front. Mar. Sci. 2020, 7, 491. [Google Scholar] [CrossRef]
- Polyakov, I.V.; Pnyushkov, A.; Alkire, M.; Ashik, I.M.; Baumann, T.M.; Carmack, E.C.; Goszczko, I.; Guthrie, J.D.; Ivanov, V.V.; Kanzow, T.; et al. Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean. Science 2017, 356, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Oziel, L.; Massicotte, P.; Babin, M.; Devred, E. Decadal changes in Arctic Ocean Chlorophyll a: Bridging ocean color observations from the 1980s to present time. Remote Sens. Environ. 2022, 275, 113020. [Google Scholar] [CrossRef]
- Ardyna, M.; Babin, M.; Gosselin, M.; Devred, E.; Rainville, L.; Tremblay, J.-É. Recent Arctic Ocean sea ice loss triggers novel fall phytoplankton blooms. Geophys. Res. Lett. 2014, 41, 6207–6212. [Google Scholar] [CrossRef]
- Renaut, S.; Devred, E.; Babin, M. Northward expansion and intensification of phytoplankton growth during the early ice-free season in Arctic. Geophys. Res. Lett. 2018, 45, 10590–10598. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, T.; Ozhigin, V.K. (Eds.) The Barents Sea: Ecosystem, Resources, Management: Half a Century of Russian-Norwegian Cooperation; Tapir Academic Press: Trondheim, Norway, 2011. [Google Scholar]
- Matishov, G.G.; Dzhenyuk, S.L.; Denisov, V.V.; Zhichkin, A.P.; Moiseev, D.V. Climate and oceanographic processes in the Barents Sea. Ber. Polarforsch. 2012, 640, 63–73. [Google Scholar]
- Loeng, H.; Drinkwater, K. An overview of the ecosystems of the Barents Sea and Norwegian Seas and their response to climate variability. Deep-Sea Res. II 2007, 54, 2478–2500. [Google Scholar] [CrossRef]
- Sakshaug, E.; Johnsen, G.; Kovacs, K. (Eds.) Ecosystem Barents Sea; Tapir Academic Press: Trondheim, Norway, 2009. [Google Scholar]
- ICES. Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES Sci. Rep. 2021, 3, 1–236. [Google Scholar]
- ICES. Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES Sci. Rep. 2022, 4, 1–235. [Google Scholar]
- Vinje, T. Anomalies and trends of sea ice extent and atmospheric circulation in the Nordic Seas during the period 1864–1998. J. Climatol. 2001, 14, 255–267. [Google Scholar] [CrossRef]
- Loeng, H. Features of the physical oceanographic conditions in the central parts of the Barents Sea. Polar Res. 1991, 10, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Wassmann, P.; Reigstad, M.; Haug, T.; Rudels, B.; Carroll, M.L.; Hop, H.; Gabrielsen, G.W.; Falk-Petersen, S.; Denisenko, S.G.; Arashkevich, E.; et al. Food webs and carbon flux in the Barents Sea. Progr. Oceanogr. 2006, 71, 232–287. [Google Scholar] [CrossRef]
- Lind, S.; Ingvaldsen, R.B.; Furevik, T. Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import. Nat. Clim. Change 2018, 8, 634–639. [Google Scholar] [CrossRef]
- Hunt, G.L.; Blanchard, A.L.; Boveng, P.; Dalpadado, P.; Drinkwater, K.F.; Eisner, L.; Hopcroft, R.R.; Kovacs, K.M.; Norcross, B.L.; Renaud, P.; et al. The Barents and Chukchi Seas: Comparison of two Arctic shelf ecosystems. J. Mar. Syst. 2013, 109–110, 43–68. [Google Scholar] [CrossRef] [Green Version]
- Kahru, M.; Brotas, V.; Manzano-Sarabia, M.; Mitchell, B.G. Are phytoplankton blooms occurring earlier in the Arctic? Glob. Chang. Biol. 2011, 17, 1733–1739. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.L. Continued increases in Arctic Ocean primary production. Prog. Oceanogr. 2015, 136, 60–70. [Google Scholar] [CrossRef]
- Stige, L.C.; Dalpadado, P.; Orlova, E.; Boulay, A.C.; Durant, J.M.; Ottersen, G.; Stenseth, N.C. Spatiotemporal statistical analyses reveal predator–driven zooplankton fluctuations in the Barents Sea. Progr. Oceanogr. 2006, 120, 243–253. [Google Scholar] [CrossRef]
- Dalpadado, P.; Arrigo, K.R.; Hjøllo, S.S.; Rey, F.; Ingvaldsen, R.B.; Sperfeld, E.; van Dijken, G.L.; Stige, L.C.; Olsen, A.; Ottersen, G. Productivity in the Barents Sea-Response to Recent Climate Variability. PLoS ONE 2014, 9, e95273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalpadado, P.; Ingvaldsen, R.; Hassel, A. Zooplankton biomass variation in relation to climatic conditions in the Barents Sea. Polar Biol. 2003, 26, 233–241. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Epiplankton in the Barents Sea: Summer variations of mesozooplankton biomass, community structure and diversity. Cont. Shelf Res. 2013, 52, 1–11. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Coastal mesozooplankton assemblages during spring bloom in the eastern Barents Sea. Biology 2022, 11, 204. [Google Scholar] [CrossRef]
- Aarflot, J.M.; Skjoldal, H.R.; Dalpadado, P.; Skern-Mauritzen, M. Contribution of Calanus species to the mesozooplankton biomass in the Barents Sea. ICES J. Mar. Sci. 2018, 75, 2342–2354. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, E.; Skjoldal, H.R.; Gjøsæter, H.; Primicerio, R. Spatial and temporal changes in the Barents Sea pelagic compartment during the recent warming. Prog. Oceanogr. 2017, 151, 206–226. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Burrows, M.T.; Brown, C.J.; Garcia Molinos, J.; Halpern, B.S.; Hoegh-Guldberg, O.; Kappel, C.V.; Moore, P.J.; Richardson, A.J.; Schoeman, D.S.; et al. Responses of marine organisms to climate change across oceans. Front. Mar. Sci. 2016, 3, 62. [Google Scholar] [CrossRef] [Green Version]
- McClain, C.R. A decade of satellite ocean color observations. Annu. Rev. Mar. Sci. 2009, 1, 19–42. [Google Scholar] [CrossRef] [Green Version]
- Gregg, W.W.; Conkright, M.E. Decadal changes in global ocean chlorophyll. Geophys. Res. Lett. 2002, 29, 20–21. [Google Scholar] [CrossRef] [Green Version]
- Gregg, W.W.; Rousseaux, C.S. Decadal trends in global pelagic ocean chlorophyll: A new assessment integrating multiple satellites, in situ data, and models. J. Geophys. Res. Ocean. 2014, 119, 5921–5933. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Matrai, P.A.; Friedrichs, M.A.M.; Saba, V.S.; Antoine, D.; Ardyna, M.; Asanuma, I.; Babin, M.; Belanger, S.; Benoît-Gagne, M.; et al. An assessment of phytoplankton primary productivity in the Arctic Ocean from satellite ocean color/in situ chlorophyll-a based models. J. Geophys. Res. Ocean. 2015, 120, 6508–6541. [Google Scholar] [CrossRef]
- Kalinka, O.P.; Vodopianova, V.V. Aspects of satellite and subsatellite studies of chlorophyll-a in Arctic waters. IOP Conf. Ser. Earth Environ. Sci. 2022, 981, 042071. [Google Scholar] [CrossRef]
- Lewis, K.M.; Arrigo, K.R. Ocean color algorithms for estimating chlorophyll a, CDOM absorption, and particle backscattering in the Arctic Ocean. J. Geophys. Res. Ocean. 2020, 125, e2019JC015706. [Google Scholar] [CrossRef]
- Matsuoka, A.; Hill, V.; Huot, Y.; Babin, M.; Bricaud, A. Seasonal variability in the light absorption properties of western Arctic waters: Parameterization of the individual components of absorption for ocean color applications. J. Geophys. Res. 2011, 116, C02007. [Google Scholar] [CrossRef] [Green Version]
- Dalpadado, P.; Arrigo, K.R.; van Dijken, G.L.; Skjoldal, H.R.; Bagøien, E.; Dolgov, A.V.; Prokopchuk, I.P.; Sperfeld, E. Climate effects on temporal and spatial dynamics of phytoplankton and zooplankton in the Barents Sea. Progr. Oceanogr. 2020, 182, 102320. [Google Scholar] [CrossRef]
- Makarevich, P.; Druzhkova, E.; Larionov, V. Primary producers of the Barents Sea. In Diversity of Ecosystems; Mahamane, A., Ed.; In Tech: Rijeka, Croatia, 2012; pp. 367–392. [Google Scholar]
- Makarevich, P.R.; Vodopianova, V.V.; Bulavina, A.S. Dynamics of the spatial chlorophyll-a distribution at the Polar Front in the marginal ice zone of the Barents Sea during spring. Water 2022, 14, 101. [Google Scholar] [CrossRef]
- Makarevich, P.R.; Vodopianova, V.V.; Bulavina, A.S.; Vashchenko, P.S.; Ishkulova, T.G. Features of the distribution of chlorophyll-a concentration along the western coast of the Novaya Zemlya archipelago in spring. Water 2021, 13, 3648. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Bulletin of the Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972. [Google Scholar]
- Aminot, A.; Rey, F. Standard Procedure for the Determination of Chlorophyll a by Spectroscopic Methods; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2000. [Google Scholar]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Sea Water Analysis; Pergamon Press: New York, NY, USA, 1992. [Google Scholar]
- Hurrell, J.W.; Kushnir, Y.; Ottersen, G.; Visbeck, M. The North Atlantic Oscillation: Climate Significance and Environmental Impact; American Geophysical Union: Washington, DC, USA, 2003; Volume 134. [Google Scholar]
- Hurrell, J.W.; Deser, C. North Atlantic climate variability: The role of the North Atlantic oscillation. J. Mar. Syst. 2009, 78, 28–41. [Google Scholar] [CrossRef]
- Jones, P.D.; Jónsson, T.; Wheeler, D. Extension to the North Atlantic Oscillation using early instrumental pressure observations from Gibraltar and South-West Iceland. Int. J. Climatol. 1997, 17, 1433–1450. [Google Scholar] [CrossRef]
- Matishov, G.; Zuyev, A.; Golubev, V.; Adrov, N.; Timofeev, S.; Karamusko, O.; Pavlova, L.; Fadyakin, O.; Buzan, A.; Braunstein, A.; et al. Climatic Atlas of the Arctic Seas 2004: Part I. Database of the Barents, Kara, Laptev, and White Seas—Oceanography and Marine Biology; NOAA Atlas NESDIS 58; U.S. Government Printing Office: Washington, DC, USA, 2004. [Google Scholar]
- Ozhigin, V.; Ivshin, V.; Trofimov, A.; Karsakov, A.L.; Antsiferov, M. The Barents Sea Water: Structure, Circulation, Variability; PINRO Press: Murmansk, Russia, 2016. [Google Scholar]
- Norwegian Polar Institute. Sea Ice Extent in the Barents Sea in September. Environmental Monitoring of Svalbard and Jan Mayen (MOSJ). 2022. Available online: http://www.mosj.no/en/climate/ocean/sea-ice-extent-barents-sea-fram-strait.html (accessed on 15 June 2022).
- Hastie, T.; Tibshirani, R. Generalized additive models: Some applications. J. Am. Stat. Assoc. 1987, 82, 371–386. [Google Scholar] [CrossRef]
- National Center for Atmospheric Research (2022) Climate Data Guide. Available online: https://climatedataguide.ucar.edu/climate-data (accessed on 1 July 2022).
- Stroeve, J.C.; Serreze, M.C.; Holland, M.M.; Kay, J.E.; Malanik, J.; Barrett, A.P. The Arctic’s rapidly shrinking sea ice cover: A research synthesis. Clim. Change 2012, 110, 1005–1027. [Google Scholar] [CrossRef] [Green Version]
- Polyakov, I.V.; Bhatt, U.S.; Walsh, J.E.; Abrahamsen, E.P.; Pnyushkov, A.V.; Wassmann, P.F. Recent oceanic changes in the Arctic in the context of long-term observations. Ecol. Appl. 2013, 23, 1745–1764. [Google Scholar] [CrossRef] [Green Version]
- Alekseev, G.V.; Glok, N.I.; Vyazilova, A.E.; Kharlanenkova, N.E.; Kulakov, M.Y. Influence of SST in Low Latitudes on the Arctic Warming and Sea Ice. J. Mar. Sci. Eng. 2021, 9, 1145. [Google Scholar] [CrossRef]
- Yashayaev, I.; Seidov, D. The role of the Atlantic Water in multidecadal ocean variability in the Nordic and Barents Seas. Progr. Oceanogr. 2015, 132, 68–127. [Google Scholar] [CrossRef]
- Stroeve, J.; Notz, D. Changing state of Arctic sea ice across all seasons. Environ. Res. Lett. 2018, 13, 103001. [Google Scholar] [CrossRef]
- Engelsen, O.; Hegseth, E.; Hop, H.; Hansen, E.; Falk-Petersen, S. Spatial variability of chlorophyll-a in the marginal ice zone of the Barents Sea, with relations to sea ice and oceanographic conditions. J. Mar. Syst. 2002, 35, 79–97. [Google Scholar] [CrossRef]
- Wassmann, P.; Ratkova, T.; Andreassen, I.; Vernet, M.; Pedersen, G.; Rey, F. Spring bloom development in the Marginal Ice Zone and the Central Barents Sea. Mar. Ecol. 1999, 20, 321–346. [Google Scholar] [CrossRef] [Green Version]
- Degerlund, M.; Eilertsen, H.C. Main Species Characteristics of Phytoplankton Spring Blooms in NE Atlantic and Arctic Waters (68–80 N). Estuar. Coast. 2010, 33, 242–269. [Google Scholar] [CrossRef]
- Raymont, J.E.G. Plankton and Productivity in the Oceans, 2nd ed.; Phytoplankton; Pergamon Press: New York, NY, USA, 1980; Volume 1. [Google Scholar]
- Lewis, K.M.; Van Dijken, G.L.; Arrigo, K.R. Changes in phytoplankton concentration now drive increased Arctic Ocean primary production. Science 2020, 369, 198–202. [Google Scholar] [CrossRef]
- Skagseth, Ø.; Furevik, T.; Ingvaldsen, R.; Loeng, H.; Mork, K.; Orvik, K.; Ozhigin, V. Volume and heat transports to the Arctic Ocean via the Norwegian and Barents Seas. In Arctic-Subarctic Ocean Fluxes; Dickson, R., Meincke, J., Rhines, P., Eds.; Springer: Dordrecht, Netherlands, 2008; pp. 45–64. [Google Scholar]
- Ingvaldsen, R.B. Width of the North Cape Current and location of the Polar Front in the western Barents Sea. Geophys. Res. Lett. 2005, 32, L16603. [Google Scholar] [CrossRef]
- Visbeck, M.; Chassignet, E.P.; Curry, R.; Delworth, T.; Dickson, B.; Krahman, G. The Ocean’s Response to North Atlantic Oscillation Variability. In The North Atlantic Oscillation: Climate Significance and Environmental Impact; Hurrell, J.W., Kushnir, Y., Ottersen, G., Visbeck, M., Eds.; Geophysical Monograph Series, No. 134; Geophysical Monograph-American Geophysical Union: Washington, DC, USA, 2003; pp. 113–146. [Google Scholar]
- McGinty, N.; Guðmundsson, K.; Ágústsdóttir, K.; Marteinsdóttir, G. Environmental and climactic effects of chlorophyll-a variability around Iceland using reconstructed satellite data fields. J. Mar. Syst. 2016, 163, 31–42. [Google Scholar] [CrossRef]
- Arrigo, K.R. Sea ice ecosystems. Ann. Rev. Mar. Sci. 2014, 6, 439–467. [Google Scholar] [CrossRef] [PubMed]
- Ardyna, M.; Arrigo, K. Phytoplankton dynamics in a changing Arctic Ocean. Nat. Clim. Change 2020, 10, 892–903. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Pavlov, V.; Berge, J.; Cottier, F.; Kovacs, K.M.; Lydersen, C. At the rainbow’s end: High productivity fueled by winter upwelling along an Arctic shelf. Polar Biol. 2015, 38, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Wassmann, P.; Reigstad, M. Future Arctic Ocean seasonal ice zones and implications for pelagic–benthic coupling. Oceanography 2011, 24, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Oziel, L.; Neukermans, G.; Ardyna, M.; Lancelot, C.; Tison, J.-L.; Wassmann, P.; Sirven, J.; Ruiz-Pino, D.; Gascard, J.-C. Role for Atlantic inflows and sea ice loss on shifting phytoplankton blooms in the Barents Sea. J. Geophys. Res. Oceans 2017, 122, 5121–5139. [Google Scholar] [CrossRef] [Green Version]
- Qu, B.; Gabric, A.J. The multi-year comparisons of chlorophyll and sea ice in Greenland Sea and Barents Sea and their relationships with the North Atlantic Oscillation. J. Mar. Syst. 2022, 231, 103749. [Google Scholar] [CrossRef]
- Dong, K.; Kvile, K.Ø.; Stenseth, N.C.; Stige, L.C. Associations among temperature, sea ice and phytoplankton bloom dynamics in the Barents Sea. Mar. Ecol. Prog. Ser. 2020, 635, 25–36. [Google Scholar] [CrossRef]
- Sorteberg, A.; Kvingedal, B. Atmospheric forcing on the Barents sea winter ice extent. J. Clim. 2006, 19, 4772–4784. [Google Scholar] [CrossRef]
- Ding, Q.; Schweiger, A.; L’Heureux, M.; Battisti, D.; Po-Chedley, S.; Johnson, N.; Blanchard-Wrigglesworth, E.; Harnos, K.; Zhang, Q.; Eastman, R.; et al. Influence of high-latitude atmospheric circulation changes on summertime Arctic sea ice. Nat. Clim. Chang. 2017, 7, 289. [Google Scholar] [CrossRef]
- Bélanger, S.; Babin, M.; Tremblay, J.É. Increasing cloudiness in Arctic damps the increase in phytoplankton primary production due to sea ice receding. Biogeosciences 2013, 10, 4087–4101. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer-fall macrozooplankton assemblages in a large Arctic estuarine zone (south-eastern Barents Sea): Environmental drivers of spatial distribution. Mar. Environ. Res. 2022, 173, 105498. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Arctic marine mesozooplankton at the beginning of the polar night: A case study for southern and south-western Svalbard waters. Polar Biol. 2020, 43, 71–79. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Mesozooplankton in the Kola Transect (Barents Sea): Autumn and winter structure. J. Sea Res. 2018, 142, 18–22. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Winter zooplankton in a small Arctic lake: Abundance and vertical distribution. Water 2021, 13, 912. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Early winter mesozooplankton of the coastal south-eastern Barents Sea. Estuar. Coast. Shelf Sci. 2015, 152, 116–123. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Copepod assemblages in a large Arctic coastal area: A baseline summer study. Diversity 2023, 15, 81. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer macrozooplankton assemblages of Arctic shelf: A latitudinal study. Cont. Shelf Res. 2019, 188, 103967. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cruise | Period | Researh Vessel | Number of Samples |
|---|---|---|---|
| 1 | August–September 1984 | Pomor | 46 |
| 2 | August–September 1984 | Dalnie Zelentsy | 29 |
| 3 | August 1984 | Akhill | 24 |
| 4 | April 1985 | Pomor | 29 |
| 5 | June 1986 | Dalnie Zelentsy | 17 |
| 6 | June–July 1987 | Dalnie Zelentsy | 48 |
| 7 | March–May1988 | Dalnie Zelentsy | 28 |
| 8 | March–May1988 | Coastal station | 28 |
| 9 | June–July 1991 | Dalnie Zelentsy | 61 |
| 10 | June 1993 | Dalnie Zelentsy | 33 |
| 11 | June 1993 | Dalnie Zelentsy | 16 |
| 12 | July 1993 | Dalnie Zelentsy | 23 |
| 13 | September–October 2006 | Dalnie Zelentsy | 18 |
| 14 | August 2010 | Dalnie Zelentsy | 34 |
| 15 | November 2013 | Dalnie Zelentsy | 10 |
| 16 | June–July 2015 | Dalnie Zelentsy | 52 |
| 17 | April–May 2016 | Dalnie Zelentsy | 51 |
| 18 | June–July 2016 | Dalnie Zelentsy | 52 |
| 19 | July 2017 | Dalnie Zelentsy | 30 |
| 20 | November–December 2017 | Dalnie Zelentsy | 27 |
| 21 | April–May 2018 | Dalnie Zelentsy | 46 |
| 22 | April 2019 | Dalnie Zelentsy | 30 |
| 23 | June 2019 | Dalnie Zelentsy | 12 |
| 24 | November 2019 | Dalnie Zelentsy | 6 |
| 25 | September 2020 | Dalnie Zelentsy | 6 |
| 26 | October 2020 | Dalnie Zelentsy | 9 |
| 27 | March–April 2021 | Dalnie Zelentsy | 38 |
| Total | 803 |
| Parameter | Season | |||
|---|---|---|---|---|
| Spring (A) | Summer (B) | Autumn (C) | Winter (D) | |
| Temperature | ||||
| min–max | −1.7–10.9 | −2–+7 | −0.8–+12.2 | −1.8–+7.3 |
| mean ± SE | 0.7 ± 0.1 BCD | 5.2 ± 0.1 AD | 6.2 ± 0.3 AD | 3.0 ± 0.6 ABC |
| Salinity | ||||
| min–max | 27.55–35.04 | 8.25–35.3 | 6.16–35.22 | 34.07–35.07 |
| mean ± SE | 34.32 ± 0.18 | 33.85 ± 0.11 | 34.01 ± 0.13 | 34.55 ± 0.07 |
| Chlorophyll a | ||||
| min–max | 0.08–1.65 | 0.01–8.25 | 0.03–12.36 | 0.05–0.28 |
| mean ± SE | 0.9 ± 0.09 D | 0.66 ± 0.04 D | 0.55 ± 0.04 | 0.18 ± 0.02 AB |
| Season | Slope | Intercept | F | p | R2 |
|---|---|---|---|---|---|
| SST | |||||
| Summer | 0.022 ± 0.025 | –39 ± 50 | 0.75 | 0.390 | 0.014 |
| All | 0.106 ± 0.018 | –209 ± 37 | 33.87 | <0.001 | 0.208 |
| SSS | |||||
| Summer | 0.013 ± 0.012 | 8 ± 25 | 1.10 | 0.230 | 0.020 |
| All | 0.027 ± 0.030 | –19 ± 60 | 0.76 | 0.385 | 0.006 |
| Chl-a | |||||
| Summer | 0.012 ± 0.007 | –23 ± 14 | 2.89 | 0.095 | 0.053 |
| All | 0.010 ± 0.006 | –19 ± 12 | 2.56 | 0.112 | 0.019 |
| Season | GAM Coefficient (β) | Standard Error | Standard Score | R Square × 100% | Non-Linear p-Value | |
|---|---|---|---|---|---|---|
| SST | ||||||
| Summer | Normal | 0.022 | 0.019 | 1.11 | 43.6 | <0.001 |
| Poisson | 0.056 | 0.022 | 2.57 | 45.3 | <0.001 | |
| All | Normal | 0.106 | 0.014 | 7.33 | 52.7 | <0.001 |
| Poisson | 0.103 | 0.016 | 6.24 | 49.8 | <0.001 | |
| SSS | ||||||
| Summer | Normal | 0.013 | 0.010 | 1.30 | 40.4 | <0.001 |
| Poisson | 0.017 | 0.000 | 42.95 | 40.1 | 0.935 | |
| All | Normal | 0.006 | 0.018 | 0.33 | 4.9 | 0.178 |
| Poisson | 0.006 | 0.044 | 0.14 | 3.6 | 0.897 | |
| Chl-a | ||||||
| Summer | Normal | 0.012 | 0.007 | 1.77 | 17.4 | 0.124 |
| Poisson | 0.015 | 0.009 | 1.65 | 27.8 | 0.144 | |
| All | Normal | 0.009 | 0.006 | 1.52 | 3.0 | 0.810 |
| Poisson | 0.009 | 0.007 | 1.42 | 5.5 | 0.773 | |
| Added Function | Final Deviance | Residual Df | Number of Observations | Outer Iteration Number | Number of Smooths | Scale Estimate | R Square × 100% |
|---|---|---|---|---|---|---|---|
| All seasons | |||||||
| Normal | 547.9 | 716.0 | 753 | 1 | 15 | 0.765 | 25.8 |
| Poisson | 365.4 | 715.4 | 753 | 18 | 243 | 1.000 | 34.5 |
| Spring | |||||||
| Normal | 296.8 | 212.0 | 249 | 1 | 15 | 1.400 | 35.4 |
| Poisson | 165.0 | 211.5 | 249 | 20 | 300 | 1.000 | 47.2 |
| Summer | |||||||
| Normal | 200.4 | 358.0 | 395 | 1 | 15 | 0.560 | 19.0 |
| Poisson | 126.2 | 357.4 | 395 | 20 | 300 | 1.000 | 35.0 |
| Autumn | |||||||
| Normal | 6.4 | 55.8 | 92 | 1 | 15 | 0.115 | 48.4 |
| Poisson | 9.3 | 55.7 | 92 | 20 | 300 | 1.000 | 57.3 |
| Normal | Poisson | |||||||
|---|---|---|---|---|---|---|---|---|
| Climatic Predictor | GAM Coefficient (β) | Standard Error | Standard Score | Non-Linear p-Value | GAM Coefficient (β) | Standard Error | Standard Score | Non-Linear p-Value |
| All seasons | ||||||||
| SST | 0.012 | 0.012 | −1.018 | <0.05 | 0.010 | 0.011 | 0.925 | 0.033 |
| SSS | 0.024 | 0.016 | 1.522 | 0.838 | 0.020 | 0.008 | 2.578 | 0.474 |
| NAOAn | −0.052 | 0.056 | −0.916 | <0.001 | 0.034 | 0.045 | 0.762 | <0.001 |
| NAOW | 0.056 | 0.057 | 0.984 | 0.911 | −0.027 | 0.047 | −0.586 | 0.646 |
| dT | 1.020 | 0.156 | 6.546 | <0.001 | 1.253 | 0.140 | 8.922 | <0.001 |
| dS | −0.130 | 0.204 | −0.637 | <0.001 | −0.528 | 0.149 | −3.544 | <0.001 |
| Annual SIE | −0.665 | 0.230 | −2.880 | <0.001 | −0.564 | 0.197 | −2.862 | <0.001 |
| April SIE | −0.380 | 0.336 | 0.926 | <0.001 | −0.318 | 0.270 | −1.170 | 0.783 |
| September SIE | −0.198 | 0.059 | −3.337 | <0.001 | −0.200 | 0.073 | −2.749 | <0.001 |
| Spring | ||||||||
| SST | −0.065 | 0.038 | −1.708 | 0.502 | −0.042 | 0.023 | −1.809 | 0.064 |
| SSS | 0.015 | 0.027 | 0.560 | 0.080 | 0.017 | 0.009 | 1.965 | 0.371 |
| NAOAn | −0.210 | 0.346 | −0.608 | <0.05 | −2.668 | 0.349 | −7.643 | 0.669 |
| NAOW | −0.100 | 0.307 | −0.326 | 0.926 | −2.239 | 0.297 | −7.537 | 0.990 |
| dT | 0.894 | 1.564 | 0.572 | 0.231 | 11.240 | 1.749 | 6.428 | 0.106 |
| dS | 0.978 | 0.738 | 1.326 | <0.001 | 4.758 | 0.751 | 6.337 | <0.001 |
| Annual SIE | −1.933 | 0.437 | −4.417 | <0.05 | −1.343 | 0.323 | −4.153 | <0.05 |
| April SIE | −0.639 | 1.358 | −0.471 | 0.882 | −9.081 | 1.488 | −6.101 | 0.868 |
| September SIE | 0.000 | 0.000 | 0.884 | 0.000 | 0.000 | 0.996 | ||
| Summer | ||||||||
| SST | 0.070 | 0.019 | 3.636 | 0.265 | 0.072 | 0.017 | 4.155 | 0.640 |
| SSS | 0.057 | 0.029 | 2.002 | 0.829 | 0.068 | 0.026 | 2.625 | 0.581 |
| NAOAn | −0.045 | 0.065 | −0.695 | 0.988 | −0.033 | 0.058 | −0.574 | 0.999 |
| NAOW | −0.021 | 0.070 | −0.296 | 0.991 | −0.058 | 0.065 | −0.896 | 0.998 |
| dT | 0.584 | 0.223 | 2.618 | 0.881 | 1.022 | 0.236 | 4.322 | 0.952 |
| dS | 0.880 | 0.324 | 2.717 | <0.05 | 0.448 | 0.305 | 1.469 | <0.001 |
| Annual SIE | −0.039 | 1.150 | −0.034 | 0.650 | 1.022 | 1.038 | 0.984 | 0.999 |
| April SIE | 3.359 | 1.718 | 1.955 | 0.794 | 4.635 | 1.739 | 2.666 | 1.000 |
| September SIE | −0.212 | 0.233 | −0.911 | 0.999 | −0.532 | 0.218 | −2.434 | 0.968 |
| Autumn | ||||||||
| SST | 0.054 | 0.021 | 2.540 | 0.435 | 0.036 | 0.037 | 0.975 | 0.913 |
| SSS | −0.003 | 0.043 | −0.063 | 0.717 | −0.019 | 0.089 | −0.210 | 0.993 |
| NAOAn | 0.200 | 0.399 | 0.503 | 0.239 | −0.049 | 0.641 | −0.077 | 1.000 |
| NAOW | 0.017 | 0.258 | 0.065 | 0.979 | 0.056 | 0.367 | 0.152 | 1.000 |
| dT | 0.129 | 0.159 | 0.810 | 0.812 | −0.428 | 0.296 | −1.445 | 0.988 |
| dS | −0.719 | 0.682 | −1.054 | 0.194 | −0.057 | 1.142 | −0.050 | 1.000 |
| Annual SIE | −1.008 | 1.116 | −0.904 | 0.815 | −2.769 | 1.862 | −1.487 | 1.000 |
| April SIE | 0.249 | 0.326 | 0.764 | 0.718 | 0.820 | 0.616 | 1.332 | 1.000 |
| September SIE | 0.000 | 0.000 | 0.958 | 0.000 | 0.000 | 0.995 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, V.G.; Vodopianova, V.V.; Bulavina, A.S. Effects of Climate Change on Chlorophyll a in the Barents Sea: A Long-Term Assessment. Biology 2023, 12, 119. https://doi.org/10.3390/biology12010119
Dvoretsky VG, Vodopianova VV, Bulavina AS. Effects of Climate Change on Chlorophyll a in the Barents Sea: A Long-Term Assessment. Biology. 2023; 12(1):119. https://doi.org/10.3390/biology12010119
Chicago/Turabian StyleDvoretsky, Vladimir G., Veronika V. Vodopianova, and Aleksandra S. Bulavina. 2023. "Effects of Climate Change on Chlorophyll a in the Barents Sea: A Long-Term Assessment" Biology 12, no. 1: 119. https://doi.org/10.3390/biology12010119
APA StyleDvoretsky, V. G., Vodopianova, V. V., & Bulavina, A. S. (2023). Effects of Climate Change on Chlorophyll a in the Barents Sea: A Long-Term Assessment. Biology, 12(1), 119. https://doi.org/10.3390/biology12010119