


Shallow-Water Bryozoan Communities in a Glacier Fjord of West Svalbard, Norway: Species Composition and Effects of Environmental Factors



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
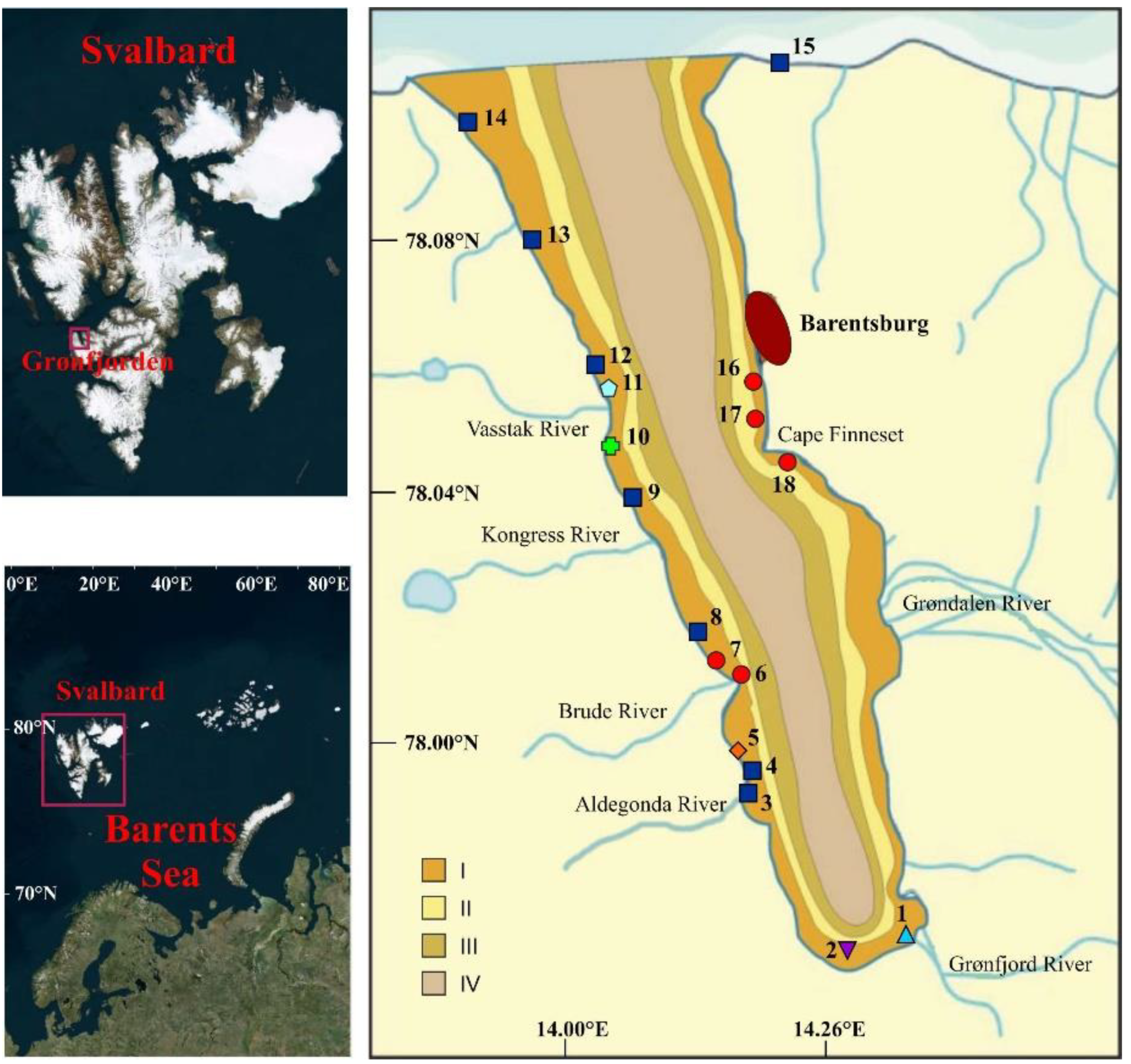
2.1. Study Area
2.2. Sampling and Processing
2.3. Statistical Analysis
3. Results
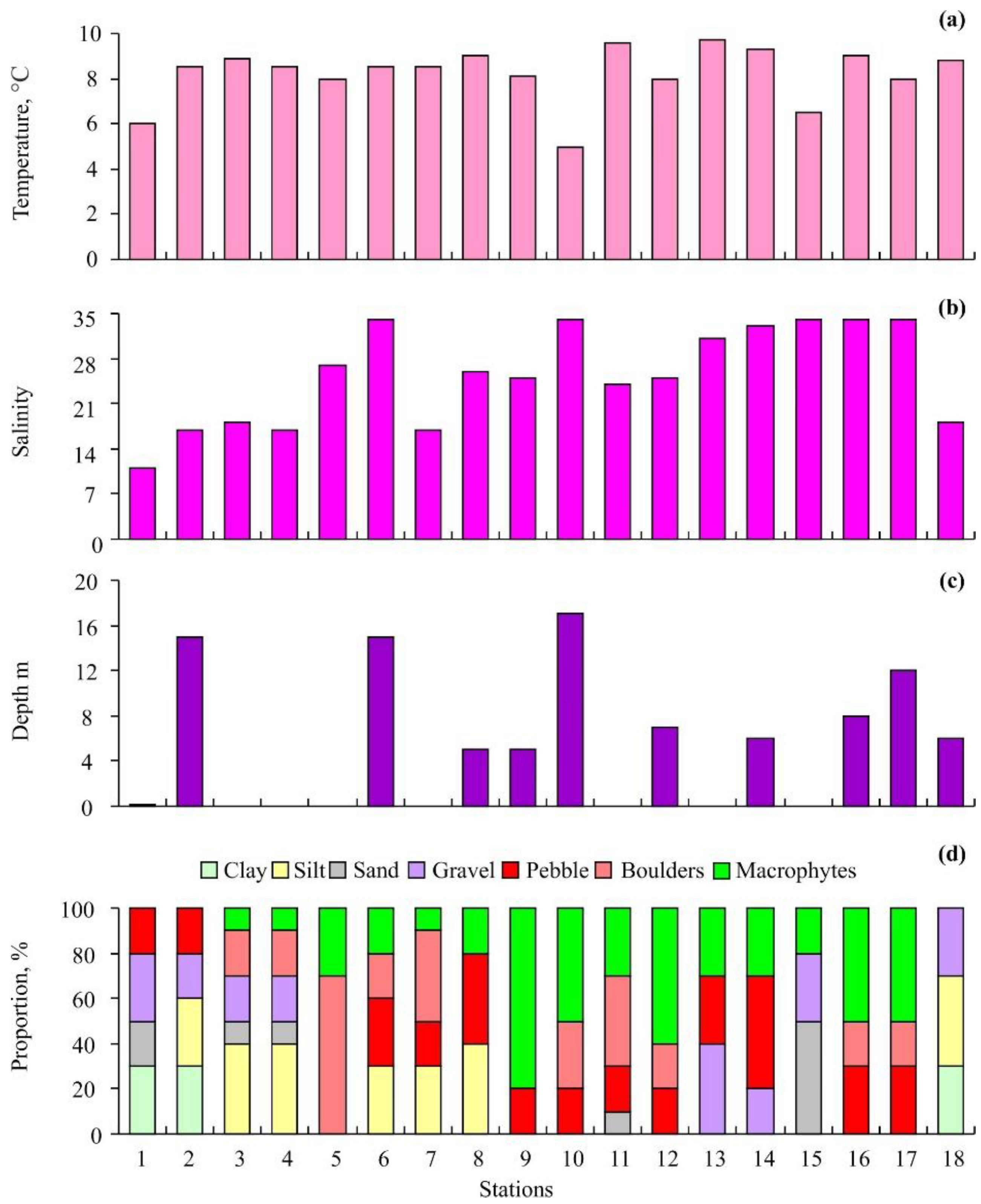
3.1. Environmental Conditions
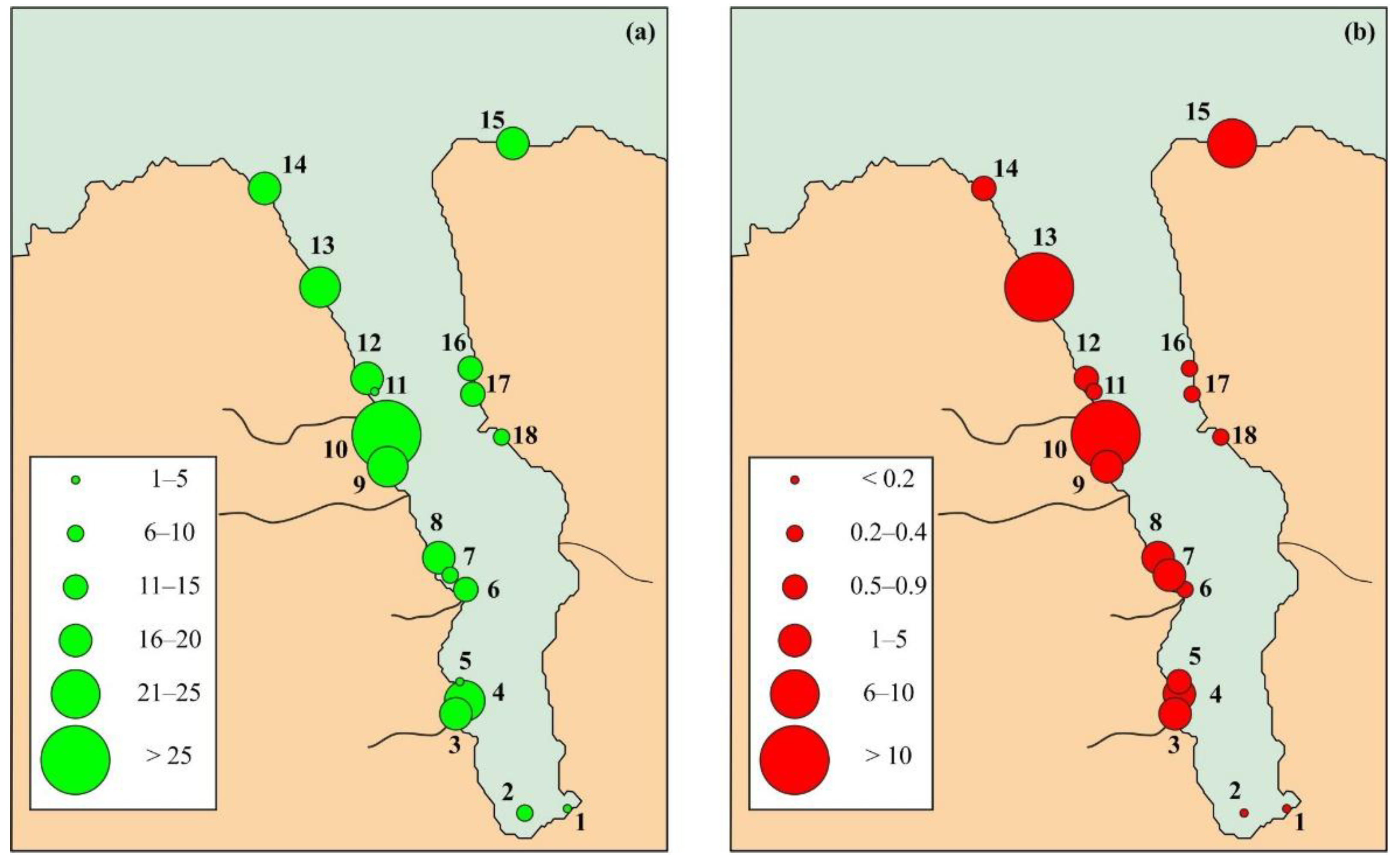
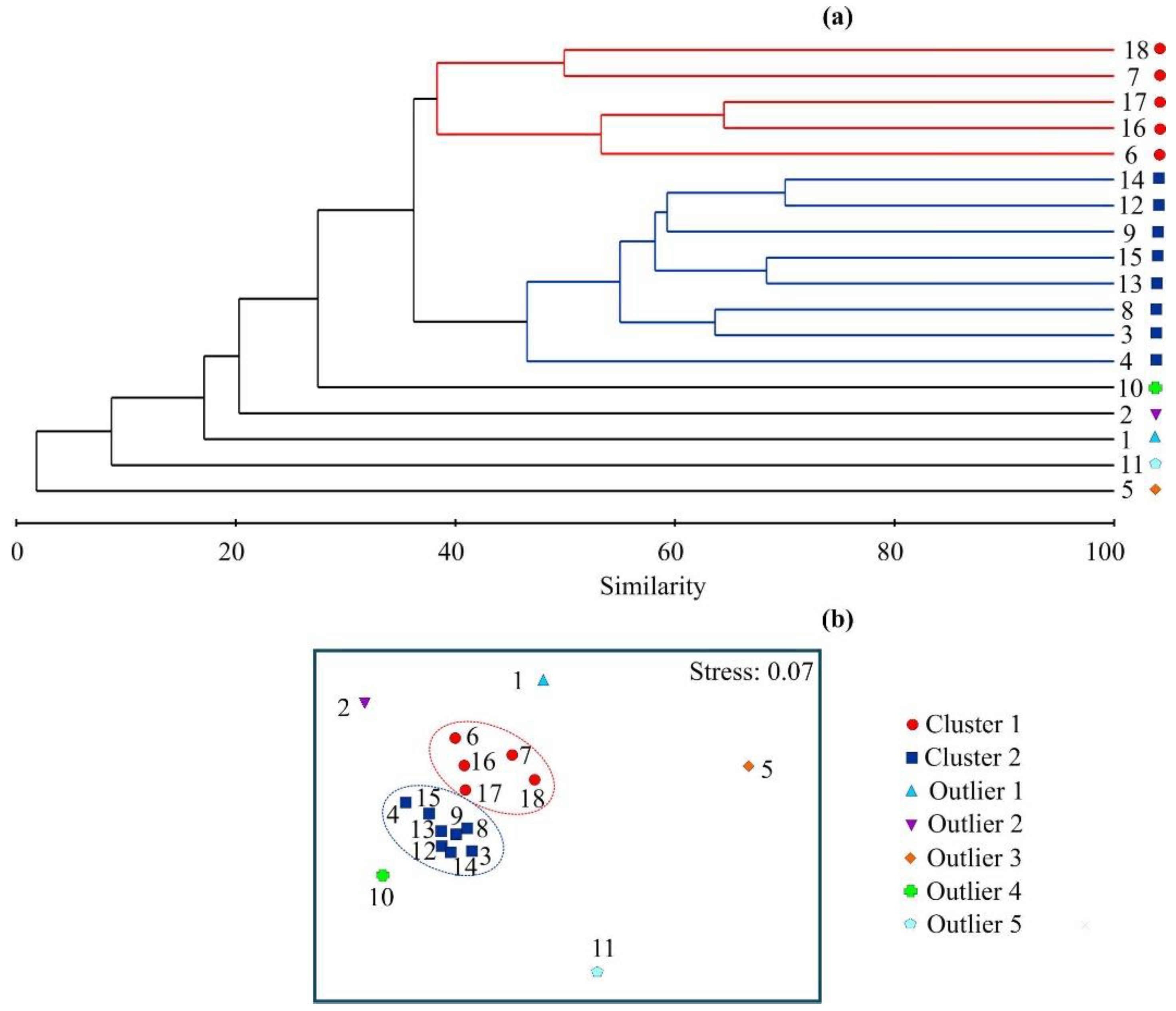
3.2. Bryozoan Diversity, Biomass, and Community
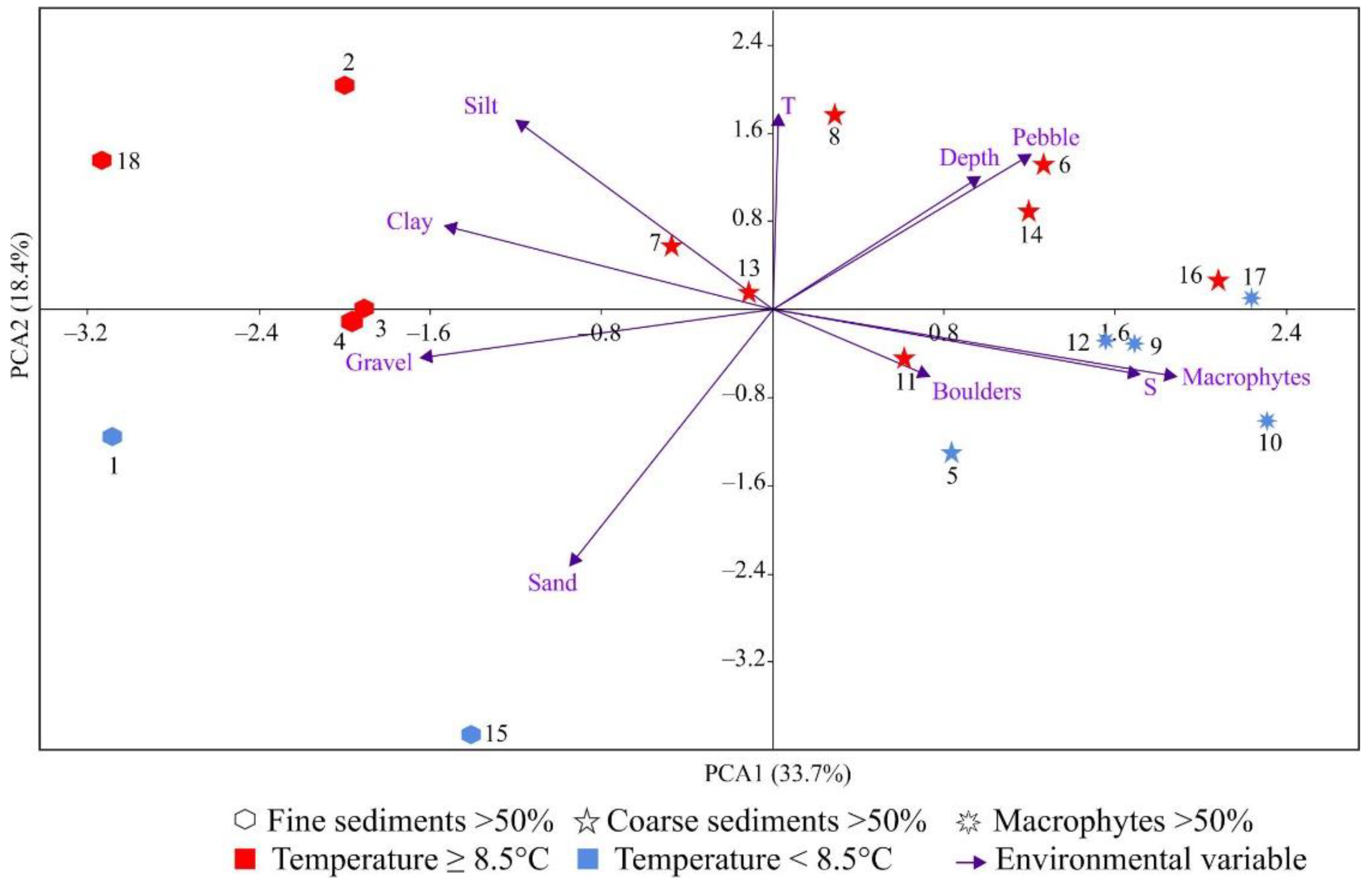
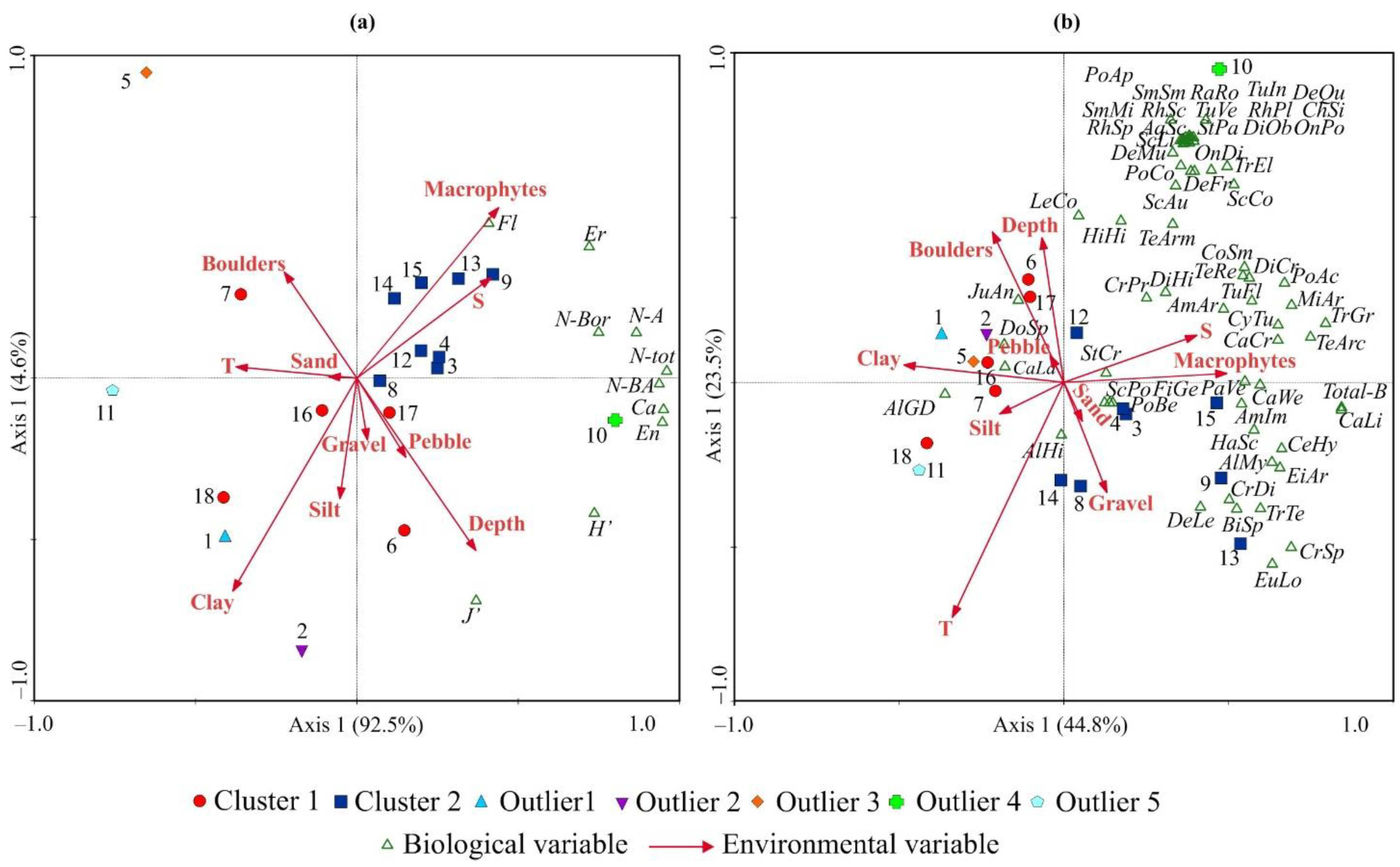
3.3. Relationships between Bryozoan Data and Environmental Variables
4. Discussion
4.1. Environmental Conditions
4.2. Bryozoan Diversity, Biomass, and Communities
4.3. Environmental Control of Bryozoan Communities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mueter, F.J. Arctic fisheries in a changing climate. In Global Arctic; Finger, M., Rekvig, G., Eds.; Springer: Cham, Switzerland, 2022; pp. 279–295. [Google Scholar]
- Jakobsen, T.; Ozhigin, V. (Eds.) The Barents Sea Ecosystem: Russian-Norwegian Cooperation in Science and Management; Tapir Academic Press: Trondheim, Norway, 2011. [Google Scholar]
- Dvoretsky, V.G.; Venger, M.P.; Vashchenko, A.V.; Maksimovskaya, T.M.; Ishkulova, T.G.; Vodopianova, V.V. Pelagic bacteria and viruses in a high Arctic region: Environmental control in the autumn period. Biology 2022, 11, 845. [Google Scholar] [CrossRef] [PubMed]
- Trofimov, A.G. The current trends in oceanographic conditions of the Barents Sea. Tr. Vniro 2021, 186, 101–118. (In Russian) [Google Scholar]
- Dvoretsky, V.G.; Vodopianova, V.V.; Bulavina, A.S. Effects of climate change on chlorophyll a in the Barents Sea: A long-term assessment. Biology 2023, 12, 119. [Google Scholar] [CrossRef] [PubMed]
- Wassmann, P.; Reigstad, M.; Haug, T.; Rudels, B.; Carroll, M.L.; Hop, H.; Gabrielsen, G.W.; Falk-Petersen, S.; Denisenko, S.G.; Arashkevich, E.; et al. Food webs and carbon flux in the Barents Sea. Prog. Oceanogr. 2006, 71, 232–287. [Google Scholar] [CrossRef]
- Hamre, J. Biodiversity and exploitation of the main fish stocks in the Norwegian-Barents Sea ecosystem. Biodiv. Conserv. 1994, 3, 473–492. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Loeng, H.; Drinkwater, K. An overview of the ecosystems of the Barents and Norwegian Seas and their response to climate variability. Deep-Sea Res. II 2007, 54, 2478–2500. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- James, P.; Siikavuopio, S.I.; Mortensen, A. Sea urchin aquaculture in Norway. In Echinoderm Aquaculture; Brown, N.P., Eddy, S.D., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 147–173. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquac. 2020, 12, 1280–1290. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Cucumaria in Russian waters of the Barents Sea: Biological aspects and aquaculture potential. Front. Mar. Sci. 2021, 8, 613453. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Checklist of fauna found in zooplankton samples from the Barents Sea. Polar Biol. 2010, 33, 991–1005. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Coastal mesozooplankton assemblages during spring bloom in the eastern Barents Sea. Biology 2022, 11, 204. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer-fall macrozooplankton assemblages in a large Arctic estuarine zone (south-eastern Barents Sea): Environmental drivers of spatial distribution. Mar. Environ. Res. 2022, 173, 105498. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Copepod assemblages in a large Arctic coastal area: A baseline summer study. Diversity 2023, 15, 81. [Google Scholar] [CrossRef]
- Mecklenburg, C.W.; Møller, P.R.; Steinke, D. Biodiversity of arctic marine fishes: Taxonomy and zoogeography. Mar. Biodiv. 2011, 41, 109–140. [Google Scholar] [CrossRef]
- Sirenko, B.I. List of species of free-living invertebrates of Eurasian Arctic seas and adjacent deep waters. Explor. Fauna Seas 2001, 51, 1–76. [Google Scholar]
- Ryland, J.S. Bryozoa: An introductory overview. Denisia 2005, 19, 9–20. [Google Scholar]
- Gontar, V.I.; Denisenko, N.V. Arctic Ocean Bryozoa. In The Arctic Seas Climatology, Oceanography, Geology, and Biology; Herman, Y., Ed.; Van Nostrand Reinhold: New York, NY, USA, 1989; pp. 341–371. [Google Scholar]
- Kuklinski, P.; Gulliksen, B.; Lønne, O.J.; Weslawski, J.M. Composition of bryozoan assemblages related to depth in Svalbard fjords and sounds. Polar Biol. 2005, 28, 619–630. [Google Scholar] [CrossRef]
- Denisenko, N.V. Species richness and the level of knowledge of the bryozoan fauna of the Arctic region. Proc. Zool. Inst. RAS 2020, 324, 353–363. [Google Scholar] [CrossRef]
- Dvoretsky, A.G. Epibionts of the great spider crab, Hyas araneus (Linnaeus, 1758), in the Barents Sea. Polar Biol. 2012, 35, 625–631. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epifauna associated with the northern stone crab Lithodes maia in the Barents Sea. Polar Biol. 2008, 31, 1149–1152. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Fouling community of the red king crab, Paralithodes camtschaticus (Tilesius 1815), in a subarctic fjord of the Barents Sea. Polar Biol. 2009, 32, 1047–1054. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epifauna associated with an introduced crab in the Barents Sea: A 5-year study. ICES J. Mar. Sci. 2010, 67, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Rossbach, F.I.; Casoli, E.; Plewka, J.; Schmidt, N.; Wild, C. New insights into a Mediterranean Sea benthic habitat: High diversity of epiphytic bryozoan assemblages on Phyllophora crispa (Rhodophyta) Mats. Diversity 2022, 14, 346. [Google Scholar] [CrossRef]
- Bader, B.; Schäfer, P. Bryozoans in polar latitudes: Arctic and Antarctic bryozoan communities and facies. Denisia 2005, 16, 263–282. [Google Scholar]
- Shunatova, N.N.; Ostrovsky, A.N. Individual autozooidal behaviour and feeding in marine bryozoans. Sarsia 2001, 86, 113–142. [Google Scholar] [CrossRef]
- Kluge, G.A. Bryozoans of Northern Seas of USSR; ZIN AN USSR: Leningrad, Russia, 1962. (In Russian) [Google Scholar]
- Kluge, G.A. Bryozoa of the Northern Seas of the USSR; Amerind Publishing Co: New Delhi, India, 1975. [Google Scholar]
- Durrant, H.; Clark, G.F.; Dworjanyn, S.A.; Byrne, M.; Johnston, E.L. Seasonal variation in the effects of ocean warming and acidification on a native bryozoan, Celleporaria nodulosa. Mar. Biol. 2013, 160, 1903–1911. [Google Scholar] [CrossRef]
- Hereu, B.; Garrabou, J.; Montero-Serra, I.; Gori, A.; Gómez-Gras, D.; Figuerola, B.; Linares, C. Divergent responses to warming of two common co-occurring Mediterranean bryozoans. Sci. Rep. 2018, 8, 17455. [Google Scholar]
- O’Dea, A.; Okamura, B. Influence of seasonal variation in temperature, salinity and food availability on module size and colony growth of the estuarine bryozoan Conopeum seurati. Mar. Biol. 1999, 135, 581–588. [Google Scholar] [CrossRef]
- Tishkov, A.A. The ecosystems of the west coast of Spitsbergen (Svalbard archipelago). Polar Geogr. 1985, 9, 70–83. [Google Scholar] [CrossRef]
- Carroll, M.L.; Ambrose, W.G. Benthic in faunal community variability on the northern Svalbard shelf. Polar Biol. 2012, 35, 1259–1272. [Google Scholar] [CrossRef]
- Lippert, H.; Iken, K.; Rachor, E.; Wiencke, C. Macrofauna associated with macroalgae in the Kongsfjord (Spitsbergen). Polar Biol. 2001, 24, 512–522. [Google Scholar]
- Gontar, V.I.; Hop, H.; Voronkov, A.Y. Diversity and distribution of Bryozoa in Kongsfjorden, Svalbard. Polish Polar Res. 2001, 22, 187–204. [Google Scholar]
- Kukliński, P. Fauna of Bryozoa from Kongsfjorden, West Spitsbergen. Polish Polar Res. 2002, 23, 193–206. [Google Scholar]
- Palerud, R.; Gulliksen, B.; Brattegard, T.; Sneli, J.-A.; Vader, W. The marine macro-organisms in Svalbard waters. In A Catalogue of the Terrestrial and Marine Animals of Svalbard; Prestrud, P., Strøm, H., Goldman, H.V., Eds.; Norwegian Polar Institute: Tromsø, Norway, 2004; pp. 5–56. [Google Scholar]
- Kuklinski, P.; Gulliksen, B.; Lønne, O.J.; Weslawski, J.M. Substratum as a structuring influence on assemblages of Arctic bryozoans. Polar Biol. 2006, 29, 652–661. [Google Scholar] [CrossRef]
- Carlsen, B.P.; Johnsen, G.; Berge, J.; Kuklinski, P. Biodiversity patterns of macro-epifauna on different lamina parts of Laminaria digitata and Saccharina latissima collected during spring and summer 2004 in Kongsfjorden, Svalbard. Polar Biol. 2007, 30, 939–943. [Google Scholar] [CrossRef]
- Voronkov, A.; Hop, H.; Gulliksen, B. Zoobenthic communities on hard-bottom habitats in Kongsfjorden, Svalbard. Polar Biol. 2016, 39, 2077–2095. [Google Scholar] [CrossRef]
- Gulliksen, B.; Palerud, R.; Brattegard, T.; Sneil, J. Distribution of Marine Benthic Macroorganisms at Svalbard (Including Bear Island) and Jan Mayen, Research Report for Directorate for Nature Management 1999-4; Trondhjem Biological Station: Trondheim, Norway, 1999. [Google Scholar]
- Kuklinski, P.; Barnes, D.K. Microhabitat diversity of Svalbard Bryozoa. J. Nat. Hist. 2005, 39, 539–554. [Google Scholar] [CrossRef]
- Włodarska-Kowalczuk, M.; Kukliński, P.; Ronowicz, M.; Legeżyńska, J.; Gromisz, S. Assessing species richness of macrofauna associated with macroalgae in Arctic kelp forests (Hornsund, Svalbard). Polar Biol. 2009, 32, 897–905. [Google Scholar] [CrossRef]
- Ronowicz, M.; Włodarska-Kowalczuk, M.; Kukliński, P. Hydroid epifaunal communities in Arctic coastal waters (Svalbard): Effects of substrate characteristics. Polar Biol. 2013, 36, 705–718. [Google Scholar] [CrossRef] [Green Version]
- Meshcheryakov, N.I. Morphometric features and geomorphological appearance of the Gulf of Gren Fjord. In Materials of the XXX Anniversary Conference of Young Scientists of the Murmansk Marine Biological Institute; Matishov, G.G., Ed.; MMBI KSC RAS: Murmansk, Russia, 2012; pp. 126–129. (In Russian) [Google Scholar]
- Meshcheryakov, N.I. Modern Sedimentation in Gren Fjord (West Svalbard). Ph.D. Thesis, MMBI KSC RAS, Murmansk, Russia, 2017. (In Russian). [Google Scholar]
- Antsiferova, A.R.; Mokrotovarova, O.I.; Siekkinen, E.D. Climate change in the Svalbard Archipelago. Climatic features of winter 2013–2014. In Complex Studies of the Nature of Svalbard, Comprehensive Studies of the Nature of Svalbard; Matishov, G.G., Tarasov, G.A., Eds.; GEOS: Moscow, Russia, 2014; pp. 16–21. (In Russian) [Google Scholar]
- Pryamikov, M.V.; Golovanov, O.F.; Tretyakov, M.V.; Nielsen, F. The main results of oceanographic research in the Gulf of Gren Fjord (West Svalbard) as part of the A–162 expedition in 2006–2007. News MPG 2008, 11, 10–15. (In Russian) [Google Scholar]
- Abramoff, M.; Magalhaes, P.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2003, 11, 36–42. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. B 1994, 345, 101–118. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Mescheryakov, N.I.; Bobrov, K.A.; Tarasov, G.A. Sedimenological and oceanological studies in April 2014 in the Gulf of Gren Fjord. In Complex Studies of the Nature of Svalbard; Matishov, G.G., Tarasov, G.A., Eds.; GEOS: Moscow, Russia, 2014; pp. 210–214. (In Russian) [Google Scholar]
- Moiseev, D.V.; Ionov, V.V. Some results of oceanographic studies in the bays and fjords of the island of Western Svalbard in the summer of 2001 and 2002. In Complex Studies of the Nature of Svalbard; Matishov, G.G., Tarasov, G.A., Eds.; KSC RAS: Apatity, Russia, 2006; pp. 261–270. (In Russian) [Google Scholar]
- Moiseev, D.V.; Gromov, M.S. Thermohaline conditions in the bays and fjords of the Western Svalbard Island in the summer of 2003 and 2008. In Complex Studies of the Nature of Svalbard; Matishov, G.G., Tarasov, G.A., Eds.; GEOS: Moscow, Russia, 2009; pp. 332–335. (In Russian) [Google Scholar]
- Berezina, N.A.; Tiunov, A.V.; Petukhov, V.A.; Gubelit, Y.I. Benthic invertebrates abundance and trophic links in the coastal zone during cladophora blooms. Diversity 2022, 14, 1053. [Google Scholar] [CrossRef]
- Rahman, M.K.; Hossain, M.B.; Majumdar, P.R.; Mustafa, M.G.; Noman, M.A.; Albeshr, M.F.; Bhat, E.A.; Arai, T. Macrobenthic assemblages, distribution and functional guilds from a freshwater-dominated tropical estuary. Diversity 2022, 14, 473. [Google Scholar] [CrossRef]
- Pavlova, L.V.; Zuyev, Y.A.; Dvoretsky, A.G. Shallow-water benthic communities on soft bottoms of a sub-arctic fjord (Southern Barents Sea, Russia) along a gradient of ecological factors. Diversity 2023, 15, 84. [Google Scholar] [CrossRef]
- Kukliński, P.; Hayward, P.J. Two new species of cheilostome Bryozoa from Svalbard. Sarsia 2004, 89, 79–84. [Google Scholar] [CrossRef]
- Kuklinski, P.; Taylor, P.D. A new genus and some cryptic species of Arctic and boreal calloporid cheilostome bryozoans. J. Mar. Biol. Ass. 2006, 86, 1035–1046. [Google Scholar] [CrossRef]
- Akhmetchina, O.Y. Formation of the structure of moss animals fauna (Bryozoa) off the Franz Josef Land Archipelago. In Perspective Directions in Investigations of Ecosystems of Russia’s Arctic and Southern Seas: Proceedings of the XXXIV Conference for Young Scientists Dedicated to the 50th Anniversary of the Birth of RAS Correspondent-Member Dmitry G. Matishov; Moiseev, D.V., Ed.; MMBI KSC RAS: Murmansk, Russia, 2017; pp. 9–14. (In Russian) [Google Scholar]
- Ruediger, S.; Fahl, K.; Fütterer, D.K.; Galimov, E.N.; Stepanets, O.V. Siberian River Run-Off in the Kara Sea: Characterisation, Quantification, Variability and Environmental Significance (Vol. 6); Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Gontar, V.I. Bryozoa of the Providence Bay of the Bering Sea. GISAP. Biol. Vet. Med. Agric. Sci. 2013, 2, 30–33. (In Russian) [Google Scholar]
- Matishov, G.G.; Matishov, D.G.; Moiseev, D.V. Inflow of Atlantic-origin waters to the Barents Sea along glacial troughs. Oceanologia 2009, 51, 321–340. [Google Scholar] [CrossRef]
- Matishov, G.; Moiseev, D.; Lyubina, O.; Zhichkin, A.; Dzhenyuk, S.; Karamushko, O.; Frolova, E. Climate and cyclic hydrobiological changes of the Barents Sea from the twentieth to twenty-first centuries. Polar Biol. 2012, 35, 1773–1790. [Google Scholar] [CrossRef]
- Skogseth, R.; Olivier, L.L.; Nilsen, F.; Falck, E.; Fraser, N.; Tverberg, V.; Ledang, A.B.; Vader, A.; Jonassen, M.O.; Søreide, J.; et al. Variability and decadal trends in the Isfjorden (Svalbard) ocean climate and circulation–An indicator for climate change in the European Arctic. Progr. Oceanogr. 2020, 187, 102394. [Google Scholar] [CrossRef]
- Chan, F.T.; Stanislawczyk, K.; Sneekes, A.C.; Dvoretsky, A.; Gollasch, S.; Minchin, D.; David, M.; Jelmert, A.; Albretsen, J.; Bailey, S.A. Climate change opens new frontiers for marine species in the Arctic: Current trends and future invasion risks. Glob. Chang. Biol. 2019, 25, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Akhmetchina, O.Y.; Zakharov, D.V. Species composition and distribution of bryozoans (Bryozoa) along the transect “Kola meridian” (Barents Sea) in 2010–2011. Vestn. MSTU 2013, 16, 415–424. (In Russian) [Google Scholar]
- Evseeva, O.Y. Bryozoa of the northern part of Barents Sea: Species composition, distribution, ecology (based on the materials of expeditions MMBI 2016–2017). Trans. Kola Sci. Cent. 2020, 11, 54–68. (In Russian) [Google Scholar] [CrossRef]
- Evseeva, O.Y.; Ishkulova, T.G.; Dvoretsky, A.G. Environmental drivers of an intertidal bryozoan community in the Barents Sea: A case study. Animals 2022, 12, 552. [Google Scholar] [CrossRef]
- Wlodarska-Kovalczuk, M.; Pearson, T. Soft-bottom faunal associations and factors affecting species distributions in an Arctic glacial fjord (Kongsfjord, Spitsbergen). Polar Biol. 2004, 27, 155–167. [Google Scholar] [CrossRef]
- Kędra, M.; Włodarska-Kowalczuk, M.; Węsławski, J.M. Decadal change in macrobenthic soft-bottom community structure in a high Arctic fjord (Kongsfjorden, Svalbard). Polar Biol. 2010, 33, 1–11. [Google Scholar] [CrossRef]
- Pavlova, L.V.; Dvoretsky, G. Prey selectivity in juvenile red king crabs from the coastal Barents Sea. Diversity 2022, 14, 568. [Google Scholar] [CrossRef]
- Holte, B.; Oug, E.; Cochrane, S. Depth-related benthic macrofaunal biodiversity patterns in three undisturbed north Norwegian fjords. Sarsia 2004, 89, 91–101. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Life cycle of Oithona similis (Copepoda, Cyclopoida) in Kola Bay (Barents Sea). Mar. Biol. 2009, 156, 1433–1446. [Google Scholar] [CrossRef]
- Anderson, E.J.; Peterson, M.S.; Andres, M.J. Drifting bryozoans increase nekton diversity in the north-central Gulf of Mexico unvegetated muddy bottom seascape. Bull. Mar. Sci. 2022, 98, 431–450. [Google Scholar] [CrossRef]
- Balazy, P.; Kuklinski, P.; Włodarska-Kowalczuk, M.; Barnes, D.; Kedra, M.; Legezynska, J.; Wesławski, J.M. Hermit crabs (Pagurus spp.) at their northernmost range: Distribution, abundance and shell use in the European Arctic. Pol. Res. 2015, 34, 21412. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibionts of an introduced king crab in the Barents Sea: A second five-year study. Diversity 2023, 15, 29. [Google Scholar] [CrossRef]
- Davenport, J. Environmental Stress and Behavioural Adaptation; Croom Helm Ltd.: Sydney, Australia, 1985. [Google Scholar]
- Polyakov, I.V.; Alkire, M.B.; Bluhm, B.A.; Brown, K.A.; Carmack, E.C.; Chierici, M.; Danielson, S.L.; Ellingsen, I.; Ershova, E.A.; Gårdfeldt, K.; et al. Borealization of the Arctic Ocean in response to anomalous advection from sub-arctic seas. Front. Mar. Sci. 2020, 7, 491. [Google Scholar] [CrossRef]
- Tesi, T.; Muschitiello, F.; Mollenhauer, G.; Miserocchi, S.; Langone, L.; Ceccarelli, C.; Panieri, G.; Chiggiato, J.; Nogarotto, A.; Hefter, J.; et al. Rapid Atlantification along the Fram Strait at the beginning of the 20th century. Sci. Adv. 2021, 7, eabj2946. [Google Scholar] [CrossRef]
- Morris, R.L.; Graham, T.D.; Kelvin, J.; Ghisalberti, M.; Swearer, S.E. Kelp beds as coastal protection: Wave attenuation of Ecklonia radiata in a shallow coastal bay. Annal. Botany 2020, 125, 235–246. [Google Scholar] [CrossRef]
- Eggleston, D. Factors influencing the distribution of sub-littoral ectoprocts off the south of the Isle of Man (Irish Sea). J. Nat. Hist. 1972, 6, 247–260. [Google Scholar] [CrossRef]
- Seed, R.; O’Connor, R.J. Community organization in marine algal epifaunas. Ann. Rev. Ecol. Syst. 1981, 12, 49–74. [Google Scholar] [CrossRef]
- Harmelin, J.-G. Bryozoan dominated assemblages in Mediterranean cryptic environments. In Bryozoa: Ordovician to Recent; Nielsen, C., Larwood, G.P., Eds.; Olsen & Olsen: Fredensborg, Denmark, 1985; pp. 135–143. [Google Scholar]
- Ward, M.A.; Thorpe, J.P. Assessment of space utilisation in a subtidal temperate bryozoan community. Mar. Biol. 1989, 103, 215–224. [Google Scholar] [CrossRef]
- Fine, M.; Loya, Y. Alternate coral–bryozoan competitive superiority during coral bleaching. Mar. Biol. 2003, 142, 989–996. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Code | X | SE | Min | Max | Or | CF | OF | Stations |
|---|---|---|---|---|---|---|---|---|---|
| Cyclostomatida | |||||||||
| Crisiella diversa (Kluge, 1955) | CrDi | 0.207 | 0.136 | 0.050 | 0.615 | A | Ca | Er | 12, 13, 14, 15 |
| Crisiella producta (Smitt, 1865) | CrPr | 0.936 | 0.902 | 0.003 | 2.740 | BA | Ca | Er | 4, 6, 10 |
| Diplosolen obelium (Johnston, 1838) | DiOb | 0.032 | 0.000 | 0.032 | 0.032 | B | Ca | En | 10 |
| Disporella crassiuscula (Smitt, 1867) | DiCr | 0.007 | 0.002 | 0.005 | 0.009 | A | Ca | En | 10, 13 |
| Disporella hispida (Fleming, 1828) | DiHi | 0.144 | 0.131 | 0.001 | 0.799 | BA | Ca | En | 2, 4, 6, 7, 9, 10 |
| Filicrisia geniculata (Milne Edwards, 1838) | FiGe | 0.002 | 0.000 | 0.002 | 0.002 | B | Ca | Er | 3 |
| Oncousoecia diastoporides (Norman, 1869) | OnDi | 0.024 | 0.021 | 0.003 | 0.045 | BA | Ca | En | 4, 10 |
| Oncousoecia polygonalis (Kluge, 1915) | OnPo | 0.005 | 0.000 | 0.005 | 0.005 | A | Ca | En | 10 |
| Patinella verrucaria (Linnaeus, 1758) | PaVe | 0.007 | 0.002 | 0.001 | 0.028 | BA | Ca | En | 4, 6, 7, 8, 9, 10, 12, 13, 15, 16, 17, 18 |
| Tubulipora flabellaris (O. Fabricius, 1780) | TuFl | 0.012 | 0.006 | 0.001 | 0.033 | BA | Ca | En | 4, 6, 10, 13, 15, 17 |
| Tubulipora ventricosa (Busk, 1855) | TuVe | 0.003 | 0.000 | 0.003 | 0.003 | BA | Ca | Er | 10 |
| Ctenostomatida | |||||||||
| Alcyonidioides mytili (Dalyell, 1848) | AlMy | 0.010 | 0.004 | 0.001 | 0.035 | BA | Fl | En | 3, 8, 9, 10, 13, 14, 17 |
| Alcyonidium gelatinosum diaphanum (Farre, 1837) | AlGD | 0.339 | 0.244 | 0.095 | 0.582 | BA | Fl | Er | 5, 18 |
| Alcyonidium hirsutum (Fleming, 1828) | AlHi | 0.481 | 0.428 | 0.003 | 3.050 | BA | Fl | En | 7, 8, 9, 10, 11, 12, 14 |
| Amathia arctica (Busk, 1880) | AmAr | 0.007 | 0.004 | 0.001 | 0.018 | A | Fl | Er | 3, 9, 10, 12 |
| Amathia imbricata (Adams, 1800) | AmIm | 0.044 | 0.035 | 0.001 | 0.250 | B | Fl | Er | 3, 8, 10, 12, 13, 14, 15 |
| Cheilostomatida | |||||||||
| Aquiloniella scabra (van Beneden, 1848) | AqSc | 0.488 | 0.000 | 0.488 | 0.488 | BA | Ca | Er | 10 |
| Bidenkapia spitzbergensis (Bidenkap, 1897) | BiSp | 0.002 | 0.001 | 0.001 | 0.004 | BA | Ca | En | 8, 13, 15 |
| Callopora craticula (Alder, 1856) | CaCr | 0.008 | 0.003 | 0.002 | 0.017 | BA | Ca | En | 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 |
| Callopora lata (Kluge, 1907) | CaLa | 0.005 | 0.000 | 0.005 | 0.005 | A | Ca | En | 16 |
| Callopora lineata (Linnaeus, 1767) | CaLi | 0.022 | 0.007 | 0.001 | 0.100 | BA | Ca | En | 2, 3, 4, 6, 8, 9, 10, 12, 13, 14, 15, 16, 17 |
| Callopora weslawski (Kuklinski and Taylor, 2006) | CaWe | 0.005 | 0.001 | 0.001 | 0.009 | A | Ca | En | 3, 9, 10, 12, 13, 14, 17 |
| Celleporella hyalina (Linnaeus, 1767) | CeHy | 0.132 | 0.071 | 0.001 | 1.077 | BA | Ca | En | 1, 2, 3, 4, 6, 7, 8, 9, 10, 12, 13, 14, 15, 16, 17 |
| Cheilopora sincera (Smitt, 1868) | ChSi | 0.018 | 0.000 | 0.018 | 0.018 | BA | Ca | En | 10 |
| Copidozoum smitti (Kluge, 1946) | CoSm | 0.029 | 0.021 | 0.003 | 0.090 | A | Ca | En | 3, 8, 9, 10 |
| Cribrilina spitzbergensis (Norman, 1903) | CrSp | 0.079 | 0.032 | 0.001 | 0.332 | A | Ca | En | 3, 4, 8, 9, 12, 13, 14, 15, 17, 18 |
| Cylindroporella tubulosa (Norman, 1868) | CyTu | 0.008 | 0.004 | 0.001 | 0.034 | BA | Ca | En | 3, 4, 8, 9, 10, 13, 16, 17 |
| Dendrobeania fruticosa (Packard, 1863) | DeFr | 0.312 | 0.220 | 0.060 | 0.970 | BA | Ca | Er | 4, 6, 10, 15 |
| Dendrobeania levinseni (Kluge, 1929) | DeLe | 0.288 | 0.000 | 0.288 | 0.288 | BA | Ca | Er | 13 |
| Dendrobeania murrayana (Bean in Johnston, 1847) | DeMu | 0.449 | 0.444 | 0.005 | 0.893 | BA | Ca | Er | 10, 14 |
| Dendrobeania quadridentata (Lovén, 1834) | DeQu | 0.113 | 0.000 | 0.113 | 0.113 | BA | Ca | Er | 10 |
| Doryporella spathulifera (Smitt, 1868) | DoSp | 0.002 | 0.000 | 0.002 | 0.002 | BA | Ca | En | 2 |
| Einhornia arctica (Borg, 1931) | EiAr | 0.027 | 0.008 | 0.001 | 0.078 | BA | Ca | En | 3, 4, 6, 8, 9, 10, 12, 13, 14, 15, 16, 17 |
| Eucratea loricata (Linnaeus, 1758) | EuLo | 1.203 | 0.677 | 0.006 | 7.570 | BA | Ca | Er | 3, 4, 7, 8, 9, 10, 12, 13, 14, 15, 18 |
| Harmeria scutulata (Busk, 1855) | HaSc | 0.086 | 0.056 | 0.001 | 0.862 | A | Ca | En | 1, 3, 4, 6, 7, 8, 9, 10, 12, 13, 14, 15, 16, 17, 18 |
| Hippoporella hippopus (Smitt, 1868) | HiHi | 0.010 | 0.002 | 0.006 | 0.014 | BA | Ca | En | 2, 4, 10 |
| Juxtacribrilina annulata (Fabricius, 1780) | JuAn | 0.003 | 0.001 | 0.001 | 0.006 | BA | Ca | En | 1, 6, 7, 15, 16, 17 |
| Lepraliella contigua (Smitt, 1868) | LeCo | 0.004 | 0.003 | 0.001 | 0.006 | BA | Ca | En | 2, 11 |
| Microporella arctica (Norman, 1903) | MiAr | 0.076 | 0.051 | 0.007 | 0.479 | B | Ca | En | 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 |
| Porella acutirostris (Smitt, 1868) | PoAc | 0.020 | 0.013 | 0.004 | 0.047 | BA | Ca | En | 9, 10, 13 |
| Porella aperta (Boeck, 1862) | PoAp | 0.006 | 0.000 | 0.006 | 0.006 | A | Ca | En | 10 |
| Porella belli (Dawson, 1859) | PoBe | 0.003 | 0.000 | 0.003 | 0.003 | BA | Ca | En | 4 |
| Porella concinna (Busk, 1854) | PoCo | 0.020 | 0.016 | 0.003 | 0.052 | BA | Ca | En | 4, 10, 12 |
| Ragionula rosacea (Busk, 1856) | RaRo | 0.044 | 0.000 | 0.044 | 0.044 | BA | Ca | Er | 10 |
| Rhamphostomella plicata (Smitt, 1868) | RhPl | 0.008 | 0.000 | 0.008 | 0.008 | BA | Ca | En | 10 |
| Rhamphostomella scabra (O. Fabricius, 1824) | RhSc | 0.018 | 0.000 | 0.018 | 0.018 | BA | Ca | En | 10 |
| Rhamphostomella spinigera (Lorenz, 1886) | RhSp | 0.006 | 0.000 | 0.006 | 0.006 | A | Ca | En | 10 |
| Schizomavella (Schizomavella) auriculata (Hassall, 1842) | ScAu | 0.107 | 0.100 | 0.007 | 0.207 | B | Ca | En | 10, 14 |
| Schizomavella (Schizomavella) lineata (Nordgaard, 1896) | ScLi | 0.061 | 0.042 | 0.019 | 0.102 | BA | Ca | En | 10, 12 |
| Schizomavella porifera (Smitt, 1868) | ScPo | 0.001 | 0.000 | 0.001 | 0.001 | BA | Ca | En | 4 |
| Schizoporella costata (Kluge, 1962) | ScCo | 0.006 | 0.004 | 0.002 | 0.010 | BA | Ca | En | 10, 15 |
| Smittina minuscula (Smitt, 1868) | SmMi | 0.001 | 0.000 | 0.001 | 0.001 | BA | Ca | En | 10 |
| Smittina smitti (Kirchenpauer, 1874) | SmSm | 0.025 | 0.000 | 0.025 | 0.025 | A | Ca | En | 10 |
| Stomacrustula cruenta (Busk, 1854) | StCr | 0.005 | 0.002 | 0.003 | 0.006 | BA | Ca | En | 4, 12 |
| Stomacrustula pachystega (Kluge, 1929) | StPa | 0.012 | 0.000 | 0.012 | 0.012 | BA | Ca | En | 10 |
| Tegella arctica (d’Orbigny, 1853) | TeArc | 0.073 | 0.024 | 0.003 | 0.359 | BA | Ca | En | 2, 3, 4, 6, 7, 8, 9, 10, 12, 13, 14, 15, 16, 17, 18 |
| Tegella armifera (Hincks, 1880) | TeArm | 0.010 | 0.003 | 0.004 | 0.019 | BA | Ca | En | 9, 10, 16, 17 |
| Tegella retroversa (Kluge, 1952) | TeRe | 0.043 | 0.023 | 0.002 | 0.129 | BA | Ca | En | 4, 8, 9, 10, 12 |
| Tricellaria elongata (Smitt, 1868) | TrEl | 0.039 | 0.035 | 0.004 | 0.074 | A | Ca | Er | 10, 15 |
| Tricellaria gracilis (Van Beneden, 1848) | TrGr | 0.667 | 0.375 | 0.001 | 3.123 | BA | Ca | Er | 4, 9, 10, 12, 13, 14, 15, 16, 17 |
| Tricellaria ternata (Ellis and Solander, 1786) | TrTe | 0.544 | 0.191 | 0.002 | 1.443 | BA | Ca | Er | 3, 9, 10, 11, 12, 13, 14, 15 |
| Turbicellepora incrassata (Lamarck, 1816) | TuIn | 4.360 | 0.000 | 4.360 | 4.360 | A | Ca | Er | 10 |
| Species | Average Dissimilarity, % | Contribution, % | Cumulative Contribution, % |
|---|---|---|---|
| Eucratea loricata | 6.05 | 9.49 | 9.49 |
| Tricellaria ternata | 4.99 | 7.82 | 17.30 |
| Tricellaria gracilis | 3.58 | 5.61 | 22.92 |
| Alcyonidium hirsutum | 3.58 | 5.60 | 28.52 |
| Cribrilina spitzbergensis | 3.41 | 5.35 | 33.87 |
| Microporella arctica | 2.87 | 4.50 | 38.38 |
| Crisiella diversa | 2.52 | 3.95 | 42.33 |
| Einhornia arctica | 2.25 | 3.53 | 45.86 |
| Callopora lineata | 2.24 | 3.50 | 49.37 |
| Amathia imbricata | 2.15 | 3.36 | 52.73 |
| Celleporella hyalina | 2.10 | 3.29 | 56.02 |
| Alcyonidioides mytili | 1.64 | 2.58 | 58.59 |
| Dendrobeania fruticosa | 1.61 | 2.52 | 61.12 |
| Disporella hispida | 1.55 | 2.43 | 63.54 |
| Harmeria scutulata | 1.52 | 2.39 | 65.93 |
| Juxtacribrilina annulata | 1.49 | 2.34 | 68.27 |
| Crisiella producta | 1.46 | 2.30 | 70.57 |
| Variable | Diversity | Biomass | |||||
|---|---|---|---|---|---|---|---|
| LambdaA | F | p | Variable | LambdaA | F | p | |
| T | 21 | 12.32 | 0.001 | T | 13 | 2.39 | 0.042 |
| Macrophytes | 17 | 3.19 | 0.081 | Macrophytes | 8 | 2.08 | 0.030 |
| Gravel | 13 | 2.80 | 0.099 | Depth | 6 | 1.43 | 0.187 |
| Pebble | 3 | 1.50 | 0.227 | Gravel | 5 | 1.24 | 0.274 |
| Sand | 1 | 0.59 | 0.624 | S | 4 | 1.09 | 0.355 |
| Boulders | 1 | 0.68 | 0.581 | Sand | 4 | 0.83 | 0.525 |
| S | 1 | 0.47 | 0.712 | Pebble | 3 | 1.02 | 0.416 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evseeva, O.Y.; Dvoretsky, A.G. Shallow-Water Bryozoan Communities in a Glacier Fjord of West Svalbard, Norway: Species Composition and Effects of Environmental Factors. Biology 2023, 12, 185. https://doi.org/10.3390/biology12020185
Evseeva OY, Dvoretsky AG. Shallow-Water Bryozoan Communities in a Glacier Fjord of West Svalbard, Norway: Species Composition and Effects of Environmental Factors. Biology. 2023; 12(2):185. https://doi.org/10.3390/biology12020185
Chicago/Turabian StyleEvseeva, Olga Yu., and Alexander G. Dvoretsky. 2023. "Shallow-Water Bryozoan Communities in a Glacier Fjord of West Svalbard, Norway: Species Composition and Effects of Environmental Factors" Biology 12, no. 2: 185. https://doi.org/10.3390/biology12020185
APA StyleEvseeva, O. Y., & Dvoretsky, A. G. (2023). Shallow-Water Bryozoan Communities in a Glacier Fjord of West Svalbard, Norway: Species Composition and Effects of Environmental Factors. Biology, 12(2), 185. https://doi.org/10.3390/biology12020185