
Cytogenetics, Typification, Molecular Phylogeny and Biogeography of Bentinckia (Arecoideae, Arecaceae), an Unplaced Indian Endemic Palm from Areceae

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
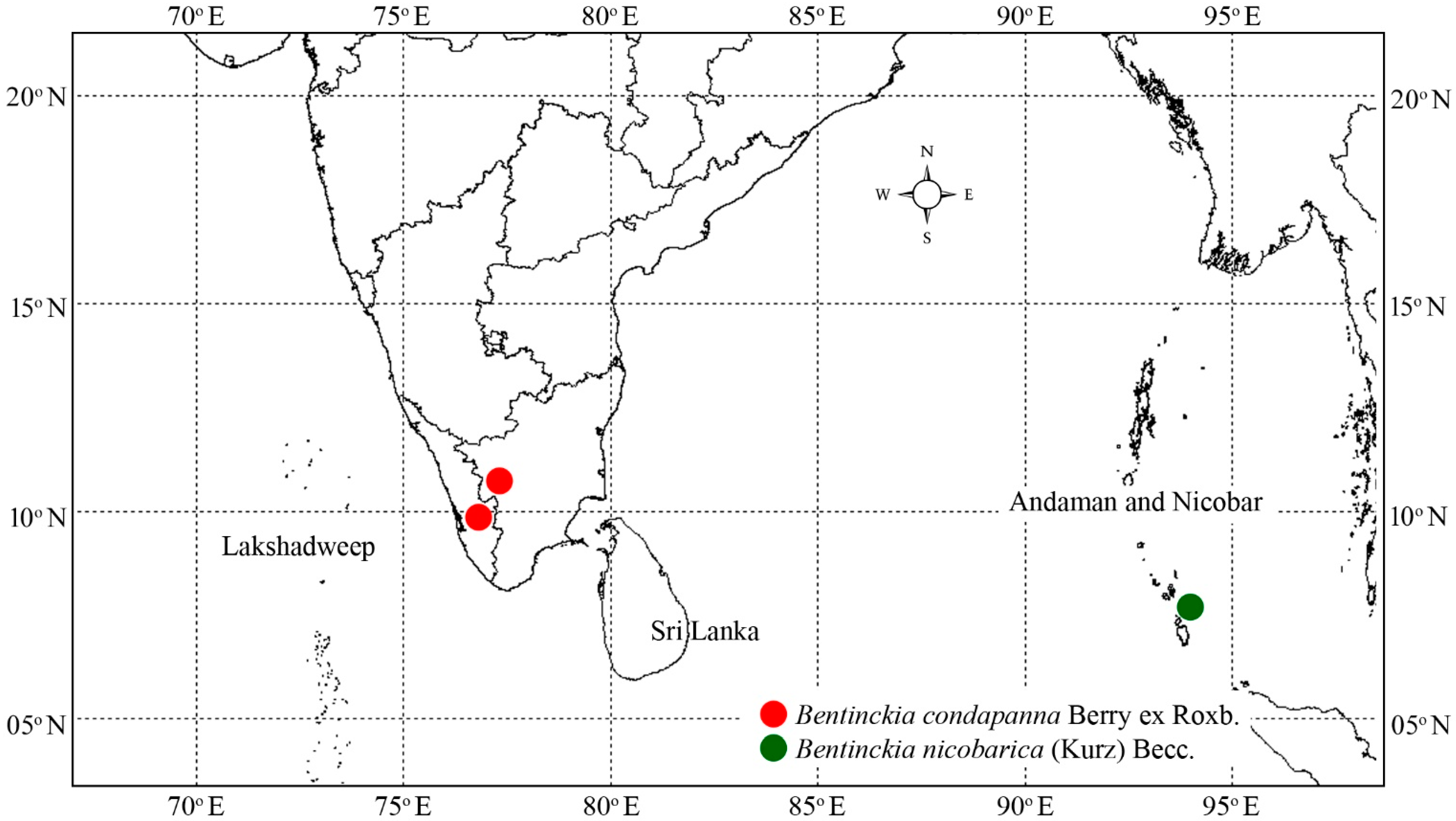
2.1. Taxon Sampling
2.2. Cytogenetics
2.3. DNA Extraction, PCR, and Sequencing
2.4. Phylogenetic Analysis
2.5. Biogeographic Analysis
3. Results
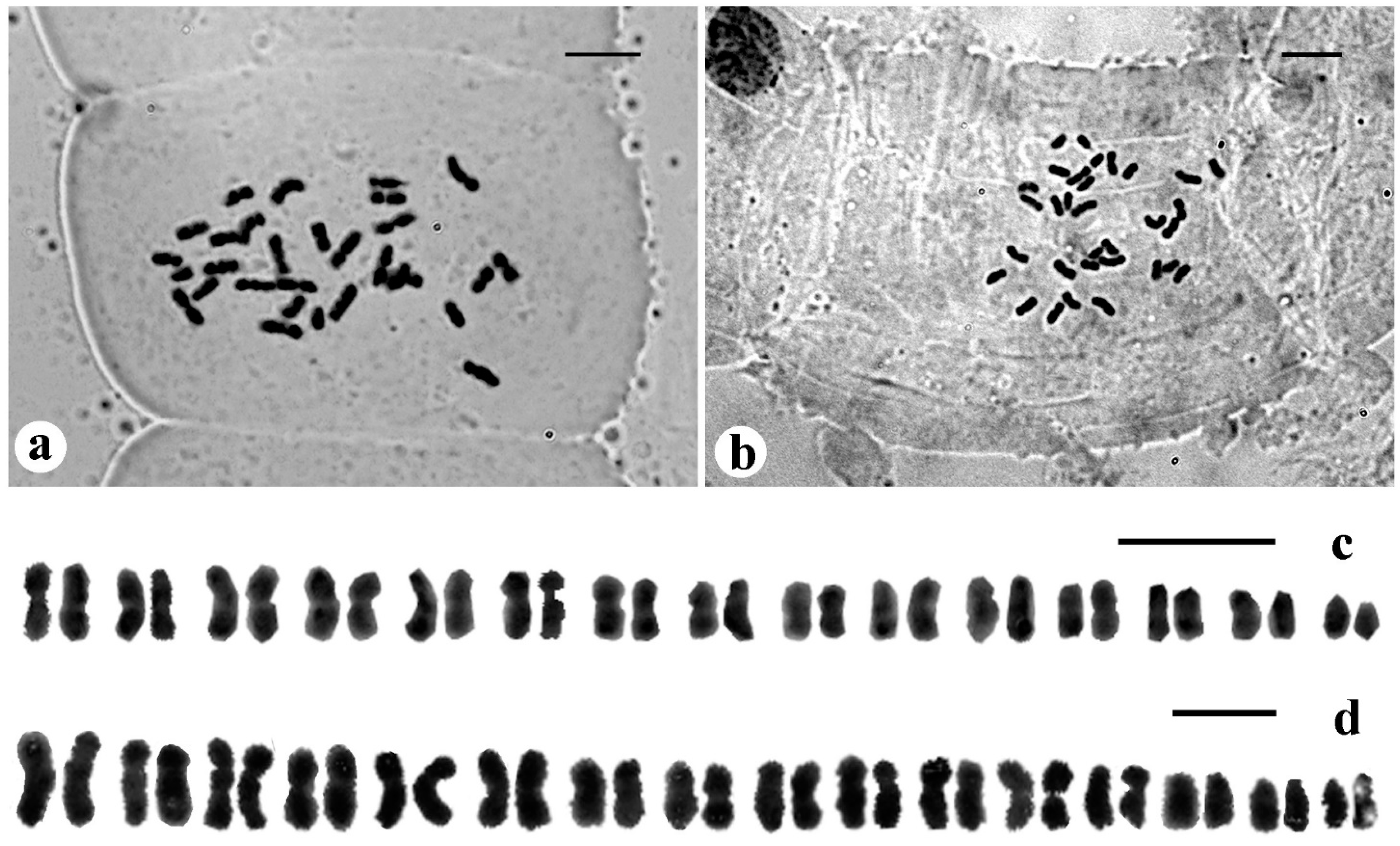
3.1. Cytogenetics
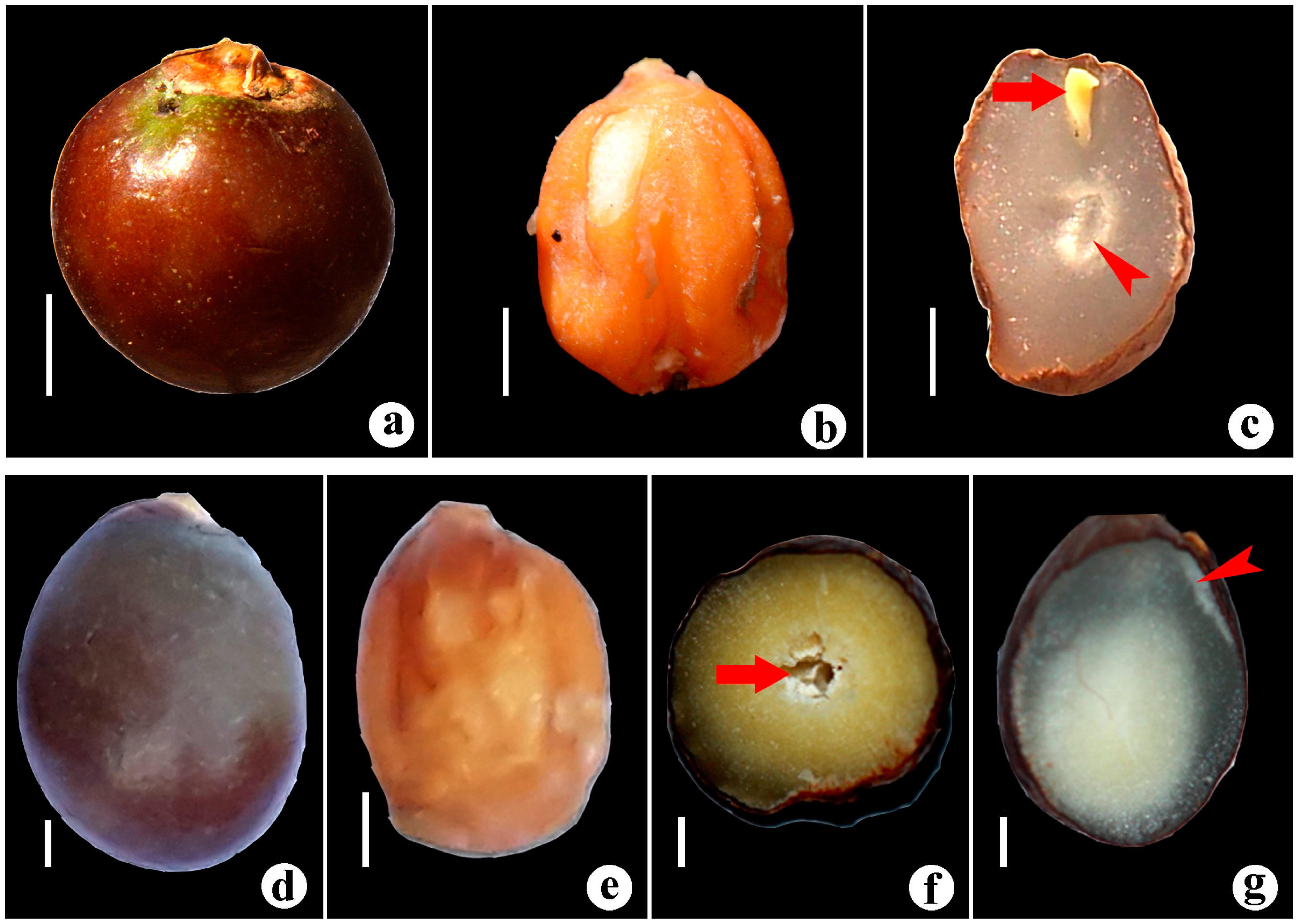
3.1.1. Bentinckia Condapanna Berry Ex Roxb
3.1.2. Bentinckia Nicobarica (Kurz) Becc
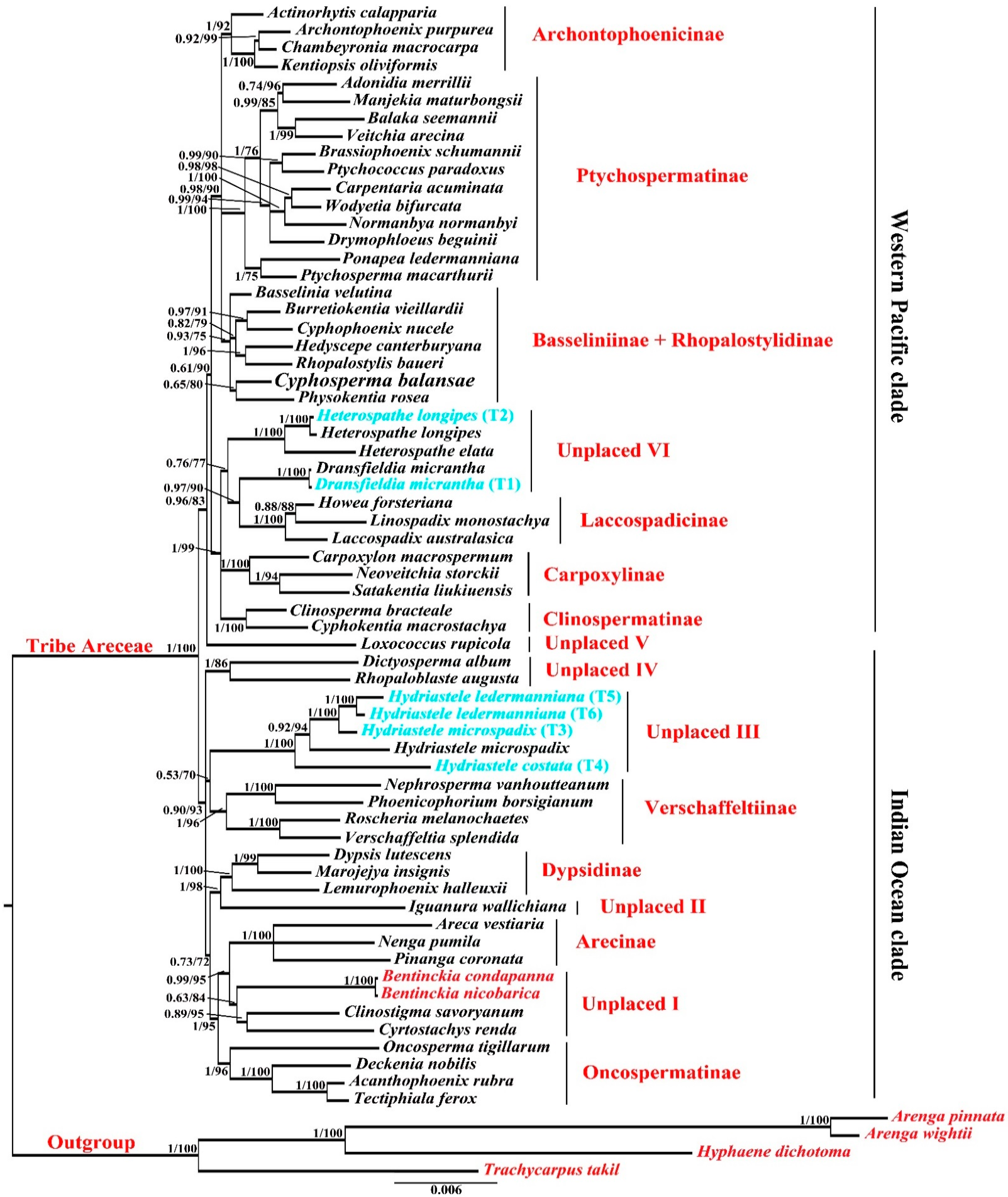
3.2. Molecular Phylogeny of Areceae
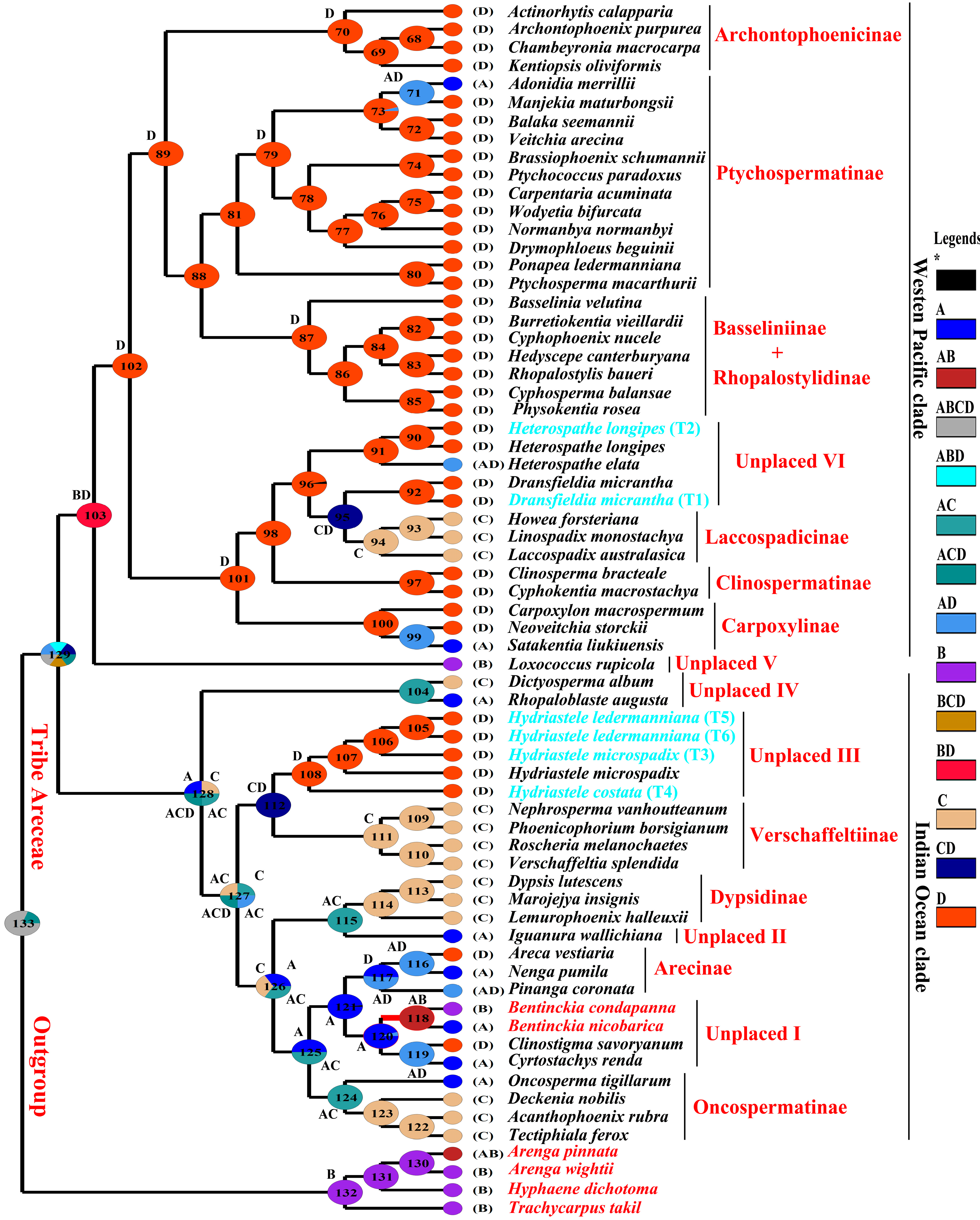
3.3. Ancestral Area Reconstruction
4. Discussion
4.1. Taxonomic Treatment of Bentinckia
4.2. Cytogenetics
4.3. Molecular Phylogeny of Areceae
Unplaced Areceae
4.4. Ancestral Area Reconstruction of Areceae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dransfield, J.; Uhl, N.W.; Asmussen, C.B.; Baker, W.J.; Harley, M.M.; Lewis, C.E. Genera palmarum: The Evolution and Classification of Palms; Royal Botanic Gardens, Kew: London, UK, 2008. [Google Scholar]
- Tomlinson, P.B.; Horn, J.W.; Fisher, J.B. Subfamily Arecoideae. In The Anatomy of Palms: Arecaceae–Palmae; Tregear, J., Ed.; Oxford Academic: London, UK, 2012; pp. 185–240. [Google Scholar]
- Renuka, C.; Sreekumar, V.A. Field Guide to the Palms of India; Kerala Forest Research Institute: Peechi, India, 2012. [Google Scholar]
- Irwin, S.J.; Narasimhan, D. Endemic Genera of Angiosperms in India: A review. Rheedea 2011, 21, 87–105. [Google Scholar]
- Basu, A.; Sarkar, M.K.; Muthukrishnan, A.; Arockiasamy, D.I. Recovery Plan for Bentinckia Condapanna Berry—An Endemic and Threatened Tree Species (palm) of Tropical Semi-Evergreen forests in Southern Western Ghats, India. Indian For. 2014, 140, 439–447. [Google Scholar]
- Ved, D.K. List of Medicinal Plants in Tamil Nadu; Foundation for Revitalisation of Local Health Traditions (FRLHT): Bangalore, India, 2000. [Google Scholar]
- Kulkarni, A.R.; Mulani, R.M. Indigenous palms of India. Curr. Sci. 2004, 86, 1598–1603. [Google Scholar]
- Mathew, S.P.; Abraham, S. The vanishing palms of the Andaman and Nicobar Islands, India. Principes 1994, 38, 100–104. [Google Scholar]
- Roxburgh, W. Flora Indica; Carey, W., Ed.; Cambridge University Press: Calcutta, India, 1832; Volumes 1–3. [Google Scholar]
- Trimen, H.; Baker, J.G. Description of new plants from Nicobar Islands. J. Bot. Brit. Foreign 1875, 13, 331–332. [Google Scholar]
- Beccari, O. Reliquiae Schefferianae illustrazione di alcune palme viventi nel Giardino Botanico di Buitenzorg. Ann. Jard. Bot. Buitenzorg 1885, 2, 77–171. [Google Scholar]
- Renuka, C. Palms of Kerala; Kerala Forest Research Institute: Thrissur, India, 1999; p. 72. [Google Scholar]
- Varghese, A.O.; Menon, A.R. Ecological niches and amplitudes of rare, threatened and endemic trees of Peppara Wildlife Sanctuary. Curr. Sci. 1997, 76, 1204–1208. [Google Scholar]
- Basu, S.K. Red Data Book of Indian Plants; Nayar, M.P., Sastry, A.R.K., Eds.; B.S.I.: Calcutta, India, 1988; Volume 32, p. 268.
- Read, R.W. More chromosomes counts by mail. Principes 1967, 11, 77. [Google Scholar]
- Roser, M. Variation and evolution of karyotype characters in palm subfamily Coryphoideae s.l. Bot. Acta 1993, 106, 170–182. [Google Scholar] [CrossRef]
- Roser, M. Pathways of karyological differentiation in Palms (Arecaceae). Plant Syst. Evol. 1994, 189, 83–122. [Google Scholar] [CrossRef]
- Sharma, A.K. Annual report, 1967–1968. Res. Bull. Univ. Calcutta. Cytogenet. Lab. 1970, 2, 1–50. [Google Scholar]
- Read, R.W. New chromosome counts in the Palmae. Principes 1966, 10, 55–61. [Google Scholar]
- Kadam, S.K.; Tamboli, A.S.; Mane, R.N.; Yadav, S.R.; Choo, Y.-S.; Burgos-Hernández, M.; Pak, J.H. Revised molecular phylogeny, global biogeography, and diversification of palms subfamily Coryphoideae (Arecaceae) based on low copy nuclear and plastid regions. J. Plant Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Rao, R.R. Handbook of Field and Herbarium Methods; Today & Tommorrow Printers and Publishers: New Delhi, India, 1977; pp. 50–80. [Google Scholar]
- Mane, R.N.; Yadav, S.R. Karyomorphological Analysis of Syzygium laetum (Myrtaceae) from India. Cytologia 2022, 87, 227–230. [Google Scholar] [CrossRef]
- Levan, A.; Fredge, K.; Sandberg, A. Nomenclature for centromeric positions on chromosomes. Hereditas 1964, 52, 201. [Google Scholar] [CrossRef]
- Stebbin, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold: London, UK, 1971. [Google Scholar]
- Peruzzi, L.; Eroğlu, H.E. Karyotype asymmetry: Again, how to measure and what to measure? Comp. Cytogenet. 2013, 7, 1–9. [Google Scholar] [CrossRef]
- Paterson, A.; Brubaker, C.; Wendel, J. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Rep. 1993, 11, 122–127. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D.J. Model Test 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772–775. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed 451 models. Bioinformatics 2012, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucl. Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Baker, W.J.; Couvreur, T.L.P. Global biogeography and diversification of palms sheds light on the evolution of tropical lineages. I. historical biogeography. J. Biogeogr. 2013, 40, 274–285. [Google Scholar] [CrossRef]
- Baker, W.J.; Couvreur, T.L.P. Global biogeography and diversification of palms sheds light on the evolution of tropical lineages. II. Historical biogeography. J. Biogeogr. 2013, 40, 286–298. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D. Bentinckia condapanna. IUCN Red List Threat. Species 1998, e.T38449A10119906. [Google Scholar] [CrossRef]
- Mane, R.N.; Lekhak, M.M. Lectotypification of Corypha taliera Roxburgh and Orania nicobarica Kurz (Arecaceae). Phytotaxa 2021, 489, 236–239. [Google Scholar] [CrossRef]
- Kurz, W.S. Descriptions of new plants from the Nicobar Islands (including a few from the Andaman Islands). J. Bot. 1875, 13, 321–326. [Google Scholar]
- Johnson, D. Bentinckia nicobarica. IUCN Red List Threat. Species 1998, e.T38450A10120169. [Google Scholar] [CrossRef]
- Hahn, W.J. A phylogenetic analysis of the Arecoid Line of palms based on plastid DNA sequence data. Mol. Phylogenet. Evol. 2002, 23, 189–204. [Google Scholar] [CrossRef]
- Lewis, C.E.; Doyle, J.J. A phylogenetic analysis of tribe Areceae (Arecaceae) using two low-copy nuclear genes. Plant Syst. Evol. 2002, 236, 1–17. [Google Scholar] [CrossRef]
- Loo, A.H.B.; Dransfield, J.; Chase, M.W.; Baker, W.J. Low copy nuclear DNA, phylogeny and the evolution of dichogamy in the betel nut palms and their relatives (Arecinae, Arecaceae). Mol. Phylogenet. Evol. 2006, 39, 598–618. [Google Scholar] [CrossRef] [PubMed]
- Norup, M.V.; Dransfield, J.; Chase, M.W.; Barfod, A.S.; Fernando, E.S.; Baker, W.J. Homoplasious character combinations and generic delimitations: A case study from the Indo-Pacific arecoid palms (Arecaceae: Areceae). Am. J. Bot. 2006, 93, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Baker, W.J.; Norup, M.V.; Clarkson, J.J.; Couvreur, T.L.P.; Dowe, J.L.; Lewis, C.E.; Pintaud, J.C.; Savolainen, V.; Wilmot, T.; Chase, M.W. Phylogenetic relationships among arecoid palms (Arecaceae: Arecoideae). Ann. Bot. 2011, 108, 1417–1432. [Google Scholar] [CrossRef] [PubMed]
- Uhl, N.W.; Dransfield, J.; Davis, J.I.; Luckow, M.A.; Hansen, K.S.; Doyle, J.J. Phylogenetic relationships among palms: Cladistic analyses of morphological and chloroplast DNA restriction site variation. In Monocotyledons: Systematics and Evolution; Rudall, P.J., Cribb, P.J., Cutler, D.F., Humphries, C.J., Eds.; Royal Botanic Gardens, Kew: London, UK, 1995; pp. 623–661. [Google Scholar]
- Asmussen, C.B.; Chase, M.W. Coding and noncoding plastid DNA in palm systematics. Am. J. Bot. 2001, 88, 1103–1117. [Google Scholar] [CrossRef]
- Lewis, C.E.; Doyle, J.J. Phylogenetic utility of the nuclear gene malate synthase in the palm family (Arecaceeae). Mol. Phylogenet. Evol. 2001, 19, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Asmussen, C.B.; Dransfield, J.; Deickmann, V.; Barfod, A.S.; Pintaud, J.C.; Baker, W.J. A new subfamily classification of the palm family (Arecaceae): Evidence from plastid DNA phylogeny. Bot. J. Linn. Soc. 2006, 151, 15–38. [Google Scholar] [CrossRef]
- Baker, W.J.; Savolainen, V.; Asmussen-Lange, C.B.; Chase, M.W.; Dransfield, J.; Forest, F.; Harley, M.M.; Uhl, N.W.; Wilkinson, M. Complete generic-level phylogenetic analyses of palms (Arecaceae) with comparisons of supertree and supermatrix approaches. Syst. Biol. 2009, 58, 240–256. [Google Scholar] [CrossRef]
- Comer, J.R.; Zomlefer, W.B.; Barrett, C.F.; Davis, J.I.; Stevenson, D.W.; Heyduk, K.; Leebens-Mack, J.H. Resolving relationships within the palm subfamily Arecoideae (Arecaceae) using plastid sequences derived from next-generation sequencing. Am. J. Bot. 2015, 102, 888–899. [Google Scholar] [CrossRef]
- Comer, J.R.; Zomlefer, W.B.; Barrett, C.F.; Stevenson, D.W.; Heyduk, K.; Leebens-Mack, J.H. Nuclear phylogenomics of the palm subfamily Arecoideae (Arecaceae). Mol. Phylogenet. Evol. 2016, 97, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Faurby, S.; Eiserhardt, W.L.; Baker, W.J.; Svenning, J. An all-evidence species-level supertree for the palms (Arecaceae). Mol. Phylogenet. Evol. 2016, 100, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Petoe, P.; Heatubun, C.D.; Baker, W.J. A monograph of Hydriastele (Areceae, Arecaceae) in New Guinea and Australia. Phytotaxa 2018, 370, 1–92. [Google Scholar] [CrossRef]
- Plants of the World. Kew Science. 2022. Available online: www.plantssoftheworldonline.org (accessed on 12 December 2022).
- Baker, W.J.; Loo, A.H.B. A synopsis of genus Hydriastele. Kew Bull. 2004, 59, 61–68. [Google Scholar] [CrossRef]
- Baker, W.J.; Zona, S.; Heatubun, C.D.; Lewis, C.E.; Maturbongs, R.A.; Norup, M.V. Dransfieldia (Arecaceae)—A New Palm Genus from Western New Guinea. Syst. Bot. 2006, 31, 61–69. [Google Scholar] [CrossRef]
- Norup, M.V. Alsmithia Subsumed in Heterospathe (Arecaceae, Arecoideae); Missouri Botanical Garden Press: St. Louis, MI, USA, 2005; Volume 15, pp. 455–457. [Google Scholar]
- Ali, S.S.; Yu, Y.; Pfosser, M.; Wetschnig, W. Inferences of biogeographical histories within subfamily Hyacinthoideae using S-DIVAand Bayesian binary MCMC analysis implemented in RASP (Reconstruct Ancestral State in Phylogenies). Ann. Bot. 2012, 109, 95–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. condapanna | B. nicobarica | |
|---|---|---|
| 2n | 30 | 32 |
| Total haploid chromosome length (THCL ± SE (µm)) | 34.04 ± 0.59 | 25.83 ± 0.40 |
| Haploid karyotype formula | 15m | 9m + 7sm |
| Coefficient of Variation of Chromosome Length (CVCL) | 18.18 | 27.17 |
| Coefficient of Variation of Centromeric Index (CVCI) | 8.05 | 11.17 |
| Mean Centromeric Asymmetry (MCA) | 14.05 | 22.02 |
| Shortest chromosome (S ± SE (µm)) | 1.48 ± 0.34 | 0.91 ± 0.25 |
| Longest chromosome (L ± SE (µm)) | 3.00 ± 0.25 | 2.30 ± 0.16 |
| Mean chromosome length (MCL ± SE (µm)) | 1.62 ± 0.59 | 1.61 ± 0.11 |
| Longest to shortest chromosome ratio (R) | 2.03 | 2.52 |
| Stebbins category (St) | 4B | 3B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadam, S.K.; Mane, R.N.; Tamboli, A.S.; Gavade, S.K.; Deshmukh, P.V.; Lekhak, M.M.; Choo, Y.-S.; Pak, J.H. Cytogenetics, Typification, Molecular Phylogeny and Biogeography of Bentinckia (Arecoideae, Arecaceae), an Unplaced Indian Endemic Palm from Areceae. Biology 2023, 12, 233. https://doi.org/10.3390/biology12020233
Kadam SK, Mane RN, Tamboli AS, Gavade SK, Deshmukh PV, Lekhak MM, Choo Y-S, Pak JH. Cytogenetics, Typification, Molecular Phylogeny and Biogeography of Bentinckia (Arecoideae, Arecaceae), an Unplaced Indian Endemic Palm from Areceae. Biology. 2023; 12(2):233. https://doi.org/10.3390/biology12020233
Chicago/Turabian StyleKadam, Suhas K., Rohit N. Mane, Asif S. Tamboli, Sandip K. Gavade, Pradip V. Deshmukh, Manoj M. Lekhak, Yeon-Sik Choo, and Jae Hong Pak. 2023. "Cytogenetics, Typification, Molecular Phylogeny and Biogeography of Bentinckia (Arecoideae, Arecaceae), an Unplaced Indian Endemic Palm from Areceae" Biology 12, no. 2: 233. https://doi.org/10.3390/biology12020233
APA StyleKadam, S. K., Mane, R. N., Tamboli, A. S., Gavade, S. K., Deshmukh, P. V., Lekhak, M. M., Choo, Y. -S., & Pak, J. H. (2023). Cytogenetics, Typification, Molecular Phylogeny and Biogeography of Bentinckia (Arecoideae, Arecaceae), an Unplaced Indian Endemic Palm from Areceae. Biology, 12(2), 233. https://doi.org/10.3390/biology12020233