

Role of Alternative Elicitor Transporters in the Onset of Plant Host Colonization by Streptomyces scabiei 87-22
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Culture Conditions
2.2. In Silico Analysis
2.3. Transcriptomics
2.4. Construction of Deletion Mutants
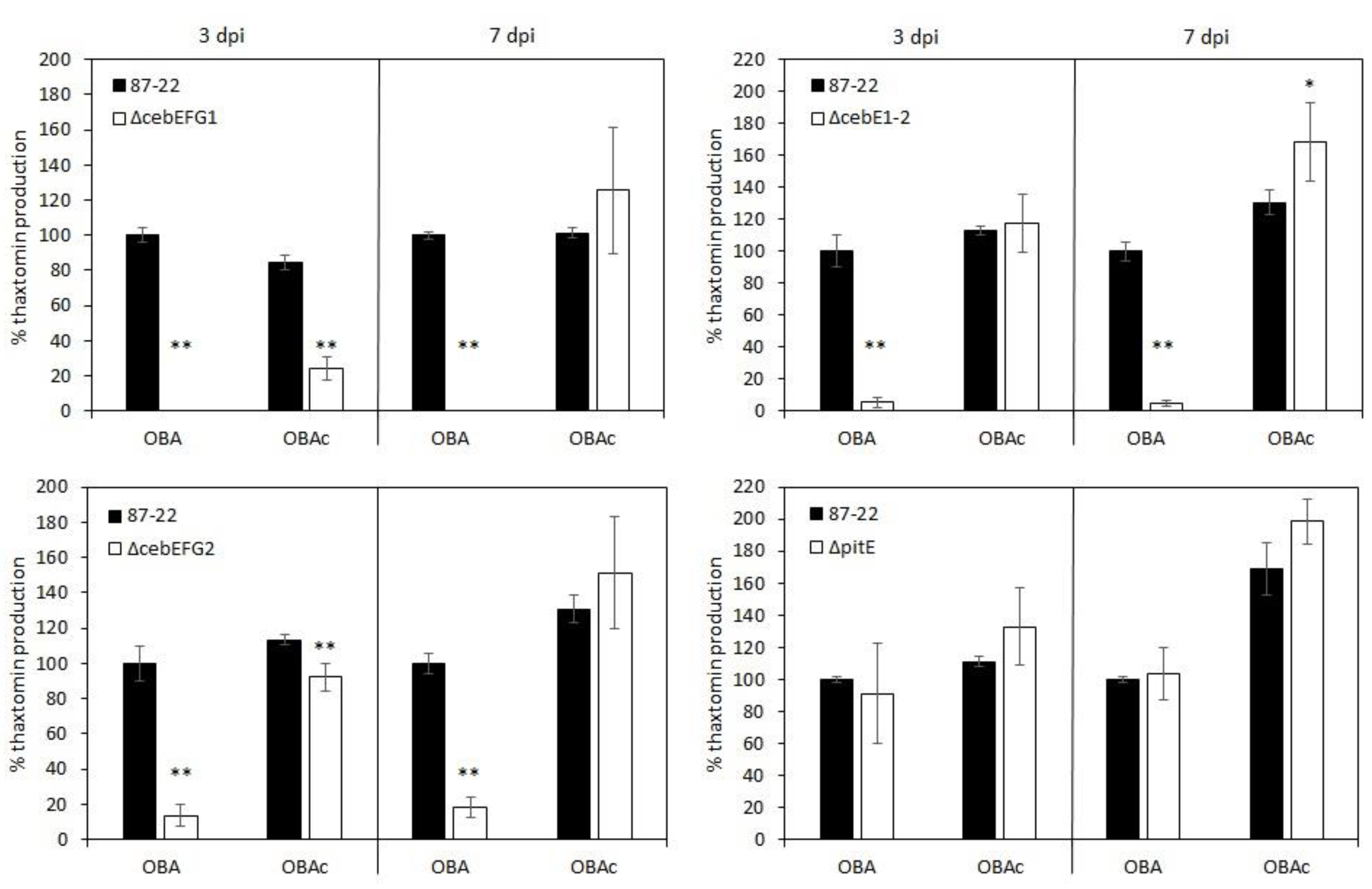
2.5. Analysis of Thaxtomin Production
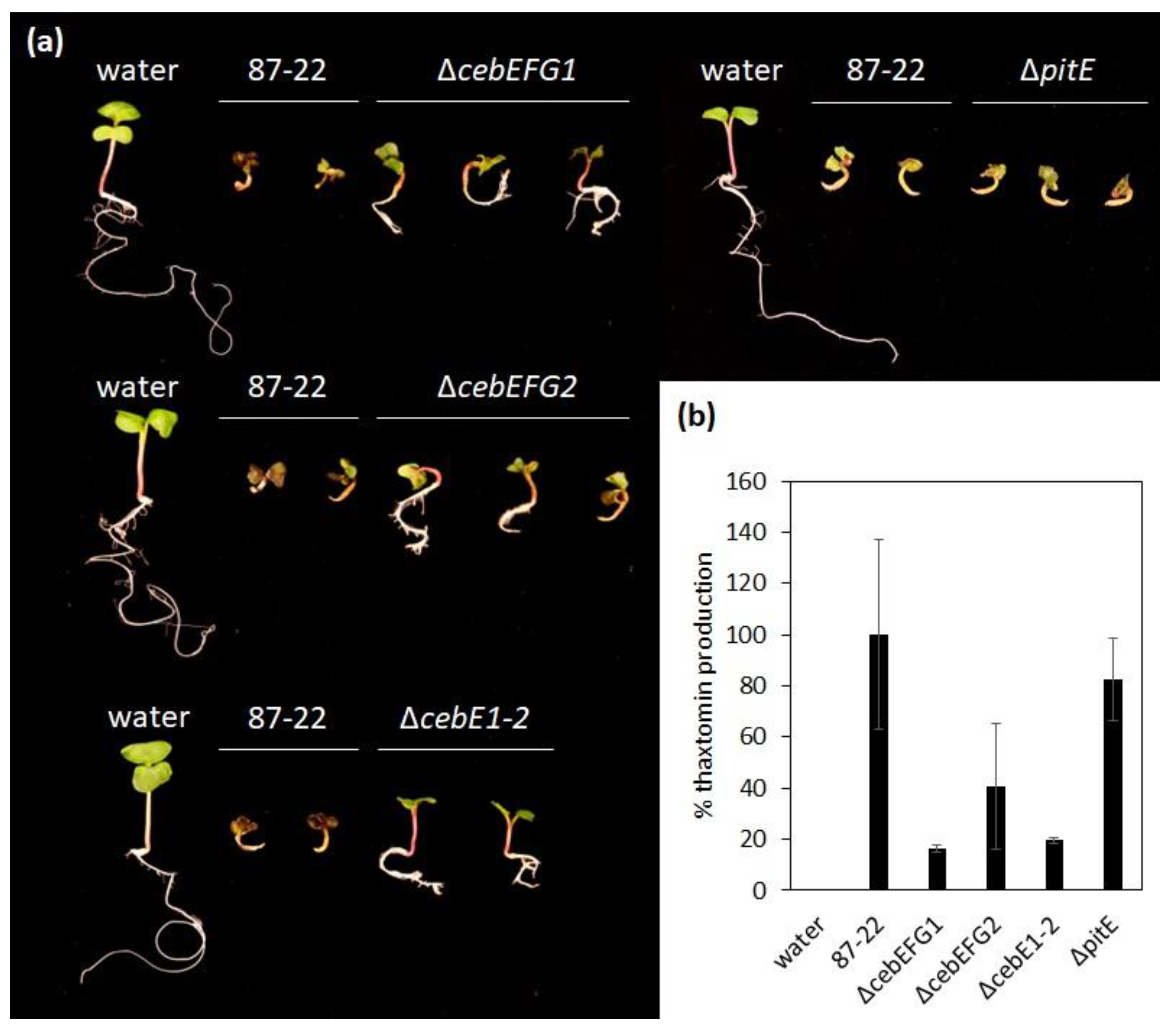
2.6. Virulence Assays
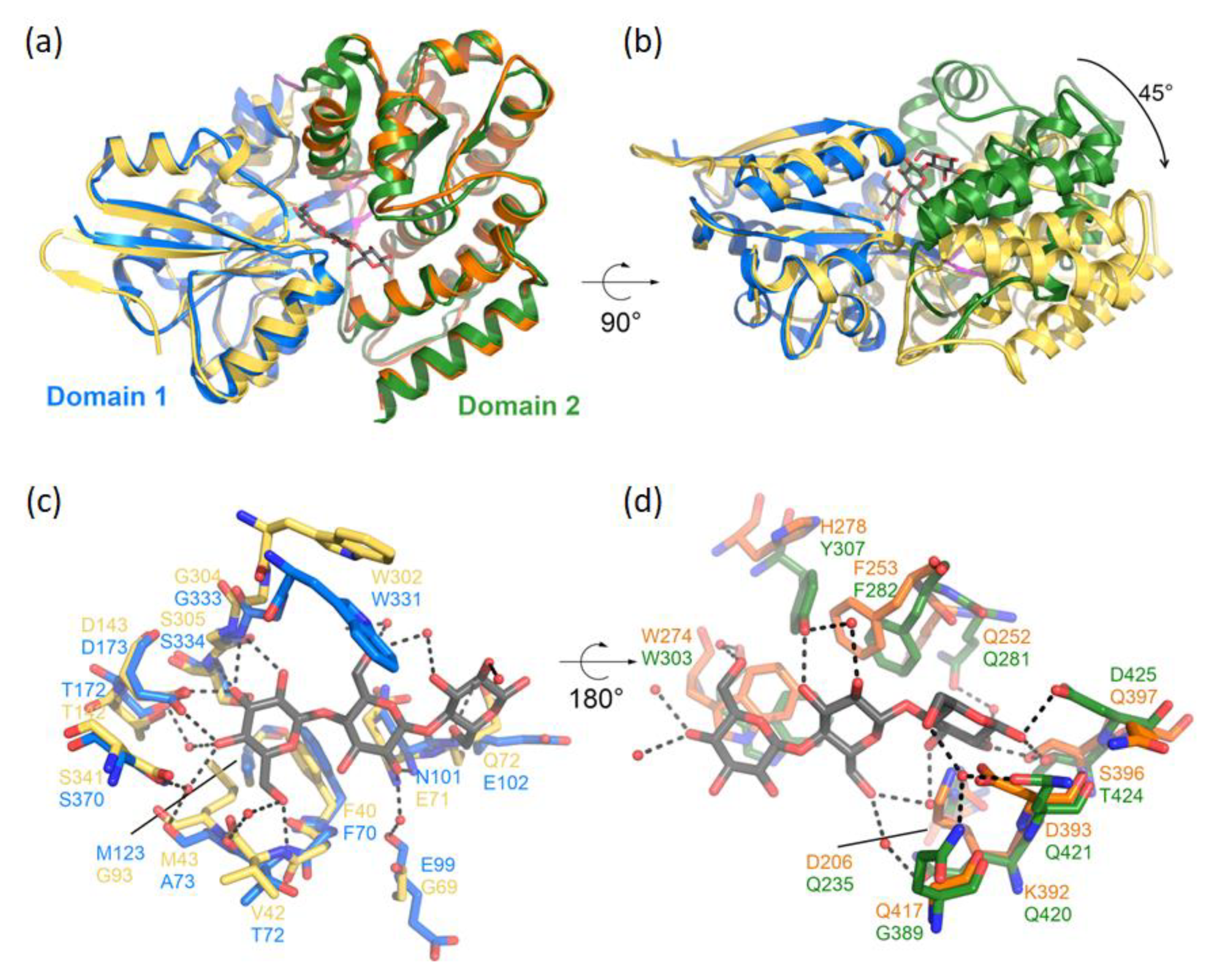
2.7. Modeling of CebE2
3. Results
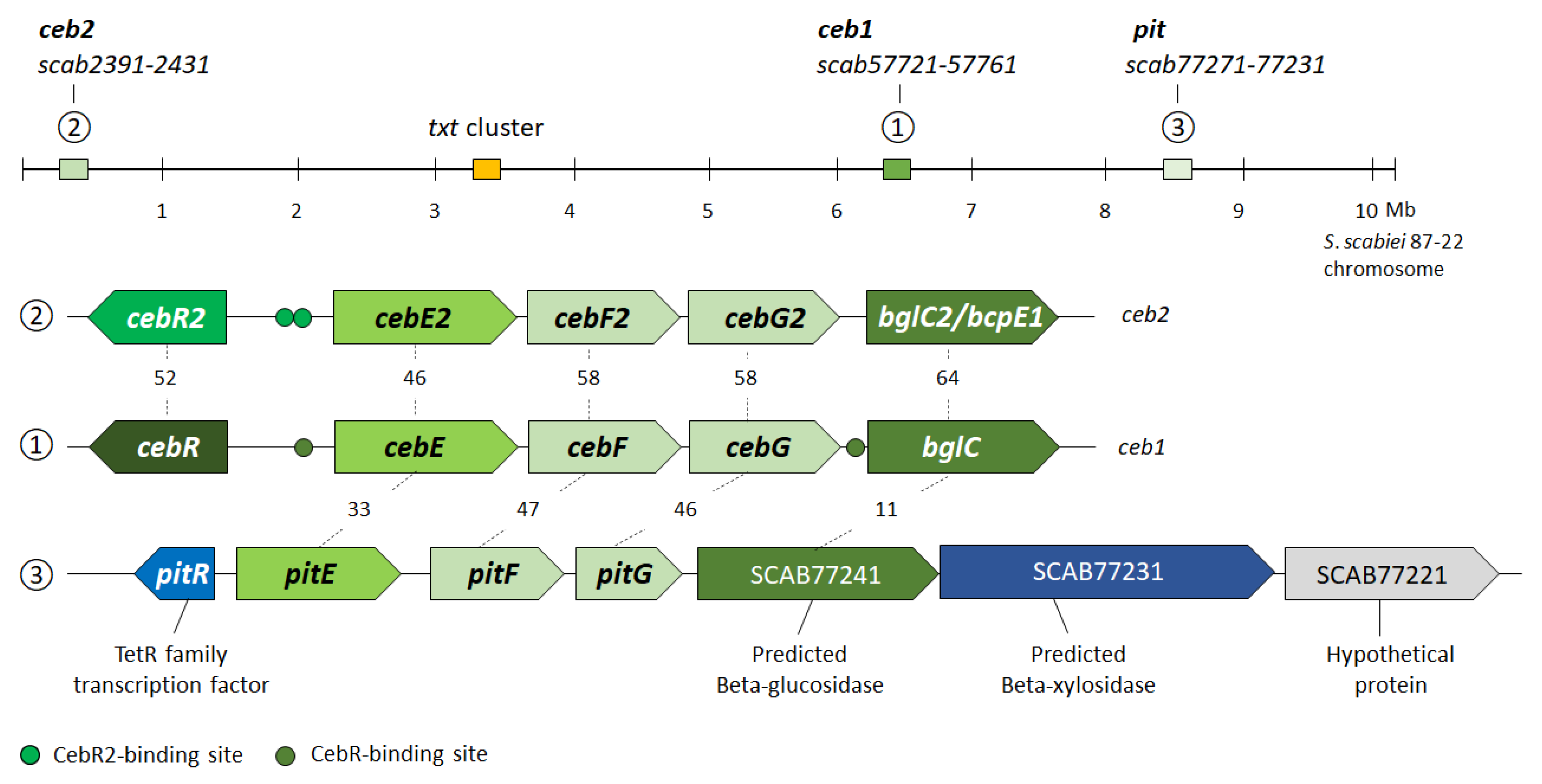
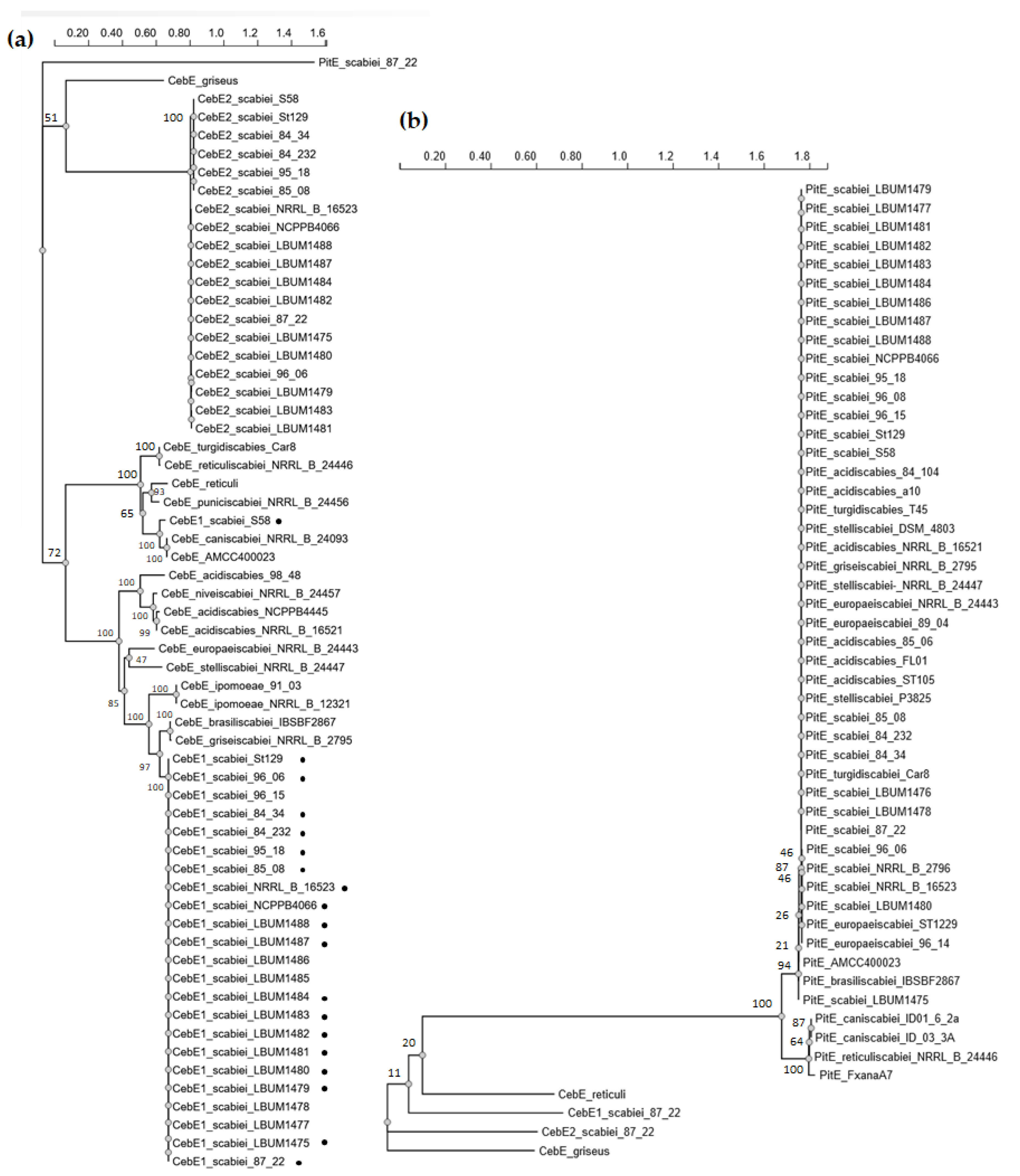
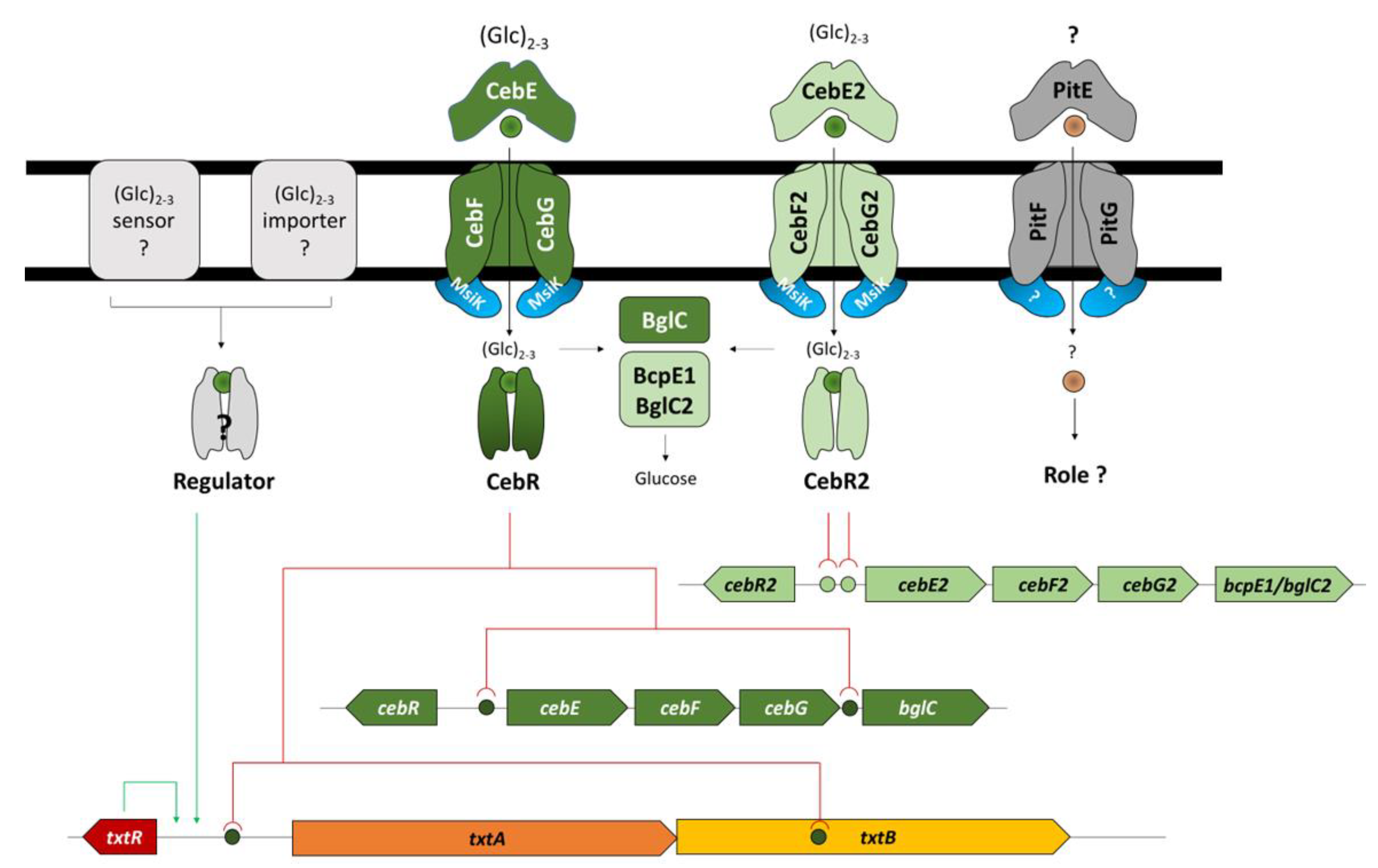
3.1. In Silico Analysis of CebE Homologs of Streptomyces scabiei
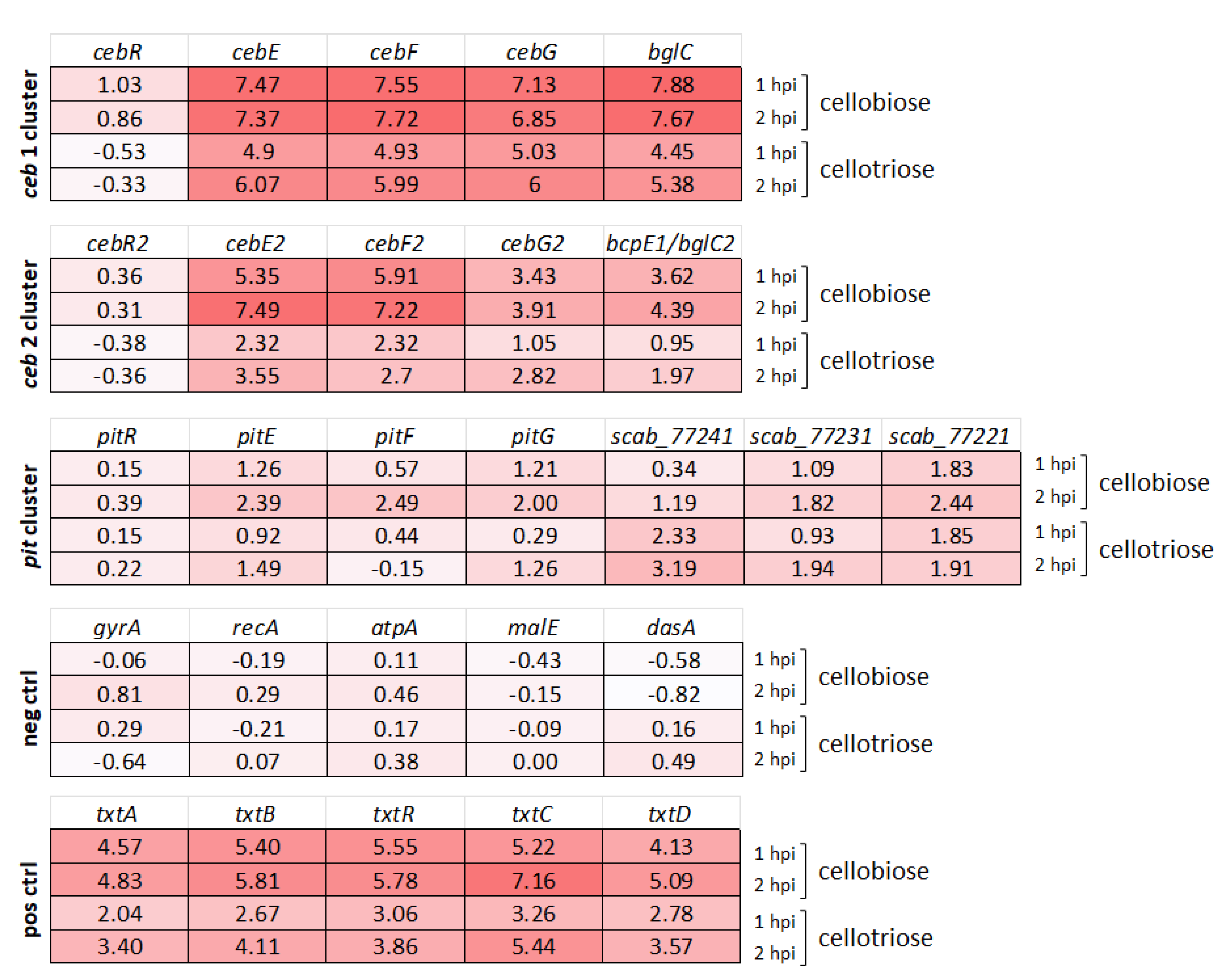
3.2. Transcriptional Response of Alternative Cello-Oligosaccharide Transporters to Cellobiose and Cellotriose
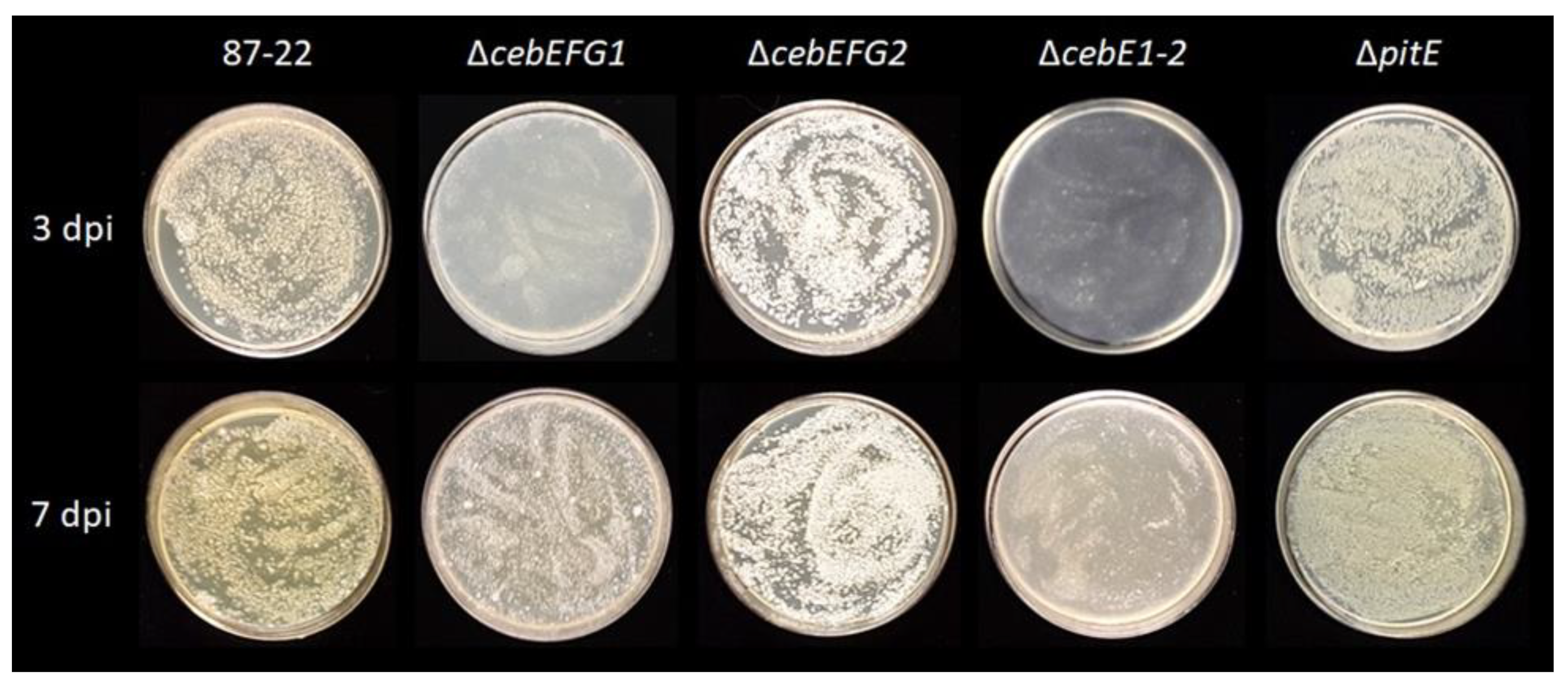
3.3. Role of the CebEFG2 and PitEFG Transporters in Virulence Elicitor Utilization by Streptomyces scabiei
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description † | Source or Reference | |
|---|---|---|
| Plasmids/cosmids | ||
| pIJ790 | λ Red plasmid (tS, CmlR) | [19] |
| pUZ8002 | Supplies transfer functions for mobilization of oriT-containing vectors from E. coli to Streptomyces (KanR) | [19] |
| pIJ773 | Template for REDIRECT© PCR targeting system, contains the [aac(3)IV + oriT] disruption cassette (AmpR, ApraR) | [19] |
| pIJ778 | Template for REDIRECT© PCR targeting system, contains the [aadA + oriT] disruption cassette (AmpR, StrepR, SpecR) | [19] |
| SuperCos1 | Cosmid cloning vector (AmpR, KanR) | Stratagene |
| Cosmid 833 | SuperCos1 derivative containing the S. scabiei 87-22 cellobiose/cellotriose-specific ABC-transporter locus (ceb1) (KanR, AmpR) | [12] |
| Cosmid 2852 | SuperCos1 derivative containing the S. scabiei 87-22 ceb2 ABC-transporter locus (KanR, AmpR) | This study |
| Cosmid 1998 | SuperCos1 derivative containing the S. scabiei 87-22 pit ABC-transporter locus (KanR, AmpR) | This study |
| E. coli strains | ||
| DH5α | General cloning host | Gibco-BRL |
| BW25113 | Host for Redirect PCR targeting system | [19] |
| ET12567 | dam−, dcm−, hsdS−; non-methylating host for transfer of DNA into Streptomyces spp. (CmlR, TetR) | [28] |
| Streptomyces strains | ||
| 87-22 | S. scabiei wild type strain | [29] |
| ∆cebEFG1 | 87-22 derivative with a deletion of the region from scab57751 (cebE1) through scab57731 (cebG1) (ApraR) | This study |
| ∆cebEFG2 | 87-22 derivative with a deletion of the region from scab2421 (cebE2) through scab2401 (cebG2) (ApraR) | This study |
| ∆pitE | 87-22 derivative with a deletion of the scab77271 (pitE) gene (ApraR) | This study |
| Streptomyces | ABC-Type Sugar-Binding Proteins | |||||
|---|---|---|---|---|---|---|
| Species | Strain | CebE1 | CebE2 | PitE | GenBank Assembly | Ref. |
| scabiei | 87-22 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_000091305.1 | [30,31] |
| scabiei | LBUM 1475 | WP_013003368.1 | WP_037723613.1 | WP_237311104.1 | GCA_017933255.1 | [32] |
| scabiei | LBUM 1477 | WP_013003368.1 | NF | WP_006378806.1 | GCA_017969045.1 | [32] |
| scabiei | LBUM 1478 | WP_013003368.1 | NF | MBP5904699.1 | GCA_017969065.1 | [32] |
| scabiei | LBUM 1479 | WP_013003368.1 | WP_060905683.1 | WP_006378806.1 | GCA_017969075.1 | [32] |
| scabiei | LBUM 1480 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_017933235.1 | [32] |
| scabiei | LBUM 1481 | WP_013003368.1 | WP_210550546.1 | WP_006378806.1 | GCA_017969025.1 | [32] |
| scabiei | LBUM 1482 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_017933115.1 | [32] |
| scabiei | LBUM 1483 | WP_013003368.1 | WP_210550546.1 | WP_006378806.1 | GCA_017969005.1 | [32] |
| scabiei | LBUM 1484 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_017968985.1 | [32] |
| scabiei | LBUM 1485 | WP_013003368.1 | NF | NF | GCA_017968925.1 | [32] |
| scabiei | LBUM 1486 | WP_013003368.1 | NF | WP_006378806.1 | GCA_017968965.1 | [32] |
| scabiei | LBUM 1487 | WP_013003368.1 | WP_037723613.1 | WP_006378806.1 | GCA_017968905.1 | [32] |
| scabiei | LBUM 1488 | WP_013003368.1 | WP_037723613.1 | WP_006378806.1 | GCA_017968935.1 | [32] |
| scabiei | NCPPB 4066 | WP_013003368.1 | WP_037723613.1 | WP_006378806.1 | GCA_000738715.1 | [33] |
| scabiei | NRRL B-16523 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_001005405.1 | [34] |
| scabiei | S58 | GAQ65654.1 | WP_059084709.1 | WP_006378806.1 | GCF_001485125.1 | [35] |
| scabiei | St129 | WP_013003368.1 | WP_037723613.1 | WP_006378806.1 | GCA_003584815.1 | [36] |
| scabiei | 84-34 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_001550245.1 | [33] |
| scabiei | 84-232 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_001572115.1 | [33] |
| scabiei | 85-08 | WP_013003368.1 | WP_037723613.1 | WP_231885169.1 | GCA_001550215.1 | [33] |
| scabiei | 95-18 | WP_013003368.1 | WP_037723613.1 | WP_006378806.1 | GCA_001550225.1 | [33] |
| scabiei | 96-06 | WP_013003368.1 | WP_060905683.1 | WP_231885169.1 | GCA_001579685.1 | [33] |
| scabiei | 96-08 | NF | WP_060905683.1 | WP_006378806.1 | GCA_001550235.1 | [33] |
| scabiei | 96-15 | WP_013003368.1 | NF | WP_006378806.1 | GCA_001550295.1 | [33] |
| europaeiscabiei | NCPPB 4086 | WP_037703405.1 | - | NF | GCF_000738695.1 | [33] |
| brasiliscabiei | IBSBF2867 | WP_216591689.1 | - | WP_210534218.1 | GCF_018927715.1 | [37] |
| griseiscabiei | NRRL B-2795 | MBZ3900963.1 | - | WP_006378806.1 | GCA_020010925.1 | [38] |
| ipomoeae | NRRL-B-12321 | WP_141569662.1 | - | NF | GCF_006547165.1 | |
| ipomoeae | 78-51 | WP_009295889.1 | - | NF | GCF_006547175.1 | [39] |
| ipomoeae | 88-35 | WP_009295889.1 | - | NF | GCF_006547185.1 | [39] |
| ipomoeae | 91-03 | WP_009295889.1 | - | NF | GCF_000317595.1 | [40] |
| europaeiscabiei | NRRL B-24443 | WP_046704818.1 | - | WP_006378806.1 | GCF_000988945.1 | [41] |
| europaeiscabiei | 89-04 | WP_046704818.1 | - | WP_006378806.1 | GCF_001550325.1 | [33] |
| europaeiscabiei | St1229 | WP_046704818.1 | - | WP_231885169.1 | GCF_003584825.1 | |
| europaeiscabiei | 96-14 | WP_060891518.1 | - | WP_231885169.1 | GCF_001550375.1 | [36] |
| stelliscabiei | DSM 41803 | WP_046918411.1 | - | WP_006378806.1 | GCF_014873495.1 | |
| stelliscabiei | P3825 | WP_046918411.1 | - | WP_231885169.1 | GCF_001189035.1 | |
| stelliscabiei | NRRL B-24447 | WP_046918411.1 | - | WP_006378806.1 | GCF_001008135.1 | [42] |
| stelliscabiei | 1222.2 | WP_046918411.1 | - | NF | GCF_900215595.1 | |
| acidiscabies | NRRL B-16521 | WP_029183343.1 | - | WP_006378806.1 | GCA_020010905.1 | [38] |
| acidiscabies | 84-104 | WP_029183343.1 | - | WP_006378806.1 | GCA_000242715.2 | [43] |
| acidiscabies | 85-06 | WP_029183343.1 | - | WP_006378806.1 | GCA_001941285.1 | [44] |
| acidiscabies | 98-49 | WP_029183343.1 | - | NF | GCA_001941265.1 | [44] |
| acidiscabies | a2 | WP_029183343.1 | - | NF | GCA_001949645.1 | |
| acidiscabies | a10 | WP_029183343.1 | - | WP_006378806.1 | GCA_001485105.1 | [35] |
| acidiscabies | FL01 | WP_029183343.1 | - | WP_006378806.1 | GCA_001941325.1 | [44] |
| acidiscabies | St105 | WP_029183343.1 | - | WP_006378806.1 | GCA_003584805.1 | |
| acidiscabies | LBUM 1476 | WP_029183343.1 | - | WP_006378806.1 | GCA_017969125.1 | |
| acidiscabies | NCPPB 4445 | WP_050369574 | - | NF | GCA_001189015.1 | |
| acidiscabies | 98-48 | WP_075734941.1 | - | NF | GCA_001941275.1 | [44] |
| niveiscabiei | NRRL B-24457 | WP_055721858.1 | - | NF | GCF_001419795.1 | |
| puniciscabiei | NRRL B-24456 | WP_055706423.1 | - | NF | GCA_001419685.1 | [45] |
| puniciscabiei | DSM 41929 | WP_055706423.1 | - | NF | GCA_006715785.1 | [45] |
| caniscabiei | 96-12 | WP_060884585.1 | - | NF | GCA_001550315.1 | [33] |
| caniscabiei | NRRL B-24093 | WP_060884585.1 | - | NF | GCA_002155765.1 | [34] |
| caniscabiei | ND05-01C | WP_193381044.1 | - | NF | GCA_014930395.1 | |
| caniscabiei | ND05-13A | WP_193381044.1 | - | NF | GCA_014930415.1 | |
| caniscabiei | ND05-3B | WP_193381044.1 | - | NF | GCA_014930355.1 | |
| caniscabiei | NE06-02D | WP_193381044.1 | - | NF | GCA_014930365.1 | [46] |
| caniscabiei | NE06-02F | WP_193381044.1 | - | NF | GCA_014930405.1 | |
| caniscabiei | ID-03-3A | WP_086802801.1 | - | WP_086802416.1 | GCA_014852565.1 | |
| caniscabiei | NRRL B-2801 | WP_086802801.1 | - | WP_237277203.1 | GCA_002155725.1 | |
| caniscabiei | ID01-6.2a | WP_086802801.1 | - | WP_086802416.1 | GCA_014852655.1 | |
| caniscabiei | ID01-12c | WP_086802801.1 | - | NF | GCA_014852675.1 | |
| sp | AMCC400023 | WP_045557721.1 | - | WP_237311104.1 | GCA_021665875.1 | |
| sp | CF124 | WP_045557721.1 | - | NF | GCA_900114955.1 | |
| sp | FxanaA7 | WP_045557721.1 | - | WP_234442873.1 | GCA_000958545.1 | |
| turgidiscabies | T45 | WP_059073075.1 | - | WP_006378806.1 | GCA_001485145.1 | [35] |
| reticuliscabiei | NRRL B-24446 | WP_059073075.1 | - | WP_245879130.1 | GCF_002154675.1 | [47] |
| turgidiscabies | Car8 | ELP70267.1 | - | WP_006378806.1 | GCA_000331005.1 | [48] |
| reticuli | - | CAB46342.1 | - | - | GCA_001511815.1 | - |
| griseus | NBRC 13350 | WP_012379731.1 | - | - | [49] | |
| Primer | Sequence (5′→3′) * | Use |
|---|---|---|
| sc052 | cggggggtcggcacgtcagggcatcaggaggacgcaatgattccggggatccgtcgacc | Redirect deletion cassette for scab57751-scab57731 (cebEFG1) |
| imf271 | ccccggtgagaagacgtacggcggcgaaccggggcctcatgtaggctggagctgcttc | Redirect deletion cassette for scab57751-scab57731 (cebEFG1) |
| sc056 | cttacacatcccgcacgttccc | Verification of ∆scab57751-scab57731 (∆cebEFG1) |
| imf272 | gcgtgtgactgaaggtgtcc | Verification of ∆scab57751-scab57731 (∆cebEFG1) |
| sc050 | tccatgggcacgttcggttggtcgcacagaaaggcgatgattccggggatccgtcgacc | Redirect deletion cassette for scab2421-scab2401 (cebEFG2) |
| imf296 | ggcggggctcggctgtgcgggggcggtgggggcgggtcatgtaggctggagctgcttc | Redirect deletion cassette for scab2421-scab2401 (cebEFG2) |
| sc058 | cgttcggttggtcgcacagaaagg | Verification of ∆scab2421-scab2401 (∆cebEFG2) |
| imf297 | acgggctggatgttcttctc | Verification of ∆scab2421-scab2401 (∆cebEFG2) |
| mjk1 | gcaacatcgcggcacaccgcctggaaataaggagtacagatgattccggggatccgtcgacc | Redirect deletion cassette for scab77271 (pitE) |
| mjk2 | cgtccgcggagggcgcccccgagcgcgcggacgaaccgatcatgtaggctggagctgcttc | Redirect deletion cassette for scab77271 (pitE) |
| mjk3 | gccatgccttcaacatacag | Verification of ∆scab77271 (∆pitE) |
| mjk4 | gctggtcgaggataaaggtg | Verification of ∆scab77271 (∆pitE) |
Appendix B
Genomic DNA Extraction
References
- Wilson, D.B. Microbial diversity of cellulose hydrolysis. Curr. Opin. Microbiol. 2011, 14, 259–263. [Google Scholar] [CrossRef]
- Loria, R.; Kers, J.; Joshi, M. Evolution of plant pathogenicity in Streptomyces. Annu. Rev. Phytopathol. 2006, 44, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Wach, J.M.; Krasnoff, S.B.; Loria, R.; Gibson, D.M. Effect of carbohydrates on the production of thaxtomin A by Streptomyces scabies. Arch. Microbiol. 2007, 18, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.G.; Joshi, M.V.; Gibson, D.M.; Loria, R. Cello-oligosaccharides released from host plants induce pathogenicity in scab-causing Streptomyces species. Physiol. Mol. Plant Pathol. 2007, 71, 18–25. [Google Scholar] [CrossRef]
- Bischoff, V.; Cookson, S.J.; Wu, S.; Scheible, W.R. Thaxtomin A affects CESA-complex density, expression of cell wall genes, cell wall composition, and causes ectopic lignification in Arabidopsis thaliana seedlings. J. Exp. Bot. 2009, 60, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.R.; Fry, B.; Kochevenko, A.; Schindelasch, D.; Zimmerli, L.; Somerville, S.; Loria, R.; Somerville, C.R. An Arabidopsis mutant resistant to thaxtomin A, a cellulose synthesis inhibitor from Streptomyces species. Plant Cell 2003, 15, 1781–1794. [Google Scholar] [CrossRef]
- Deflandre, B.; Stulanovic, N.; Planckaert, S.; Anderssen, S.; Bonometti, B.; Krim, L.; Coppieters, W.; Devreese, B.; Rigali, S. The virulome of Streptomyces scabiei in response to cello-olligasaccharides. Microb. Genom. 2022, 8, 000760. [Google Scholar] [CrossRef]
- Jourdan, S.; Francis, I.M.; Deflandre, B.; Loria, R.; Rigali, S. Tracking the subtle mutations driving host sensing by the plant pathogen Streptomyces scabies. mSphere 2017, 2, 27144. [Google Scholar] [CrossRef]
- Jourdan, S.; Francis, I.M.; Kim, M.J.; Salazar, J.J.C.; Planckaert, S.; Frère, J.M.; Matagne, A.; Kerff, F.; Devreese, B.; Loria, R.; et al. The CebE/MsiK transporter is a doorway to the cello-oligosaccharide-mediated induction of Streptomyces scabies pathogenicity. Sci. Rep. 2016, 2, e00367-16. [Google Scholar] [CrossRef]
- Jourdan, S.; Francis, I.M.; Deflandre, B.; Tenconi, E.; Riley, J.; Planckaert, S.; Tocquin, P.; Martinet, L.; Devreese, B.; Loria, R.; et al. Contribution of the β-glucosidase BglC to the onset of the pathogenic lifestyle of Streptomyces scabies. Mol. Plant Pathol. 2018, 19, 1480–1490. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.V.; Bignell, D.R.D.; Johnson, E.G.; Sparks, J.P.; Gibson, D.M.; Loria, R. The AraC/XylS regulator TxtR modulates thaxtomin biosynthesis and virulence in Streptomyces scabies. Mol. Microbiol. 2007, 66, 633–642. [Google Scholar] [CrossRef]
- Francis, I.M.; Jourdan, S.; Fanara, S.; Loria, R.; Rigali, S. The cellobiose sensor CebR is the gatekeeper of Streptomyces scabies pathogenicity. mBio 2015, 6, e02018. [Google Scholar] [CrossRef]
- Deflandre, B.; Thiébaut, N.; Planckaert, S.; Jourdan, S.; Anderssen, S.; Hanikenne, M.; Devreese, B.; Francis, I.; Rigali, S. Deletion of bglC triggers a genetic compensation response by awakening the expression of alternative beta-glucosidase. BBA Gene Regul. Mech. 2020, 1863, 194615. [Google Scholar] [CrossRef] [PubMed]
- Lerat, S.; Simao-Beaunoir, A.M.; Wu, R.; Beaudoin, N.; Beaulieu, C. Involvement of the plant polymer suberin and the disaccharide cellobiose in triggering thaxtomin A biosynthesis, a phytotoxin produced by the pathogenic agent Streptomyces scabies. Phytopathology 2010, 100, 91–96. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [PubMed]
- Hiard, S.; Marée, R.; Colson, S.; Hoskisson, P.; Titgemeyer, F.; van Wezel, G.V.; Joris, B.; Wehenkel, L.; Rigali, S. PREDetector: A new tool to identify regulatory elements in bacterial genomes. Biochem. Biophys. Res. Commun. 2007, 357, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Anderssen, S.; Naômé, A.; Jadot, C.; Brans, A.; Tocquin, P.; Rigali, S. AURTHO: Autoregulation of transcription factors as facilitator of cis-acting element discovery. BBA Gene Regul. Mech. 2022, 1865, 194847. [Google Scholar] [CrossRef] [PubMed]
- Gust, B.; Challis, G.L.; Fowler, K.; Kieser, T.; Chater, K.F. PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. USA 2003, 100, 1541–1546. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Schlösser, A.; Jantos, J.; Hackmann, K.; Schrempf, H. Characterization of the binding protein-dependent cellobiose and cellotriose transport system of the cellulose degrader Streptomyces reticuli. Appl. Environ. Microbiol. 1999, 65, 2636–2643. [Google Scholar] [CrossRef] [PubMed]
- Marushima, K.; Ohnishi, Y.; Horinouchi, S. CebR as a master regulator for cellulose/cellooligosaccharide catabolism affects morphological development in Streptomyces griseus. J. Bacteriol. 2009, 191, 5930. [Google Scholar] [CrossRef] [PubMed]
- Bukhalid, R.A.; Takeuchi, T.; Labeda, D.; Loria, R. Horizontal transfer of the plant virulence gene, nec1, and flanking sequences among genetically distinct Streptomyces strains in the Diastatochromogenes cluster. Appl. Environ. Microbiol. 2002, 68, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Schlösser, A.; Aldekamp, T.; Schrempf, H. Binding characteristics of CebR, the regulator of the ceb operon required for cellobiose/cellotriose uptake in Streptomyces reticuli. FEMS Microbiol. Lett. 2000, 190, 127–132. [Google Scholar] [CrossRef]
- Book, A.J.; Lewin, G.R.; McDonald, B.R.; Takasuka, T.E.; Doering, D.T.; Adams, A.S.; Blodgett, J.A.; Clardy, J.; Raffa, K.F.; Fox, B.G.; et al. Cellulolytic Streptomyces strains associated with herbivorous insects share a phylogenetically linked capacity to degrade lignocellulose. Appl. Environ. Microbiol. 2014, 80, 4692–4701. [Google Scholar] [CrossRef]
- Bertram, R.; Schlicht, M.; Mahr, K.; Nothaft, H.; Saier, M.H.; Titgemeyer, F. In silico and transcriptional analysis of carbohydrate uptake systems of Streptomyces coelicolor A3(2). J. Bacteriol. 2004, 186, 1362–1373. [Google Scholar] [CrossRef]
- Hurtubise, Y.; Shareck, F.; Kluepfel, D.; Morosoli, R. A cellulase/xylanase-negative mutant of Streptomyces lividans 1326 defective in cellobiose and xylobiose uptake is mutated in a gene encoding a protein homologous to ATP-binding proteins. Mol. Microbiol. 1995, 17, 367–377. [Google Scholar] [CrossRef]
- MacNeil, D.J.; Gewain, K.M.; Ruby, C.L.; Dezeny, G.; Gibbons, P.H.; MacNeil, T. Analysis of Streptomyces avermitilis genes required for avermectin biosynthesis utilizing a novel integrative vector. Gene 1992, 111, 61–68. [Google Scholar] [CrossRef]
- Loria, R.; Bukhalid, R.A.; Creath, R.A.; Leiner, R.H.; Olivier, M.; Steffens, J.C. Differential production of thaxtomins by pathogenic Streptomyces species in vitro. Phytopathology 1995, 85, 537–541. [Google Scholar] [CrossRef]
- Bignell, D.R.D.; Huguet-Tapia, J.C.; Joshi, M.V.; Pettis, G.S.; Loria, R. What does it take to be a plant pathogen: Genomic insights from Streptomyces species. Antonie Van Leeuwenhoek 2010, 98, 179–194. [Google Scholar] [CrossRef]
- Bignell, D.R.D.; Seipke, R.F.; Huguet-Tapia, J.C.; Chambers, A.H.; Parry, R.J.; Loria, R. Streptomyces scabies 87-22 contains a coronafacic acid-like biosynthetic cluster that contributes to plant-microbe interactions. Mol. Plant Microbe Interact. 2010, 23, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Hudec, C.; Biessy, A.; Novinscak, A.; St-Onge, R.J.; Lamarre, S.; Blom, J.; Fillion, M. Comparative genomics of potato common scab-causing Streptomyces spp. displaying varying virulence. Front. Microbiol. 2021, 12, 716522. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bignell, D.R.D.; Zuo, R.; Fan, Q.; Huguet-Tapia, J.C.; Ding, Y.; Loria, R. Promiscuous pathogenicity islands and phylogeny of pathogenic Streptomyces spp. Mol. Plant Microb. Interact. 2016, 29, 640–650. [Google Scholar] [CrossRef]
- Labeda, D.P.; Goodfellow, M.; Brown, R.; Ward, A.C.; Lanoot, B.; Vanncanneyt, M.; Swings, J.; Kim, S.B.; Liu, Z.; Chun, J.; et al. Phylogenetic study of the species within the family Streptomycetaceae. Antonie Van Leeuwenhoek 2012, 101, 73–104. [Google Scholar] [CrossRef]
- Tomihama, T.; Nishi, Y.; Sakai, M.; Ikenaga, M.; Okubo, T.; Ikeda, S. Draft genome sequences of Streptomyces scabiei S58, Streptomyces turgidiscabies T45, and Streptomyces acidiscabies a10, the pathogens of potato common scab, isolated in Japan. Genome Announc. 2016, 4, e00062-16. [Google Scholar] [CrossRef] [PubMed]
- Croce, V.; López-Radcenco, A.; Lapaz, M.I.; Pianzzola, M.J.; Moyna, G.; Siri, M.I. An integrative approach for the characterization of plant-pathogenic Streptomyces spp. strains based on metabolomic, bioactivity, and phylogenetic analysis. Front. Microbiol. 2021, 12, 643792. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, D.B.A.; do Amaral, D.T.; da Silva, M.J.; Destéfano, S.A.L. Sterptomyces brasiliscabiei, a new species causing potato scab in south Brazil. Antonie Van Leeuwenhoek 2021, 114, 913–931. [Google Scholar] [CrossRef]
- USDA Agricultural Research Service. Whole-Genome Sequencing NRRL B-16521 and NRRL B-2795. National Center for Biotechnology Information. 2022. Available online: https://data.nal.usda.gov/dataset/whole-genome-sequencing-nrrl-b-16521-and-nrrl-b-2795 (accessed on 30 December 2022).
- Clark, C.; Chen, C.; Ward-Rainey, N.; Pettis, G.S. Diversity within Streptomyces ipomoeae based on inhibitory interactions, rep-PCR, and plasmid profiles. Phytopathology 1998, 88, 1179–1186. [Google Scholar] [CrossRef]
- Huguet-Tapia, J.C.; Lefébure, T.; Badger, J.H.; Guan, D.; Pettis, G.S.; Stanhope, M.J.; Loria, R. Genome content and phylogenomics reveal both ancestral and lateral evolutionary pathways in plant pathogenic Streptomyces species. Appl. Environ. Microbiol. 2016, 82, 2146–2155. [Google Scholar] [CrossRef]
- USDA/ARS/NCAUR. Streptomyces Europaeiscabiei Strain: NRRL B-24443 Genome Sequencing and Assembly. National Center for Biotechnology Information. 2022. Available online: https://data.nal.usda.gov/dataset/streptomyces-europaeiscabiei-strainnrrl-b-24443-genome-sequencing-and-assembly (accessed on 30 December 2022).
- USDA/ARS/NCAUR. Streptomyces Stelliscabiei Strain: NRRL B-24447 Genome Sequencing and Assembly. National Center for Biotechnology Information. 2022. Available online: https://data.nal.usda.gov/dataset/streptomyces-stelliscabiei-strainnrrl-b-24447-genome-sequencing-and-assembly (accessed on 30 December 2022).
- Huguet-Tapia, J.C.; Loria, R. Draft genome sequence of Streptomyces acidiscabies 84-104, an emergent plant pathogen. J. Bacteriol. 2012, 194, 1847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Loria, R. Emergence of novel pathogenic Streptomyces by site-specific accretion and cis-mobilization of pathogenicity islands. Mol. Plant Microbe Interact. 2017, 30, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Park, D.H.; Kim, J.S.; Kwon, S.W.; Wilson, C.; Yu, Y.M.; Hur, J.H.; Lim, C.K. Streptomyces luridiscabiei sp. nov. Streptomyces puniciscabiei sp. nov. and Streptomyces niveiscabiei sp. nov. which cause potato common scab disease in Korea. Int. J. Syst. Evol. Microbiol. 2003, 53, 2049–2054. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Weisberg, A.J.; Chang, J.H.; Clarke, C.R. Streptomyces caniscabiei sp. nov., which causes potato common scab and is distributed across the world. Int. J. Syst. Evol. Microbiol. 2022, 72, 005225. [Google Scholar] [CrossRef] [PubMed]
- Bouchek-Mechiche, K.; Gardan, L.; Normand, P.; Jouan, B. DNA relatedness among strains of Streptomyces pathogenic to potato in France: Description of three new species, S. europaeiscabiei sp. nov. and S. stelliscabiei sp. nov. associated with common scab, and S. reticuliscabiei sp. nov. associated with netted scab. Int. J. Syst. Evol. Microbiol. 2000, 50, 91–99. [Google Scholar] [CrossRef]
- Huguet-Tapia, J.C.; Badger, J.H.; Loria, R.; Pettis, G.S. Streptomyces turgidiscabies Car8 contains a modular pathogenicity island that shares virulence genes with other actinobacterial plant pathogens. Plasmid 2011, 65, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, Y.; Ishikawa, J.; Hara, H.; Suzuki, H.; Ikenoya, M.; Ikeda, H.; Yamashita, A.; Hattori, M.; Horinouchi, S. Genome sequence of the streptomycin-producing microorganism Streptomyces griseus IFO 13350. J. Bacteriol. 2008, 190, 4050–4060. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francis, I.M.; Bergin, D.; Deflandre, B.; Gupta, S.; Salazar, J.J.C.; Villagrana, R.; Stulanovic, N.; Ribeiro Monteiro, S.; Kerff, F.; Loria, R.; et al. Role of Alternative Elicitor Transporters in the Onset of Plant Host Colonization by Streptomyces scabiei 87-22. Biology 2023, 12, 234. https://doi.org/10.3390/biology12020234
Francis IM, Bergin D, Deflandre B, Gupta S, Salazar JJC, Villagrana R, Stulanovic N, Ribeiro Monteiro S, Kerff F, Loria R, et al. Role of Alternative Elicitor Transporters in the Onset of Plant Host Colonization by Streptomyces scabiei 87-22. Biology. 2023; 12(2):234. https://doi.org/10.3390/biology12020234
Chicago/Turabian StyleFrancis, Isolde M., Danica Bergin, Benoit Deflandre, Sagar Gupta, Joren J. C. Salazar, Richard Villagrana, Nudzejma Stulanovic, Silvia Ribeiro Monteiro, Frédéric Kerff, Rosemary Loria, and et al. 2023. "Role of Alternative Elicitor Transporters in the Onset of Plant Host Colonization by Streptomyces scabiei 87-22" Biology 12, no. 2: 234. https://doi.org/10.3390/biology12020234
APA StyleFrancis, I. M., Bergin, D., Deflandre, B., Gupta, S., Salazar, J. J. C., Villagrana, R., Stulanovic, N., Ribeiro Monteiro, S., Kerff, F., Loria, R., & Rigali, S. (2023). Role of Alternative Elicitor Transporters in the Onset of Plant Host Colonization by Streptomyces scabiei 87-22. Biology, 12(2), 234. https://doi.org/10.3390/biology12020234