


The Ecological Status and Change in High-Latitude Coral Assemblages at the Xuwen Coral Reef, Northern South China Sea: Insight into the Status and Causes in 2020

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
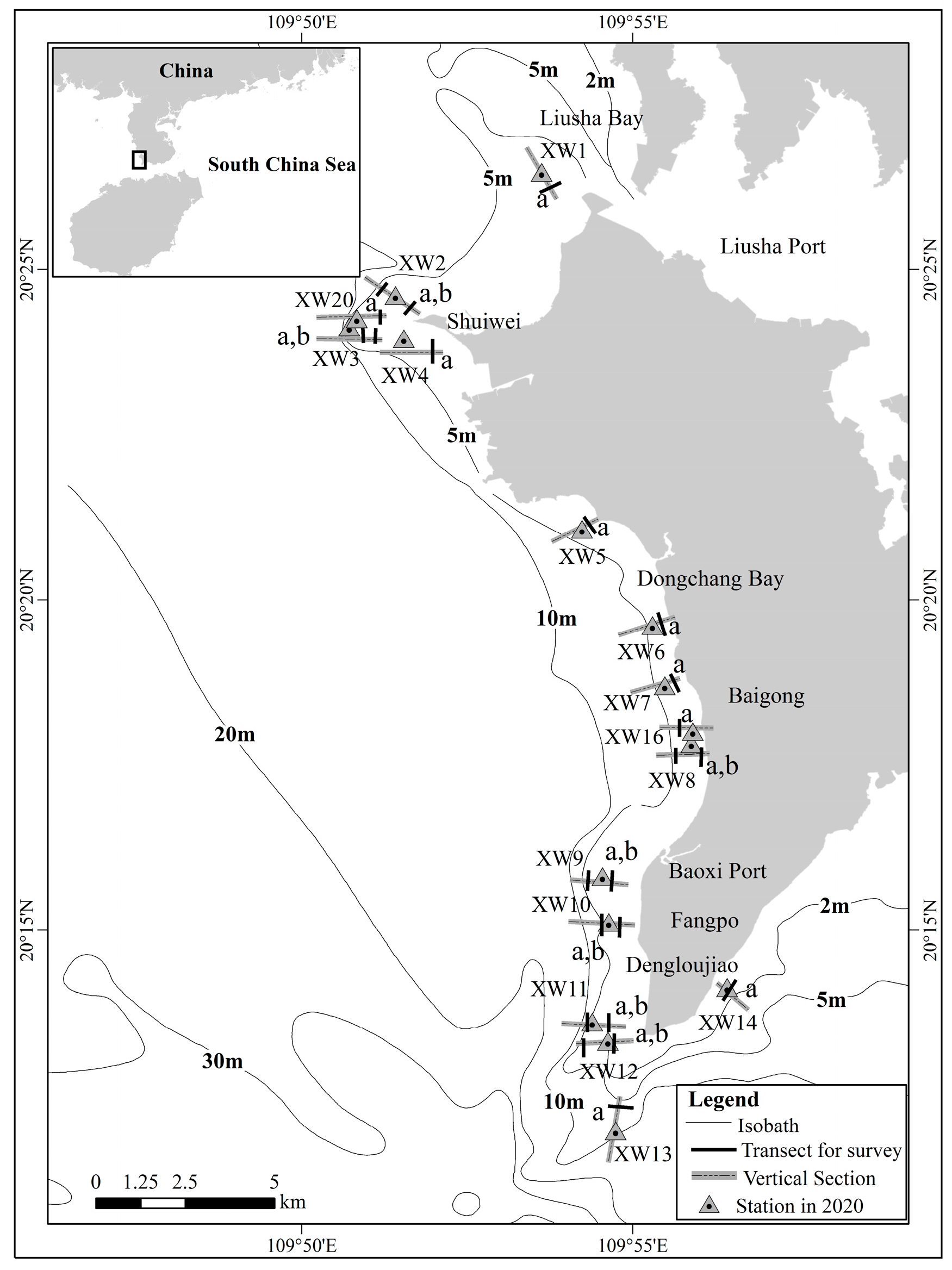
2.1. Field Surveys
2.2. Data Extraction and Treatment
3. Results
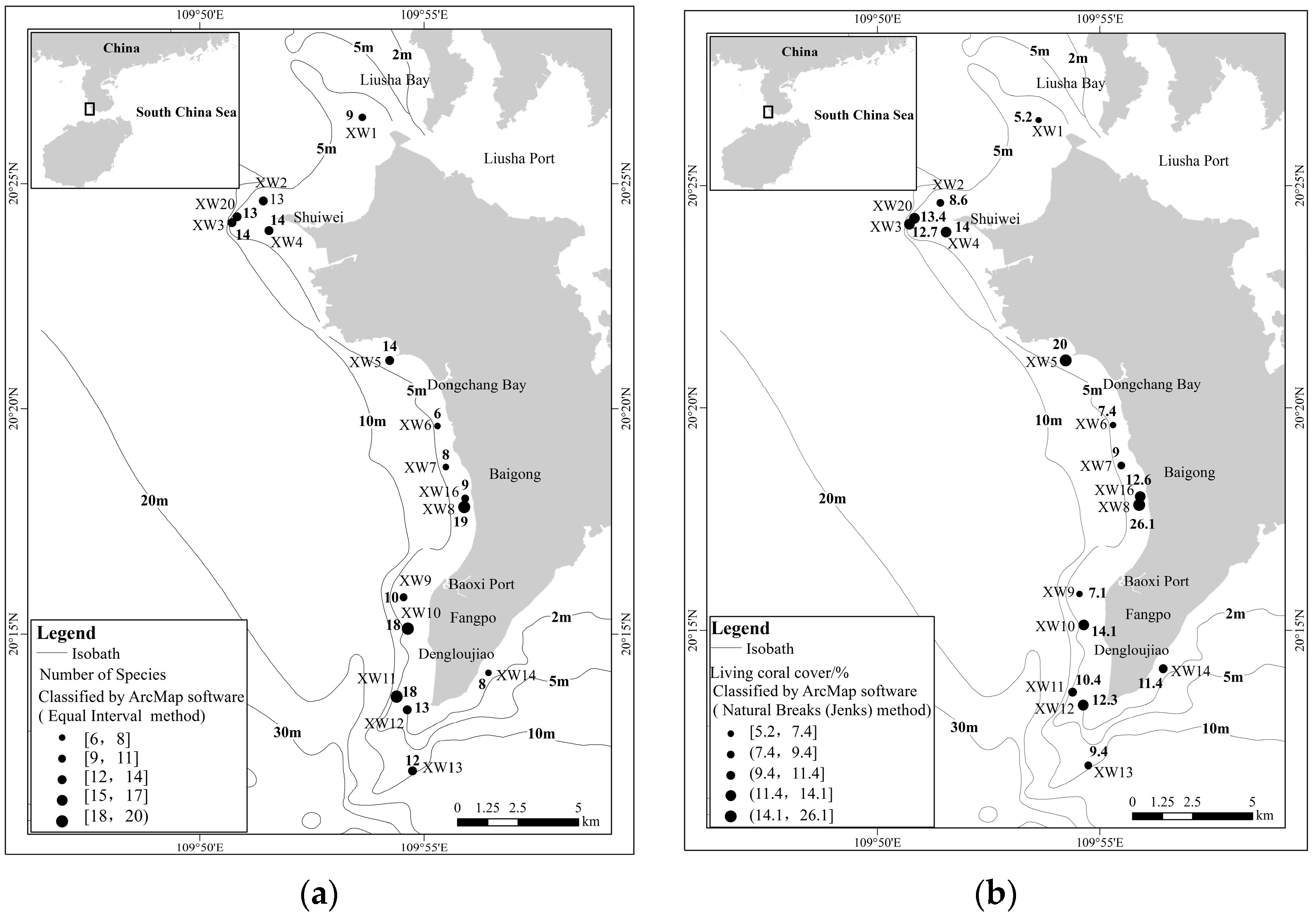
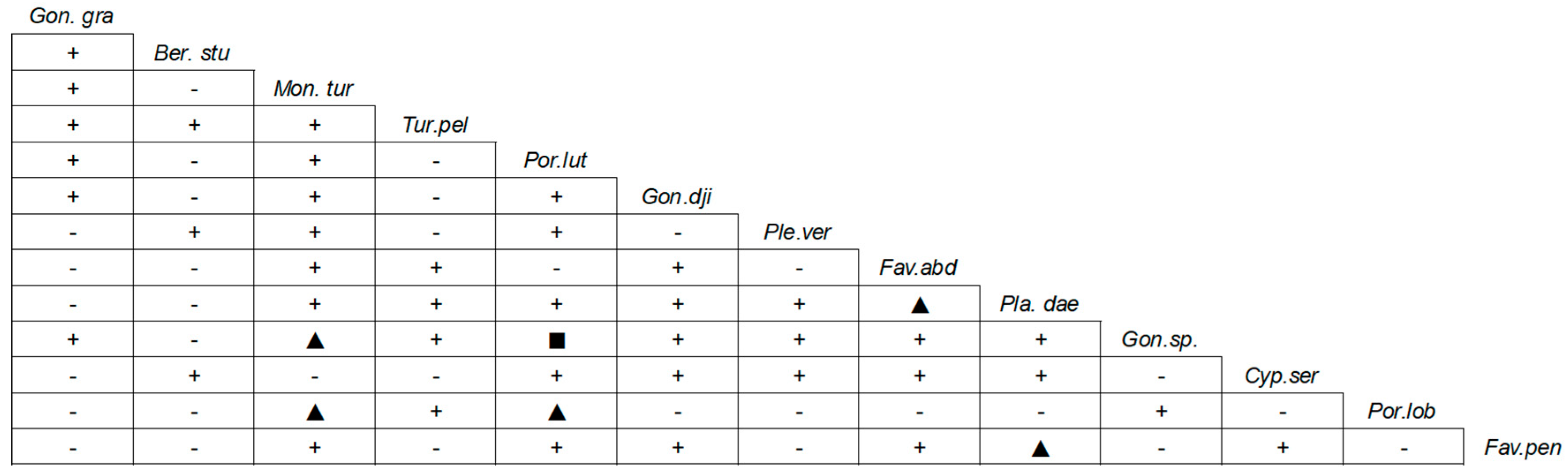
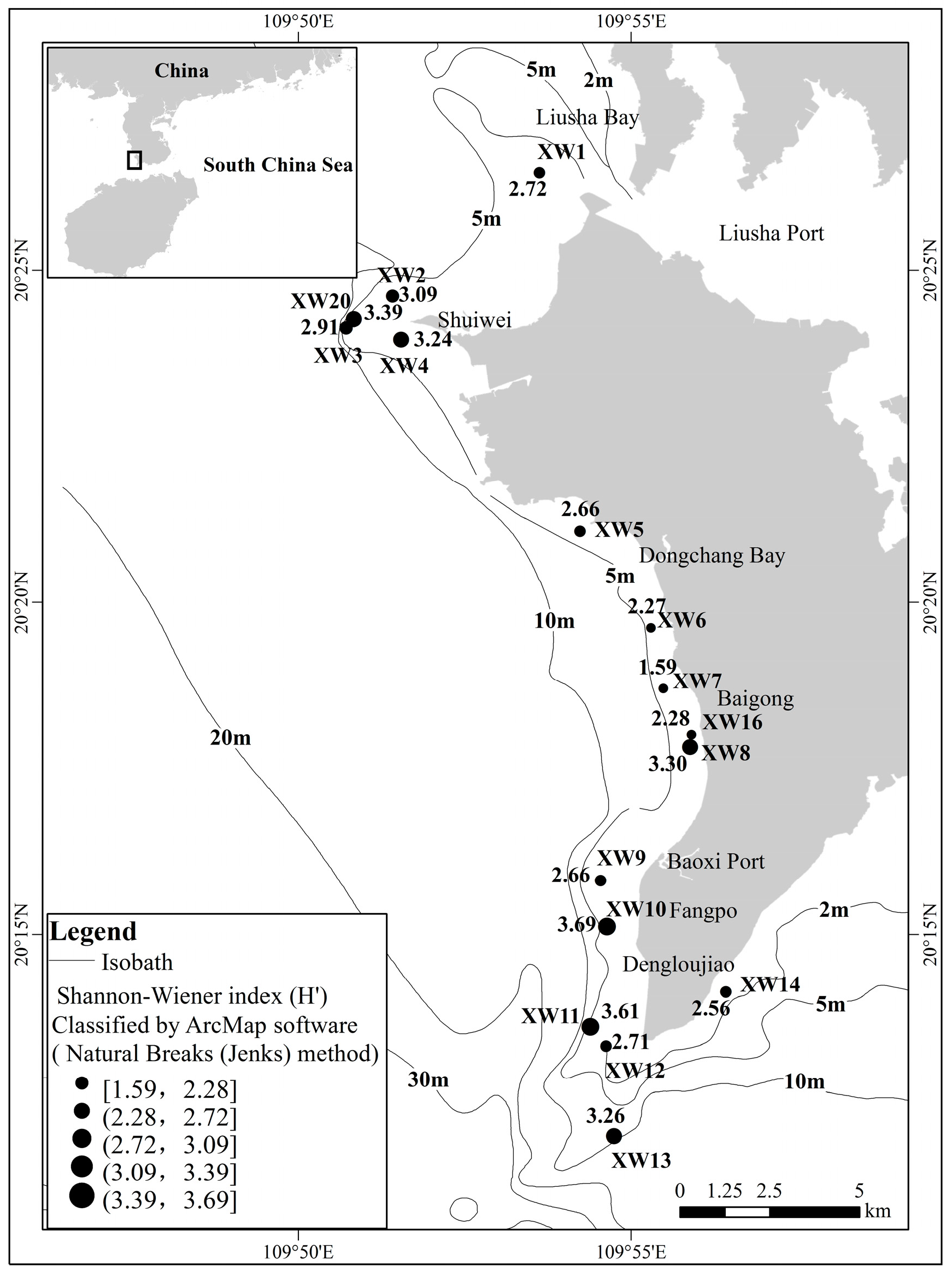
3.1. Coral Community Structure
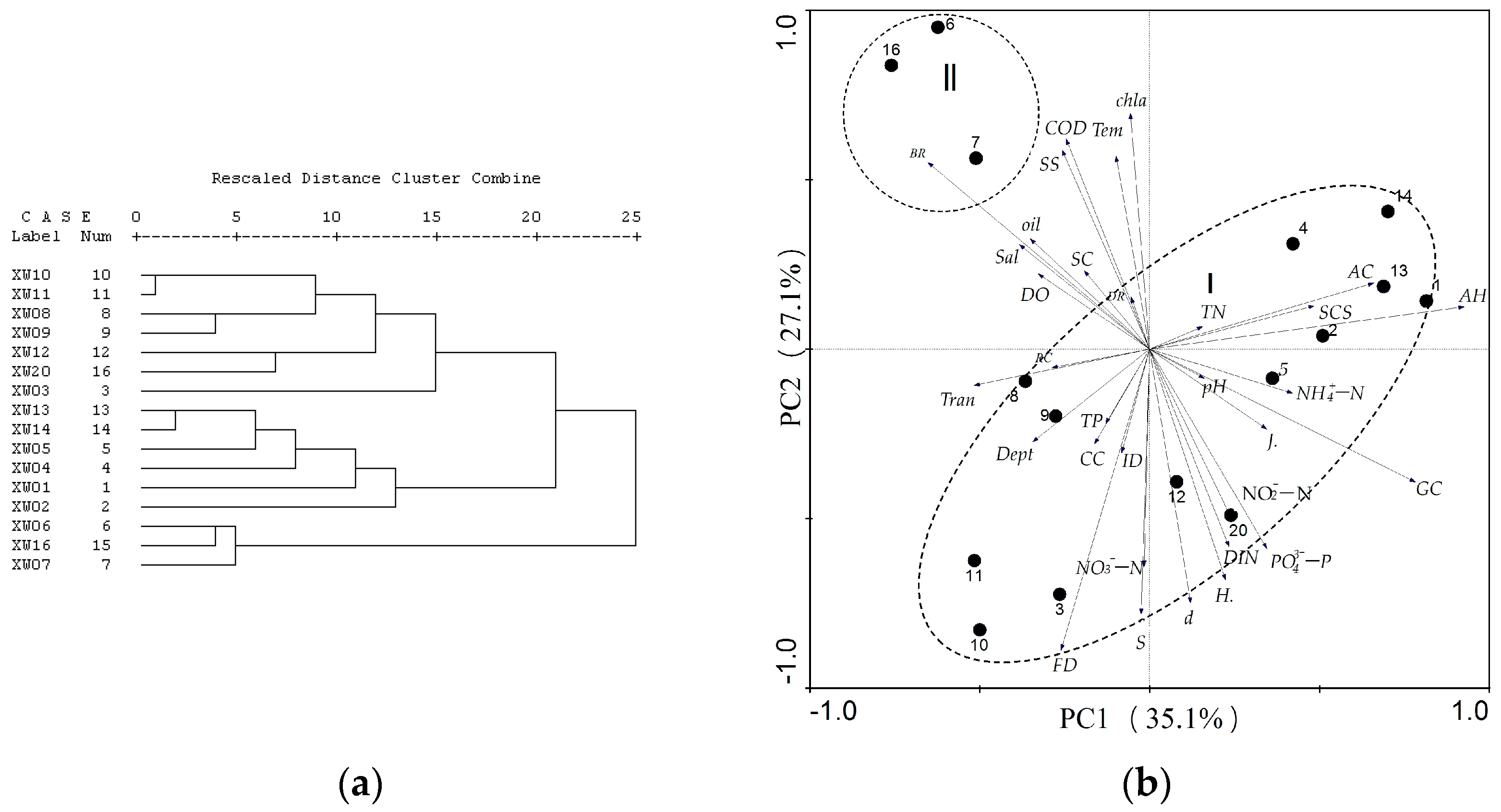
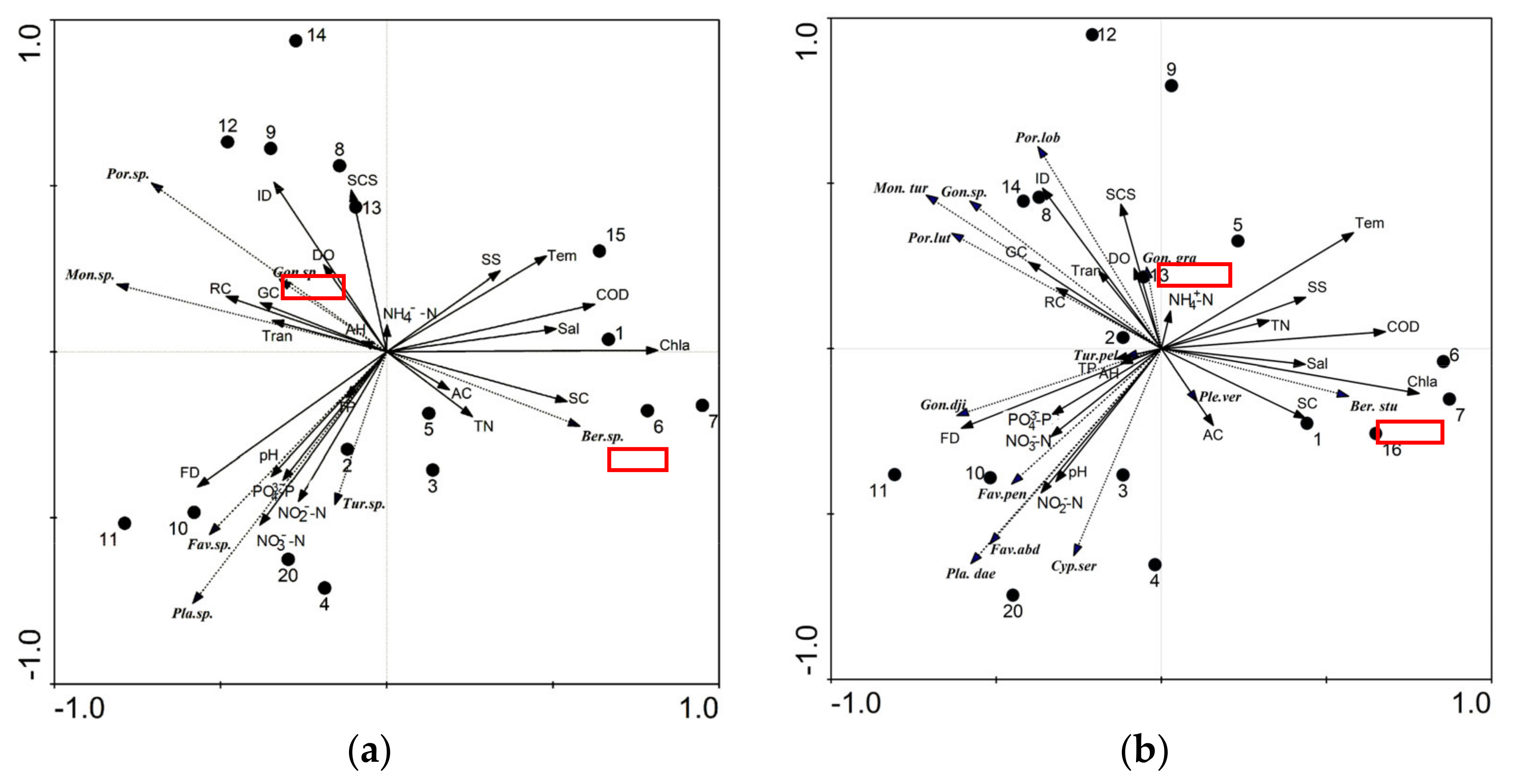
3.2. Reef Dwelling Organism Composition and Environmental Factors in Water
3.3. Water Environment Factors Affecting Coral Growth
4. Discussion
4.1. Temporal Variations in the Coral Community on the Xuwen Coral Reef
4.2. Global Warming Affecting the Xuwen Coral Reef
4.3. Escalating Anthropogenic Activities Weakening the Coral Habitat Function on the Xuwen Coral Reef
4.4. Factors Influencing Coral Community Structure
4.4.1. High Suspended Solids on the Xuwen Coral Reef
4.4.2. Elevated Nutrient Loads Affect Coral Growth and Reproduction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nie, B.F.; Chen, T.G.; Liang, M.T.; Zhong, J.L.; Yu, K.F. Coral reefs of Leizhou Peninsula and holocene high sea level. Chin. Sci. Bull. 1997, 42, 511–514. [Google Scholar]
- Zhao, J.X.; Yu, K.F. Mass spectrometry of reef building corals in Leizhou Peninsula, South China Sea. Uranium age and holocene high sea level. Chin. Sci. Bull. 2001, 46, 1734–1738. [Google Scholar]
- Zhao, H.T.; Wang, L.R.; Song, C.J.; Yu, K.F.; Yuan, J.Y. Features of fringing reef at Dengloujiao, Leizhou Peninsula. Mar. Geol. Quat. Geol. 2002, 22, 35–40. [Google Scholar]
- Zhao, H.T.; Wang, L.R.; Song, C.J. The distribution and protection of coral reefs in the west of Xuwen County. Trop. Geo. 2006, 26, 202–206. [Google Scholar]
- Huang, H.; Lian, J.S.; Wang, H.J.; Chen, Y.H. Xuwen Coral Reefs and their Biodiversity; Ocean Press: Beijing, China, 2007. [Google Scholar]
- Chen, T.R.; Li, S.b.; Yu, K.F.; Zheng, Z.Y.; Wang, L.R.; Chen, T.G. Increasing temperature anomalies reduce coral growth in the Weizhou Island, northern South China Sea. Estuar. Coast. Shelf Sci. 2013, 130, 121–126. [Google Scholar] [CrossRef]
- Barber, R.T.; Hilting, A.K.; Hayes, M.L. The changing health of coral reefs. Hum. Ecol. Risk Assess. Int. J. 2001, 7, 1255–1270. [Google Scholar] [CrossRef]
- Chen, T.R.; Zheng, Z.Y.; Mo, S.H.; Zhou, X.; Chen, T.G. Variation of skeletal extension rate for Porites corals around Weizhou Island in response to global warming and increase of extreme events. J. Trop. Oceanogr. 2013, 32, 79–84. [Google Scholar] [CrossRef]
- Wang, L.R.; Chen, R.Q.; Zhao, H.T. A preliminary study on coral-associated communities in the Xuwen Coral Reef conservation. Mar. Sci. 2008, 32, 56–62. [Google Scholar]
- Wang, L.R.; Chen, R.Q.; Zhao, H.T. Organism in intertidal zone of fringing reef at Qiongzhou Straits. J. Oceanogr. Taiwan Strait. 2003, 22, 286–294. [Google Scholar]
- Lu, H.S.; He, X.L.; Chen, C.L.; Xuan, L.Q. Species and distribution of “corals” in the coastal sea area of Western Xuwen, Guangdong. J. Oceanogr. Taiwan Strait. 2003, 22, 445–448. [Google Scholar]
- Zhao, H.T.; Wang, L.R.; Song, C.J.; Fang, J.W.; Denise, M.C.; Huang, Z.J. Anthozoa species in western littoral waters of Xuwen County, Guangdong Province. J. Oceanogr. Taiwan Strait. 2009, 28, 405–409. [Google Scholar]
- Liao, B.L.; Liu, L.; Liu, C.Y. The research status and prospect in Xuwen coral reefs. J. Guangdong Ocean Univ. 2011, 8, 91–96. [Google Scholar]
- Huang, H.; Zhang, Y.Y.; Lian, J.S.; Li, X.B.; You, F.; Yang, J.H.; Lei, X.M.; Zhang, C.L. Structure and diversity of scleractinia coral communities along the west seashore of Xuwen County. Biodiversity Sci. 2011, 19, 505–510. [Google Scholar]
- Marine and Fishery Bureau of Guangdong Province. Bulletin on Marine Environment of Guangdong Province in 2015. Guangzhou, China, 2016. Available online: http://gdee.gd.gov.cn/attachment/0/484/484320/2466222.pdf (accessed on 29 December 2022).
- Li, F.; Shen, C.; Zhang, Y.P.; Zhou, J.; Peng, H.P.; Liu, L. Analysis of coral species and coverage in Xuwen Coral Reef National Nature Reserve in Guangdong. Jiangsu Agric. Sci. 2019, 47, 304–308. [Google Scholar] [CrossRef]
- Hughes, T.P.; Graham, N.A.; Jackson, J.B.; Mumby, P.J.; Steneck, R.S. Rising to the challenge of sustaining coral reef resilience. Trends. Ecol. Evol. 2010, 25, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.F. Coral reefs in the South China Sea: Their response to and records on past environmental changes. Sci. China. Earth. Sci. 2012, 55, 1217–1229. [Google Scholar] [CrossRef]
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Nat. Acad. Sci. USA. 2012, 109, 17995–17999. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.R.; Yu, K.F.; Shi, Q.; Li, S.; Price, G.J.; Wang, L.R.; Zhao, M.X.; Chen, T.G.; Zhao, J. Twenty-five years of change in scleractinian coral communities of Daya Bay (northern South China Sea) and its response to the 2008 AD extreme cold climate event. Chin Sci Bull. 2009, 54, 2107–2117. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Huang, X.P.; Yue, W.Z.; Lin, Y.T.; Zou, R.L.; Huang, H. Ecological study on coral reef and intertidal benthos around yongxing island, south China sea. Oceanol. Limnol. Sin. 2004, 35, 176–182. [Google Scholar]
- Shi, Q.; Yan, H.; Zhang, H.; Zhao, M. Spatial variations of stony coral cover on the reef slope of Yongxing Island, Xisha Islands. J. Trop. Oceanogr. 2011, 30, 10–17. [Google Scholar]
- Pandolfi, J.M.; Roger, H.B.; Enric, S.; Terence, P.H.; Karen, A.B.; Richard, G.C.; Deborah, M.; Loren, M.; Marah, J.H.N.; Gustavo, P.; et al. Global trajectories of the long-term decline of coral reef ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.H.; Yu, K.F.; Wang, Y.H. A Review on the research of coral reefs in weizhou Island, Beibu Gulf. Trop. Geo. 2016, 36, 72–79. [Google Scholar] [CrossRef]
- Lybolt, M.; Neil, D.; Zhao, J.X.; Feng, Y.X.; Yu, K.F.; Pandolfi, J. Instability in a marginal coral reef: The shift from natural variability to a human-dominated seascape. Front. Ecol. Environ. 2011, 9, 154–160. [Google Scholar] [CrossRef]
- Zhang, R.J.; Zhang, R.L.; Yu, K.F.; Wang, Y.H.; Huang, X.Y.; Pe, J.Y.; Wei, C.S.; Pan, Z.L.; Qin, Z.J.; Zhang, G. Occurrence, sources and transport of antibiotics in the surface water of coral reef regions in the South China Sea: Potential risk to coral growth. Environ. Pollut. 2018, 232, 450–457. [Google Scholar] [CrossRef]
- National Meteorological Centre. Xuwen Weather Forecast. Available online: http://www.nmc.cn/publish/forecast/AGD/xuwen.html (accessed on 29 December 2022).
- Hill, J.; Wilkinson, C. Methods for Ecological Monitoring of Coral Reefs; Australian Institute of Marine Science: Townsville, Australian, 2004. [Google Scholar]
- Foster, M.S.; Harrold, C.; Hardin, D.D. Point vs. photo quadrat estimates of the cover of sessile marine organisms. J. Exp. Mar. Biol. Ecol. 1991, 146, 193–203. [Google Scholar] [CrossRef]
- Veron, J.E.N. Corals of the World; Australian Institute of Marine Science: Townsville, Australian, 2000. [Google Scholar]
- Zou, R.L. Fauna Sinica-Hermatypic Coral; Science Press: Beijing, China, 2001. [Google Scholar]
- Zhang, Q.M.; Shi, Q.; Chen, G.; Fong, T.C.; Wong, D.C.; Huang, H.; Wang, H.K.; Zhao, M.X. Status monitoring and health assessment of Luhuitou fringing reef of Sanya, Hainan, China. Chin. Sci. Bull. 2006, 51, 81–88. [Google Scholar] [CrossRef]
- English, S.; Wilkinson, C.; Baker, V. Sumanual Rvey for Tropical Marine Resources; Australian Institute Marine Science: Townsville, Australian, 1997. [Google Scholar]
- Standardization Administration of China. Specification for Oceanographic Survey (GB 12763-2007); Ocean Press: Beijing, China, 2007. [Google Scholar]
- Standardization Administration of China. Specification for Marine Monitoring (GB 17378-2007); Ocean Press: Beijing, China, 2007. [Google Scholar]
- Gj, M. Ethnobotany: A Methods Manual; Chapman and Hall: London, UK, 1995. [Google Scholar]
- Melese, S.M.; Ayele, B. Woody plant diversity, structure and regeneration in the Ambo State Forest, South Gondar Zone, Northwest Ethiopia. J. For. Res. 2017, 28, 133–144. [Google Scholar] [CrossRef]
- Toksha, B.G.; Shirsath, S.E.; Patange, S.M.; Jadhav, K.M. Structural investigations and magnetic properties of cobalt ferrite nanoparticles prepared by sol-gel auto combustion method. Solid. State. Commun. 2008, 147, 479–483. [Google Scholar] [CrossRef]
- Yu, W.J.; Wang, W.H.; Yu, K.F.; Wang, Y.H.; Huang, X.Y.; Huang, R.Y.; Liao, Z.H.; Xu, S.D.; Chen, X.Y. Rapid decline of a relatively high latitude coral assemblage at Weizhou Island, northern South China Sea. Biodiversity Conserv. 2019, 28, 3925–3949. [Google Scholar] [CrossRef]
- Liang, W.; Li, G.Z.; Fan, H.Q.; Nong, H.Q.; Wang, X. Study on biodiversity of coral reef species in Weizhou Island, Guangxi. Mar. Sci. Bull. 2010, 29, 412–416. [Google Scholar]
- Sournia, A. Phytoplankton Manual//Sournia A, ed. Monographs on Oceanographic Methodology 6; UNESCO: Pairs, UK, 1978; pp. 251–260. [Google Scholar]
- Ren, J.Z. Research Methods of Grassland Science; China Agricultural Press: Beijing, China, 1998; pp. 45–56. [Google Scholar]
- Mehmood, A.; Shah, A.H.; Shah, A.H.; Khan, S.U.; Khan, K.R.; Farooq, M.; Ahmad, H.; Sakhi, S. Classification and ordination analysis of herbaceous flora in district Tor Ghar, western Himalaya. Acta. Ecol. Sin. 2021, 41, 451–462. [Google Scholar] [CrossRef]
- Hughes, T.P.; Huang, H.; Young, M.A. The wicked problem of China’s disappearing coral reefs. Conserv Biol. 2013, 27, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.X.; Yu, K.F.; Zhang, Q.; Shi, Q. Long-term change in coral cover in Luhuitou fringing reef, Sanya. Oceanol. Limnol. Sin. 2010, 41, 440–447. [Google Scholar]
- Darling, E.S.; McClanahan, T.R.; Côté, I.M. Combined effects of two stressors on Kenyan coral reefs are additive or antagonistic, not synergistic. Conserv. Lett. 2010, 3, 122–130. [Google Scholar] [CrossRef]
- Zhao, H.T.; Wang, L.R.; Song, C.J. conditions for the existence and development of coral reefs on the West Bank of Xuwen County. Trop. Geo. 2008, 5, 234–241. [Google Scholar] [CrossRef]
- Yu, K.F.; Liu, D.S.; Shen, C.D.; Zhao, J.X.; Chen, T.G.; Zhong, J.L.; Zhao, H.T.; Song, C.J. Environmental mutation events revealed by coral growth during Holocene high temperature period in Leizhou Peninsula. Sci. China Ser. D Earth Sci. 2002, 32, 149–156. [Google Scholar]
- Qin, Z.J.; Yu, K.F.; Liang, Y.T.; Chen, B.; Huang, X.Y. Latitudinal variation in reef coral tissue thickness in the South China Sea: Potential linkage with coral tolerance to environmental stress. Sci. Total Environ. 2020, 711, 134610. [Google Scholar] [CrossRef]
- Zhao, M.; Xu, W.B.; Kong, Y.Y.; Wang, X.Z.; Yu, Y.W. Research on the Vegetation Composition and Community Stability of Arid Grassland in the East of Qilian Mountains. Acta Bot. Boreali Occident. Sin. 2017, 37, 1847–1853. [Google Scholar]
- Sancia, E.T.; Suharsono; Hoeksema, B.W. Long-term changes in coral assemblages under natural and anthropogenic stress in Jakarta Bay (1920–2005). Mar. Pollut. Bull. 2010, 60, 1442–1454. [Google Scholar] [CrossRef] [Green Version]
- Bak, R.P.M.; Nieuwland, G. Long-term change in coral communities along depth gradients over leeward reefs in the Netherlands Antilles. Bull. Mar. Sci. 1995, 56, 609–619. [Google Scholar]
- Karlson, R.H.; Cornell, H.V. Species richness of coral assemblages: Detecting regional influences at local spatial scales. Ecol. 2002, 83, 452–463. [Google Scholar] [CrossRef]
- Yu, K.F. Introduction to Coral Reef Science; Science Press: Beijing, China, 2018; pp. 182–189. [Google Scholar]
- Yang, X.D.; Shen, Y.C.; Liu, L.; Jin, L. Effect of temperature, pH and salinity on growths of three species of coral nubbins. Mar. Environ. Sci. 2014, 33, 53–59. [Google Scholar]
- The WMO had Pointed Out that 2020 was One of the Three Warmest Three Years Since the Records. Available online: http://www.cma.gov.cn/2011xwzx/2011xmtjj/202104/t20210421_575664.html (accessed on 21 April 2021).
- Heron, S.; Eakin, C.M.; Douvere, F. Impacts of Climate Change on World Heritage Coral Reefs, A First Global Scientific Assessment. United Nations Educational Scientific and Cultural Organization (UNESCO). 2017. Available online: https://repository.library.noaa.gov/view/noaa/16386. (accessed on 29 December 2022).
- Chen, X.G.; Qian, G.M.; Chen, T.G.; Li, C.M.; Yu, K.F. Characteristics of climate warming in Guangdong and impact on climate belts change in Guangdong. J. Trop. Meteorol. 2006, 22, 547–552. [Google Scholar]
- Yu, K.F. The sea surface temperature changing trend of the last 40 years in the LeiQiong sea area. Trop. Geo. 2000, 20, 111–115. [Google Scholar]
- Lyu, Y.H.; Zhou, Z.H.; Zhang, Y.M.; Chen, Z.Q.; Deng, W.; Shi, R.G. The mass coral bleaching event of inshore corals form South China Sea witnessed in 2020: Insight into the causes, process and consequence. Coral Reefs. Report. 2022, 41, 1351–1364. [Google Scholar] [CrossRef]
- Yang, S.Y.; Chen, B.; Li, P.L. Study on the characteristics of the South China Sea water entering the Beibu Gulf through Qiongzhou Strait in summer by using temperature and salt data. J. Oceanol. Limn. 2006, 1, 1–7. [Google Scholar] [CrossRef]
- Risk, M.J. Paradise lost, how marine science failed the world’s coral reefs. Mar. Freshwater Res. 1999, 50, 831–837. [Google Scholar] [CrossRef]
- Yuan, X.C.; Guo, Y.J.; Cai, W.J.; Huang, H.; Zhou, W.H.; Liu, S. Coral responses to ocean warming and acidification: Implications for future distribution of coral reefs in the South China Sea. Mar. Pollut. Bull. 2019, 138, 241–248. [Google Scholar] [CrossRef]
- Wang, L.R.; Zhao, H.T.; Song, C.J. The impact of human activities on the coral reef ecosystem of Xuwen Dengloujiao. Ocean. Dev. Manag. 2006, 23, 81–85. [Google Scholar]
- Jordan, L.K.B.; Banks, K.W.; Fisher, L.E.; Walker, B.K.; Gilliam, D.S. Elevated sedimentation on coral reefs adjacent to a beach nourishment project. Mar. Pollut. Bull. 2010, 60, 261–271. [Google Scholar] [CrossRef]
- Schiel, D.R.; Wood, S.A.; Dunmore, R.A.; Taylor, D.I. Sediment on rocky intertidal reefs: Effects on early post-settlement stages of habitat-forming seaweeds. J. Exp. Mar. Biol. Ecol. 2006, 331, 158–172. [Google Scholar] [CrossRef]
- Wang, X.; Li, G.Z. Research status and Prospect of coral reefs in Weizhou Island, Beibu Gulf. J. Guangxi Acad. Sci. 2009, 25, 72–75, 80. [Google Scholar]
- Gilmour, J. Experimental investigation into the effects of suspended sediment on fertilisation, larval survival and settlement in a scleractinian coral. Mar. Biol. 1999, 135, 451–462. [Google Scholar] [CrossRef]
- Rogers, C.S. Responses of coral reefs and reef organisms to sedimentation. Mar. Ecol. Prog. Ser. 1990, 62, 185–202. [Google Scholar] [CrossRef]
- Xing, S.; Tan, Y.H.; Zhou, L.B.; Lian, X.P.; Huang, L.M. Effects of water turbidity on the symbiotic zooxanthella of hermatypic corals. Chin. Sci. Bull. 2012, 57, 348–354. [Google Scholar] [CrossRef]
- Stimson, J.; Sakai, K.; Sembali, H. Interspecific comparison of the symbiotic relationship in corals with high and low rates of bleaching-induced mortality. Coral Reefs. 2002, 21, 409–421. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs, review, and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Yang, Z.X.; Zhang, J.H.; Lv, X.L.; Zheng, Y.J.; Jiang, G.J.; Shi, X.J.; Tao, W.; Li, X.Q.; Lv, Y.H. The ecological succession of scleratinian coral communites and their environmental factors in Weizhou Island. Acta Ecol. Sin. 2021, 41, 7168–7179. [Google Scholar] [CrossRef]
- Song, L.X.; Lin, J.Y.; Huang, J.; Cheng, L.G. Dictionary of Chinese Salt History; Shanghai Dictionary Publishing House: Shanghai, China, 2010; p. 39. [Google Scholar]
- Yellowlees, D. Land Use Patterns and Nutrient Loading of the Great Barrier Reef Region. Proceedings of the Workshop Held at the James Cook University of North Queensland, 17–18 November 1990; Jamees Cook University Press: Townseille, Australian, 1991; pp. 162–179. [Google Scholar]
- Zhang, C.X.; Sun, S.L.; Xie, W.L.; Xie, S.Y.; Zhan, D.L.; Zhang, Y.B.; Zhang, J.B.; Chen, C.L. Seasonal changes of phytoplankton in Xuwen coral reef area. Oceanol. Limnol. Sin. 2009, 40, 159–165. [Google Scholar]
- Justić, D.; Rabalais, N.N.; Turner, R.E. Stoichiometric nutrient balance and origin of coastal eutrophication. Mar. Pollut. Bull. 1995, 30, 41–46. [Google Scholar] [CrossRef]
- Den, H.J.; Huisman, J.; Brocke, H.J.; Goehlich, H.; Latijnhouwers, K.R.; van, H.S.; Honcoop, S.A.S.; Bleyenberg, T.E.; Schouten, S.; Cerli, C.; et al. Nitrogen and phosphorus uptake rates of different species from a coral reef community after a nutrient pulse. Sci. Rep. 2016, 6, 28821. [Google Scholar] [CrossRef] [PubMed]
- Duprey, N.N.; Yasuhara, M.; Baker, D.M. Reefs of tomorrow: Eutrophication reduces coral biodiversity in an urbanized seascape. Glob. Chang. Biol. 2016, 22, 3550–3565. [Google Scholar] [CrossRef] [PubMed]
- Wiedenmann, J.; D’Angelo, C.; Smith, E.G.; Hunt, A.N.; Legiret, F.E.; Postle, A.D.; Achterberg, E.P. Nutrient enrichment can increase the susceptibility of reef corals to bleaching. Nat. Clim. Chang. 2013, 2, 160–164. [Google Scholar] [CrossRef]
- Thurber, R.L.V.; Burkepile, D.E.; Fuchs, C.; Shantz, A.A.; McMinds, R.; Zaneveld, J.R. Chronic nutrient enrichment increases prevalence and severity of coral disease and bleaching. Glob. Change. Biol. 2014, 20, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, Q.; Zhang, Y.; Ahmad, M.; Ling, J.; Tang, X.; Dong, J. Shifts in abundance and network complexity of coral bacteria in response to elevated ammonium stress. Sci. Total Environ. 2021, 768, 144631. [Google Scholar] [CrossRef]
- Erler, D.V.; Santos, I.R.; Eyre, B.D. Inorganic nitrogen transformations within permeable carbonate sands. Cont. Shelf Res. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Ning, Z.M.; Yu, K.F.; Wang, Y.H.; Huang, X.Y.; Hana, M.W.; Zhang, J. Carbon and nutrient dynamics of permeable carbonate and silicate sands adjacent to coral reefs around Weizhou Island in the northern South China Sea. Estuarine Coastal Shelf Sci. 2019, 225, 106229. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | Phylum/Family | Genus | Species | Dominant Species |
|---|---|---|---|---|
| Coral | Poritidae | Bernardpora | Bernardpora stutchburyi | a  b  c  Goniopora sp. (a–c) d  e  Bernardpora stutchburyi (d–e) |
| Goniopora | Goniopora columna | |||
| Goniopora djiboutiensis | ||||
| Goniopora gracilis | ||||
| Goniopora sp. | ||||
| Porites | Porites lobata | |||
| Porites lutea | ||||
| Porites sp. | ||||
| Agariciidae | Pavona | Pavona explanulata | ||
| Acroporidae | Acropora | Acropora humilis | ||
| Montipora | Montipora turgescens | |||
| Merulinidae | Astrea | Astrea curta | ||
| Coelastrea | Coelastrea palauensis | |||
| Cyphastrea | Cyphastrea serailia | |||
| Dipsastraea | Dipsastraea favus | |||
| Dipsastraea matthaii | ||||
| Dipsastraea rotumana | ||||
| Dipsastraea sp. | ||||
| Dipsastraea speciosa | ||||
| Echinopora | Echinopora gemmacea | |||
| Favites | Favites abdita | |||
| Favites flexuosa | ||||
| Favites halicora | ||||
| Favites pentagona | ||||
| Goniastrea | Goniastrea aspera | |||
| Goniastrea retiformis | ||||
| Merulina | Merulina ampliata | |||
| Platygyra | Platygyra daedalea | |||
| Platygyra sp. | ||||
| Dendrophylliidae | Turbinaria | Turbinaria peltata | ||
| Siderastreidae | Pseudosiderastrea | Pseudosiderastrea tayamai | ||
| Plesiastreidae | Plesiastrea | Plesiastrea versipora | ||
| Lobophylliidae | Lobophyllia | Lobophyllia corymbosa | ||
| Fish | Pomacentridae | Neopomacentrus | Neopomacentrus bankieri | f  Neopomacentrus bankieri (f) |
| Abudefduf | Abudefduf bengalensis | |||
| Pomadasyidae | Diagramma | Diagramma pictum | ||
| Lutjanidae | Lutjanus | Lutjanus ehrenbergii | ||
| Microdesmidae | Parioglossus | Parioglossus philippinus | ||
| Labridae | Halichoeres | Halichoeres nigrescens | ||
| Gerreidae | Gerres oyena | Gerres oyena | ||
| Algae | Chlorophyta | Boergesenia | Boergesenia forbesii | g  Caulerpa racemosa (g) |
| Bryopsis | Bryopsis pennata | |||
| Caulerpa | Caulerpa racemosa | |||
| Caulerpa sertularioides | ||||
| Ochrophyta | Dictyota | Dictyota sp. | ||
| Lobophora | Lobophora variegata | |||
| Padina | Padina sp. | |||
| Rhodophyta | Ceratodictyon | Ceratodictyon spongiosum | ||
| Hydropuntia | Hydropuntia edulis | |||
| Agarophyton | Agarophyton tenuistipitatum | |||
| Jania | Jania adhaerens | |||
| Laurencia | Laurencia decumbens | |||
| Peyssonnelia | Peyssonnelia rubra | |||
| Peyssonnelia sp. |
| Area | Family | Species | IVs | Percentage of IVs (%) |
|---|---|---|---|---|
| All | Poritidae | Goniopora gracilis | 1.438 | 11.6 |
| All | Poritidae | Bernardpora stutchburyi | 1.088 | 8.7 |
| All | Acroporidae | Montipora turgescens | 0.998 | 8.0 |
| All | Dendrophylliidae | Turbinaria peltata | 0.978 | 7.9 |
| All | Poritidae | Porites lutea | 0.962 | 7.7 |
| All | Merulinidae | Favites abdita | 0.773 | 6.2 |
| All | Plesiastreidae | Plesiastrea versipora | 0.707 | 5.7 |
| All | Poritidae | Goniopora djiboutiensis | 0.549 | 4.4 |
| 2 m | Poritidae | Goniopora gracilis | 1.401 | 11.9 |
| 2 m | Poritidae | Bernardpora stutchburyi | 1.204 | 10.2 |
| 2 m | Dendrophylliidae | Turbinaria peltata | 0.911 | 7.7 |
| 2 m | Poritidae | Porites lutea | 0.894 | 7.6 |
| 2 m | Merulinidae | Favites abdita | 0.844 | 7.1 |
| 2 m | Acroporidae | Montipora turgescens | 0.810 | 6.9 |
| 2 m | Plesiastreidae | Plesiastrea versipora | 0.794 | 6.7 |
| 4 m | Poritidae | Goniopora gracilis | 1.476 | 10.8 |
| 4 m | Acroporidae | Montipora turgescens | 1.340 | 9.8 |
| 4 m | Dendrophylliidae | Turbinaria peltata | 1.129 | 8.2 |
| 4 m | Poritidae | Porites lutea | 1.117 | 8.1 |
| 4 m | Poritidae | Bernardpora stutchburyi | 0.867 | 6.3 |
| 4 m | Poritidae | Goniopora djiboutiensis | 0.847 | 6.2 |
| 4 m | Poritidae | Goniopora sp. | 0.787 | 5.7 |
| 4 m | Poritidae | Porites lobata | 0.648 | 4.7 |
| Items | Abundance of Juvenile Corals (ind./m2.) | Mortality Rate (Recently Dead Corals, %) | Bleaching Rate (%) | Macroalgae Cover (%) | Density of Fish (ind./250 m2) | Sponge Cover (%) | Density of Invertebrates (ind./100 m2.) |
|---|---|---|---|---|---|---|---|
| Range | 0.20–8.80 | 0.0–7.1 | 0.0–8.2 | 0.0–7.7 | 0.00–49.00 | 0.0–4.4 | 77.00–245.00 |
| Average | 3.01 ± 2.39 | 2.8 ± 2.5 | 1.6 ± 2.3 | 0.7 ± 1.9 | 8.38 ± 13.56 | 1.3 ± 1.2 | 155.06 ± 51.05 |
| Items | Temperature/(°C) | Salinity | pH | DO/(mg/L) | COD/(mg/L) | Suspended Solids /(mg/L) | Transparency /(m) | Oils/(mg/L) |
|---|---|---|---|---|---|---|---|---|
| Range | 31.75–32.70 | 33.00–33.24 | 8.25–8.30 | 4.04–5.91 | 0.24–0.88 | 1.3–31.6 | 1.2–4.0 | 0.015–0.043 |
| Average | 32.29 ± 0.28 | 33.15 ± 0.07 | 8.27 ± 0.01 | 5.30 ± 0.42 | 0.49 ± 0.17 | 17.0 ± 10.5 | 2.1 ± 0.7 | 0.024 ± 0.008 |
| Items | NO3−-N/ (μmol/L) | NO2−-N / (μmol/L) | NH4+ -N / (μmol/L) | DIN/ (μmol/L) | PO43−-P / (μmol/L) | TP (μmol/L) | TN (μmol/L) | Chla/(µg/L) |
| Range | 0.89–2.89 | 0.08–1.46 | 1.13–2.31 | 2.19–5.31 | 0.00–0.32 | 0.22–0.70 | 4.11–17.46 | 2.17–19.36 |
| Average | 1.79 ± 0.53 | 0.35 ± 0.34 | 1.52 ± 0.35 | 3.66 ± 0.94 | 0.13 ± 0.11 | 0.35 ± 0.14 | 11.69 ± 4.11 | 7.90 ± 4.79 |
| Number | Features of Coral Community | Regression Equation with Environmental Factors |
|---|---|---|
| 1 | Living coral cover | Y = 1.913 − 53.431 × NH4+-N − 0.374 × Chla (P NH4+-N = 0.002, PChla = 0.015, p < 0.05) |
| 2 | Number of coral species | Y = 1.311 − 0.310 × Chla + 0.117 × FD (PChla = 0.005, PFD = 0.009, p < 0.01) |
| 3 | Coral diversity index (H’) | Y = 11.388 − 0.145 × Chla − 0.320 × Dept − 6.869 × Tem + 0.037 × FD (PChla = 0.005, PDept = 0.002, PTem = 0.022, PFD = 0.042, p < 0.05) |
| 4 | Abundance of juvenile corals | Y = −1.672 + 0.954 × ID + 0.258 × AH (PID = 0.004, PAH = 0.005, p < 0.05) |
| 5 | Coral bleaching | Y = 0.826 − 0.598 × GC (PGC = 0.000, p < 0.01) |
| Years | Number of Species | Living Coral Cover | Dominant Species | References |
|---|---|---|---|---|
| 2004 | 37 | 28.9% | Goniopora gracilis, Bernardpora stutchburyi, Favites abdita, Platygyra daedalea, Turbinaria sp. | Huang et al., 2007 [5], 2004, 18 sites |
| 2008 | 57 | 12.1% | Goniopora planulata, Turbinaria peltata, Cyphastrea serailia | Huang et al., 2011 [14], 2008, 25 sites |
| 2020 | 33 | 12.6% | Goniopora gracilis, Bernardpora stutchburyi, Montipora turgescens, Turbinaria peltate, Porites lutea | This study, 2020, 16 sites |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Tao, W.; Liu, Y.; Yu, W.; Peng, X.; Lu, C.; Jiang, G.; Chen, B.; Deng, W.; Lv, Y. The Ecological Status and Change in High-Latitude Coral Assemblages at the Xuwen Coral Reef, Northern South China Sea: Insight into the Status and Causes in 2020. Biology 2023, 12, 330. https://doi.org/10.3390/biology12020330
Yang Z, Tao W, Liu Y, Yu W, Peng X, Lu C, Jiang G, Chen B, Deng W, Lv Y. The Ecological Status and Change in High-Latitude Coral Assemblages at the Xuwen Coral Reef, Northern South China Sea: Insight into the Status and Causes in 2020. Biology. 2023; 12(2):330. https://doi.org/10.3390/biology12020330
Chicago/Turabian StyleYang, Zhenxiong, Wei Tao, Yue Liu, Wei Yu, Xiaojuan Peng, Chuqian Lu, Guangjia Jiang, Bin Chen, Wei Deng, and Yihua Lv. 2023. "The Ecological Status and Change in High-Latitude Coral Assemblages at the Xuwen Coral Reef, Northern South China Sea: Insight into the Status and Causes in 2020" Biology 12, no. 2: 330. https://doi.org/10.3390/biology12020330
APA StyleYang, Z., Tao, W., Liu, Y., Yu, W., Peng, X., Lu, C., Jiang, G., Chen, B., Deng, W., & Lv, Y. (2023). The Ecological Status and Change in High-Latitude Coral Assemblages at the Xuwen Coral Reef, Northern South China Sea: Insight into the Status and Causes in 2020. Biology, 12(2), 330. https://doi.org/10.3390/biology12020330