
Soil Moisture and Available Phosphorus as the Factors Driving Variation in Functional Characteristics across Different Restoration Communities in a Subtropical Mountain Ecosystem


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Community Composition Investigation and Sampling Analysis
2.3. Statistical Analysis
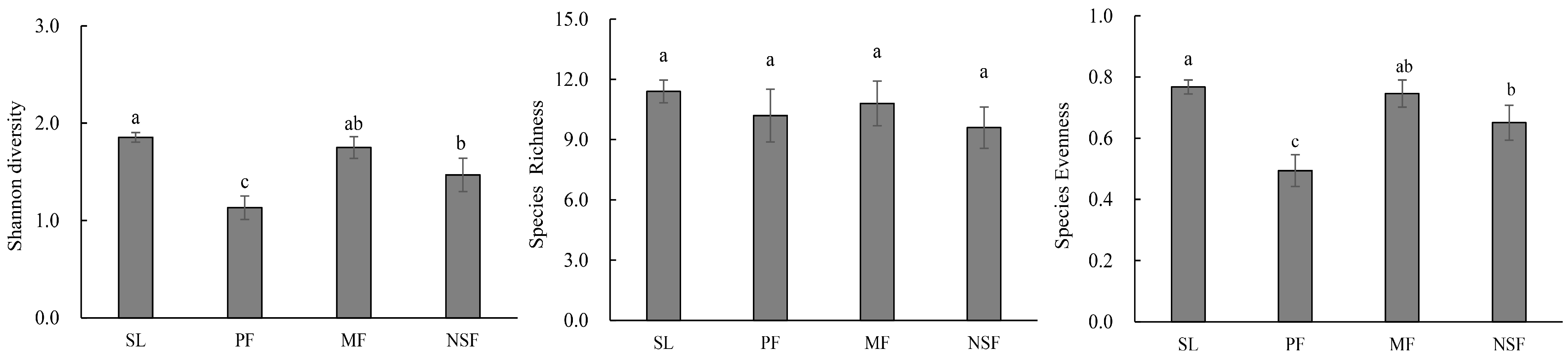
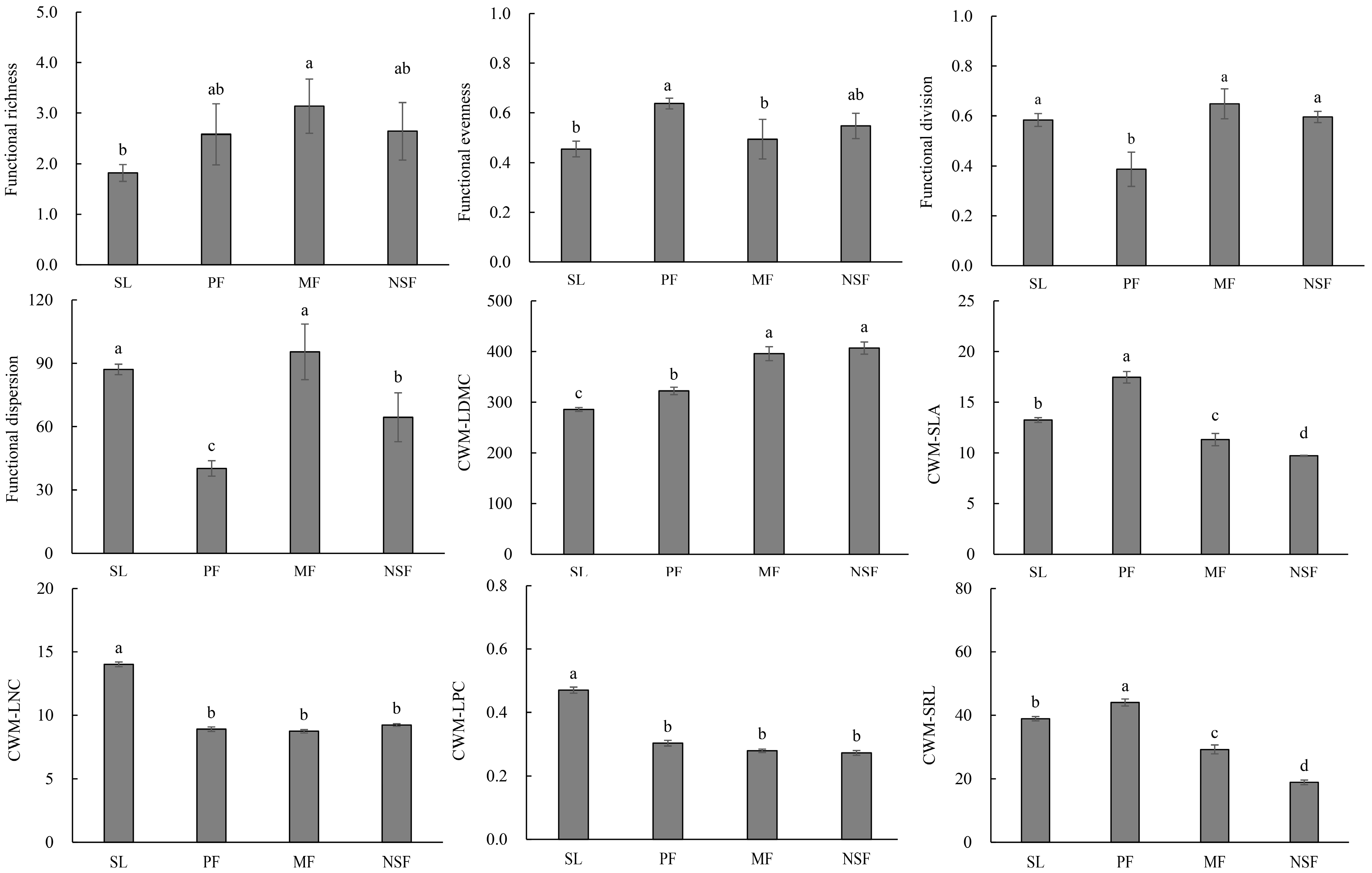
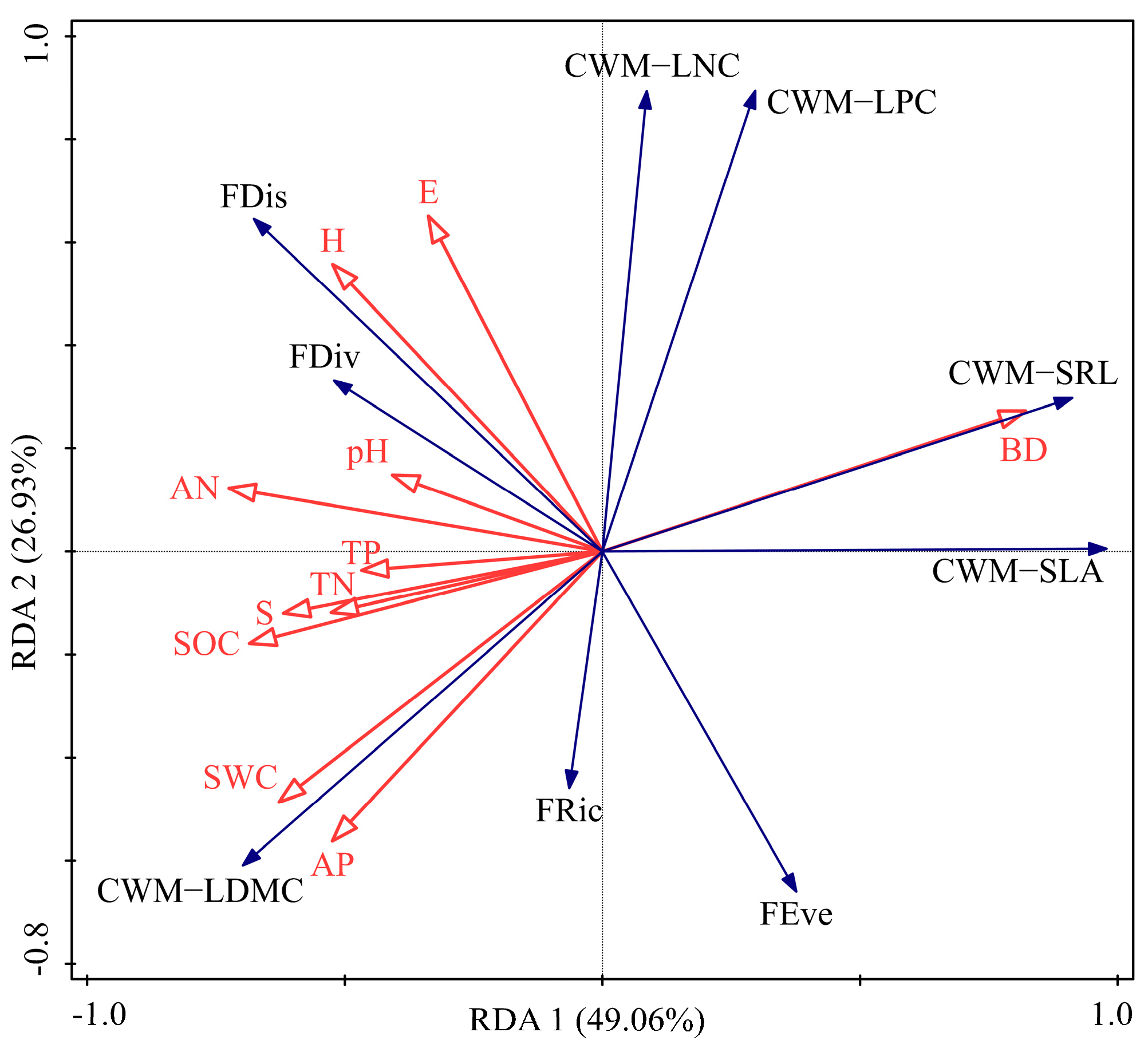
3. Results
3.1. Community Structural Properties and Soil Properties
3.2. Functional Characteristics of Plant Communities
3.3. Relationships among Functional Traits and Diversity Indices
3.4. Influence of Functional Diversity
4. Discussion
4.1. Functional Characteristics among Vegetation Restoration Types
4.2. Species Diversity and Functional Characteristics
4.3. Contributions of Soil Properties to Functional Characteristics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crouzeilles, R.; Curran, M.; Ferreira, M.S.; Lindenmayer, D.B.; Grelle, C.E.V.; Rey Benayas, J.M. A Global Meta-Analysis on the Ecological Drivers of Forest Restoration Success. Nat. Commun. 2016, 7, 11666. [Google Scholar] [CrossRef] [PubMed]
- Brudvig, L.A. The Restoration of Biodiversity: Where Has Research Been and Where Does It Need to Go? Am. J. Bot. 2011, 98, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayer, J.; Chokkalingam, U.; Poulsen, J. The Restoration of Forest Biodiversity and Ecological Values. For. Ecol. Manag. 2004, 201, 3–11. [Google Scholar] [CrossRef]
- Landesman, W.J.; Nelson, D.M.; Fitzpatrick, M.C. Soil Properties and Tree Species Drive SS-Diversity of Soil Bacterial Communities. Soil Biol. Biochem. 2014, 76, 201–209. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Drivers of Microbial Community Structure in Forest Soils. Appl. Microbiol. Biotechnol. 2018, 102, 4331–4338. [Google Scholar] [CrossRef]
- Bardgett, R.D. Plant Trait-Based Approaches for Interrogating Belowground Function. Biol. Environ. 2017, 117B, 1–13. [Google Scholar] [CrossRef]
- Faucon, M.P.; Houben, D.; Lambers, H. Plant Functional Traits: Soil and Ecosystem Services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef]
- Grigulis, K.; Lavorel, S.; Krainer, U.; Legay, N.; Baxendale, C.; Dumont, M.; Kastl, E.; Arnoldi, C.; Bardgett, R.D.; Poly, F.; et al. Relative Contributions of Plant Traits and Soil Microbial Properties to Mountain Grassland Ecosystem Services. J. Ecol. 2013, 101, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Culp, J.M.; Armanini, D.G.; Dunbar, M.J.; Orlofske, J.M.; Poff, N.L.; Pollard, A.I.; Yates, A.G.; Hose, G.C. Incorporating Traits in Aquatic Biomonitoring to Enhance Causal Diagnosis and Prediction. Integr. Environ. Assess. Manag. 2011, 7, 187–197. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional Richness, Functional Evenness and Functional Divergence: The Primary Components of Functional Diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing Functional Diversity in the Field—Methodology Matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant Functional Markers Capture Ecosystem Properties during Secondary Succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Strahan, R.T.; Sánchez Meador, A.J.; Huffman, D.W.; Laughlin, D.C. Shifts in Community-level Traits and Functional Diversity in a Mixed Conifer Forest: A Legacy of Land-use Change. J. Appl. Ecol. 2016, 53, 1755–1765. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, Z.; Castro, D.M.P.; Tan, X.; Jiang, X.; Meng, X.; Ge, Y.; Xie, Z. Effects of Different Types of Land-Use on Taxonomic and Functional Diversity of Benthic Macroinvertebrates in a Subtropical River Network. Environ. Sci. Pollut. Res. 2021, 28, 44339–44353. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Strahan, R.T.; Moore, M.M.; Fulé, P.Z.; Huffman, D.W.; Covington, W.W. The Hierarchy of Predictability in Ecological Restoration: Are Vegetation Structure and Functional Diversity More Predictable than Community Composition? J. Appl. Ecol. 2017, 54, 1058–1069. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Chiu, C.-H.; Jost, L. Unifying Species Diversity, Phylogenetic Diversity, Functional Diversity, and Related Similarity and Differentiation Measures through Hill Numbers. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 297–324. [Google Scholar] [CrossRef] [Green Version]
- Petchey, O.L.; Gaston, K.J. Functional Diversity (FD), Species Richness and Community Composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Poos, M.S.; Walker, S.C.; Jackson, D.A. Functional-diversity Indices Can Be Driven by Methodological Choices and Species Richness. Ecology 2009, 90, 341–347. [Google Scholar] [CrossRef]
- Schönbeck, L.; Lohbeck, M.; Bongers, F.; Martinez Ramos, M.; Sterck, F. How Do Light and Water Acquisition Strategies Affect Species Selection during Secondary Succession in Moist Tropical Forests? Forests 2015, 6, 2047–2065. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Vargas, G.; Sánchez-Velásquez, L.R.; López-Acosta, J.C.; Noa-Carrazana, J.C.; Perroni, Y. Relationship between Soil Properties and Leaf Functional Traits in Early Secondary Succession of Tropical Montane Cloud Forest. Ecol. Res. 2019, 34, 213–224. [Google Scholar] [CrossRef]
- Fukami, T.; Martijn Bezemer, T.; Mortimer, S.R.; van der Putten, W.H. Species Divergence and Trait Convergence in Experimental Plant Community Assembly. Ecol. Lett. 2005, 8, 1283–1290. [Google Scholar] [CrossRef]
- Galia Selaya, N.; Oomen, R.J.; Netten, J.J.C.; Werger, M.J.A.; Anten, N.P.R. Biomass Allocation and Leaf Life Span in Relation to Light Interception by Tropical Forest Plants during the First Years of Secondary Succession. J. Ecol. 2008, 96, 1211–1221. [Google Scholar] [CrossRef]
- Méndez-Toribio, M.; Meave, J.A.; Zermeño-Hernández, I.; Ibarra-Manríquez, G. Effects of Slope Aspect and Topographic Position on Environmental Variables, Disturbance Regime and Tree Community Attributes in a Seasonal Tropical Dry Forest. J. Veg. Sci. 2016, 27, 1094–1103. [Google Scholar] [CrossRef]
- Méndez-Toribio, M.; Ibarra-Manríquez, G.; Navarrete-Segueda, A.; Paz, H. Topographic Position, but Not Slope Aspect, Drives the Dominance of Functional Strategies of Tropical Dry Forest Trees. Environ. Res. Lett. 2017, 12, 85002. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A Handbook of Protocols for Standardised and Easy Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Casanoves, F.; Pla, L.; Di Rienzo, J.A.; Díaz, S. FDiversity: A Software Package for the Integrated Analysis of Functional Diversity. Methods Ecol. Evol. 2011, 2, 233–237. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Carter, M.R. Soil Sampling and Methods of Analysis; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- R Core Team R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Petchey, O.L. Integrating Methods That Investigate How Complementarity Influences Ecosystem Functioning. Oikos 2003, 101, 323–330. [Google Scholar] [CrossRef]
- Huang, C.; Xu, Y.; Zang, R. Variations in Functional Richness and Assembly Mechanisms of the Subtropical Evergreen Broadleaved Forest Communities along Geographical and Environmental Gradients. Forests 2022, 13, 1206. [Google Scholar] [CrossRef]
- Thakur, D.; Chawla, A. Functional Diversity along Elevational Gradients in the High Altitude Vegetation of the Western Himalaya. Biodivers. Conserv. 2019, 28, 1977–1996. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A Distance-Based Framework for Measuring Functional Diversity from Multiple Traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional Diversity Measures: An Overview of Their Redundancy and Their Ability to Discriminate Community Assembly Rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Fu, D.; Duan, C.; Hou, X.; Xia, T.; Gao, K. Patterns and Relationships of Plant Traits, Community Structural Attributes, and Eco-Hydrological Functions during a Subtropical Secondary Succession in Central Yunnan (Southwest China). Arch. Biol. Sci. 2009, 61, 741–749. [Google Scholar] [CrossRef]
- Funk, J.L.; Larson, J.E.; Ames, G.M.; Butterfield, B.J.; Cavender-Bares, J.; Firn, J.; Laughlin, D.C.; Sutton-Grier, A.E.; Williams, L.; Wright, J. Revisiting the H Oly G Rail: Using Plant Functional Traits to Understand Ecological Processes. Biol. Rev. 2017, 92, 1156–1173. [Google Scholar] [CrossRef]
- Brown, A.; Butler, D.W.; Radford-Smith, J.; Dwyer, J.M. Changes in Trait Covariance along an Orographic Moisture Gradient Reveal the Relative Importance of Light-and Moisture-driven Trade-offs in Subtropical Rainforest Communities. New Phytol. 2022, 236, 839–851. [Google Scholar] [CrossRef]
- Wagner, K.; Wanek, W.; Zotz, G. Functional Traits of a Rainforest Vascular Epiphyte Community: Trait Covariation and Indications for Host Specificity. Diversity 2021, 13, 97. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Laughlin, D.C. Constraints on Trait Combinations Explain Climatic Drivers of Biodiversity: The Importance of Trait Covariance in Community Assembly. Ecol. Lett. 2017, 20, 872–882. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, X.; Zhang, X.; Ju, W.; Duan, C.; Guo, X.; Wang, Y.; Fang, L. Soil Moisture Mediates Microbial Carbon and Phosphorus Metabolism during Vegetation Succession in a Semiarid Region. Soil Biol. Biochem. 2020, 147, 107814. [Google Scholar] [CrossRef]
- Hou, E.; Luo, Y.; Kuang, Y.; Chen, C.; Lu, X.; Jiang, L.; Luo, X.; Wen, D. Global Meta-Analysis Shows Pervasive Phosphorus Limitation of Aboveground Plant Production in Natural Terrestrial Ecosystems. Nat. Commun. 2020, 11, 637. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.; Li, Z.; Xu, Y.; Sun, J.; Zhao, X.; Huang, Z. Sapling Root Ecological Stoichiometric Characteristics of Dominant Species in Three Forests in Yunnan, China, and Their Relationship with Soil Physicochemical Factors. J. Veg. Sci. 2022, 33, e13166. [Google Scholar] [CrossRef]
- Klumpp, K.; Soussana, J. Using Functional Traits to Predict Grassland Ecosystem Change: A Mathematical Test of the Response-and-effect Trait Approach. Glob. Chang. Biol. 2009, 15, 2921–2934. [Google Scholar] [CrossRef]
- Li, W.; Epstein, H.E.; Wen, Z.; Zhao, J.; Jin, J.; Jing, G.; Cheng, J.; Du, G. Community-Weighted Mean Traits but Not Functional Diversity Determine the Changes in Soil Properties during Wetland Drying on the Tibetan Plateau. Solid Earth 2017, 8, 137–1477. [Google Scholar] [CrossRef] [Green Version]
- Seyfried, G.S.; Dalling, J.W.; Yang, W.H. Mycorrhizal Type Effects on Leaf Litter Decomposition Depend on Litter Quality and Environmental Context. Biogeochemistry 2021, 155, 21–38. [Google Scholar] [CrossRef]
- Carteron, A.; Cichonski, F.; Laliberté, E. Ectomycorrhizal Stands Accelerate Decomposition to a Greater Extent than Arbuscular Mycorrhizal Stands in a Northern Deciduous Forest. Ecosystems 2022, 25, 1234–1248. [Google Scholar] [CrossRef]
- Adeoyo, O.R. A Review on Mycorrhizae and Related Endophytic Fungi as Potential Sources of Enzymes for the Bio-Economy. J. Adv. Microbiol 2021, 21, 35–49. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type | Age (Year) | DBH (cm) | Tree Density (Trees/ha) | Cover | Common Dominant Species |
|---|---|---|---|---|---|
| SL | 50–60 | — | — | 0.73 ± 0.16 | Ternstroemia gymnanthera, Eupatorium adenophorum |
| PF | 40–50 | 10.34 ± 2.68 | 2436 ± 1244 | 0.64 ± 0.15 | Pinus yunnanensis |
| MF | 40–50 | 9.77 ± 2.42 | 3946 ± 2803 | 0.83 ± 0.21 | P. yunnanensis, Keteleeria evelyniana, Cyclobalanopsis glaucoides |
| NSF | 10–20 | 7.43 ± 2.65 | 6612 ± 3733 | 0.85 ± 0.22 | K. evelyniana, C. glaucoides, Lithocarpus dealbatus |
| Soil Index | SL | PF | MF | NSF |
|---|---|---|---|---|
| pH | 4.25 ± 0.23 a | 4.16 ± 0.07 a | 4.32 ± 0.04 a | 4.28 ± 0.17 a |
| BD (g/cm3) | 1.25 ± 0.05 a | 1.34 ± 0.02 a | 1.14 ± 0.06 b | 0.98 ± 0.07 c |
| SWC (%) | 27.33 ± 6.23 ab | 25.36 ± 2.76 b | 34.88 ± 3.08 a | 38.45 ± 4.62 a |
| TN (%) | 0.08 ± 0.01 ab | 0.06 ± 0.02 ab | 0.05 ± 0.02 b | 0.10 ± 0.03 a |
| AN (mg/kg) | 81.01 ± 6.56 a | 54.74 ± 24.68 a | 59.71 ± 18.25 a | 107.81 ± 32.59 a |
| TP (%) | 0.02 ± 0.00 b | 0.01 ± 0.00 c | 0.02 ± 0.00 b | 0.03 ± 0.00 a |
| AP (mg/kg) | 0.64 ± 0.06 c | 1.35 ±0.53 ab | 1.04 ± 0.09 bc | 2.15 ± 1.15 a |
| SOM (%) | 2.03 ± 0.08 ab | 1.57 ± 0.58 b | 1.71 ± 0.10 ab | 2.75 ± 0.58 a |
| LDMC | SLA | LNC | LPC | SRL | |
|---|---|---|---|---|---|
| LDMC | −0.58 ** | −0.71 ** | −0.86 ** | −0.64 ** | |
| SLA | −0.26 * | 0.31 | 0.54 ** | 0.94 ** | |
| LNC | −0.44 ** | 0.50 ** | 0.74 ** | 0.22 | |
| LPC | −0.40 ** | 0.55 ** | 0.67 ** | 0.58 ** | |
| SRL | −0.34 ** | 0.37 ** | 0.47 ** | 0.42 ** |
| Functional Diversity | Species Diversity | ||
|---|---|---|---|
| Shannon | Richness | Evenness | |
| FRic | 0.09 | 0.44 | −0.08 |
| FEve | −0.16 | 0.01 | −0.15 |
| FDiv | 0.53 ** | −0.17 | 0.68 ** |
| FDis | 0.80 ** | 0.19 | 0.84 ** |
| CWM-LDMC | −0.17 | −0.24 | −0.07 |
| CWM-SLA | −0.37 | 0.13 | −0.49 * |
| CWM-LNC | 0.55 ** | 0.26 | 0.49 * |
| CWM-LPC | 0.47 * | 0.29 | 0.38 |
| CWM-SRL | −0.10 | 0.18 | −0.20 |
| RDA1 | RDA2 | R2 | P | |
|---|---|---|---|---|
| H | 0.21 | −0.98 | 0.67 | 0.01 ** |
| S | 0.89 | −0.45 | 0.61 | 0.01 ** |
| E | −0.10 | −1.00 | 0.54 | 0.04 * |
| pH | 0.63 | −0.78 | 0.19 | 0.38 |
| SWC | 0.99 | 0.16 | 0.59 | 0.02 * |
| BD | −0.99 | 0.11 | 0.45 | 0.05 |
| SOM | 1.00 | −0.09 | 0.44 | 0.05 |
| TN | 0.99 | 0.17 | 0.04 | 0.84 |
| TP | 0.97 | 0.22 | 0.07 | 0.72 |
| AN | 0.69 | −0.72 | 0.15 | 0.49 |
| AP | 0.88 | 0.47 | 0.56 | 0.02 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Shen, C.; Ma, X.; Hu, L.; He, Y.; Shang, H.; Fu, D. Soil Moisture and Available Phosphorus as the Factors Driving Variation in Functional Characteristics across Different Restoration Communities in a Subtropical Mountain Ecosystem. Biology 2023, 12, 427. https://doi.org/10.3390/biology12030427
Wu X, Shen C, Ma X, Hu L, He Y, Shang H, Fu D. Soil Moisture and Available Phosphorus as the Factors Driving Variation in Functional Characteristics across Different Restoration Communities in a Subtropical Mountain Ecosystem. Biology. 2023; 12(3):427. https://doi.org/10.3390/biology12030427
Chicago/Turabian StyleWu, Xiaoni, Chunjie Shen, Xudong Ma, Lianyu Hu, Yongjian He, Huaye Shang, and Denggao Fu. 2023. "Soil Moisture and Available Phosphorus as the Factors Driving Variation in Functional Characteristics across Different Restoration Communities in a Subtropical Mountain Ecosystem" Biology 12, no. 3: 427. https://doi.org/10.3390/biology12030427
APA StyleWu, X., Shen, C., Ma, X., Hu, L., He, Y., Shang, H., & Fu, D. (2023). Soil Moisture and Available Phosphorus as the Factors Driving Variation in Functional Characteristics across Different Restoration Communities in a Subtropical Mountain Ecosystem. Biology, 12(3), 427. https://doi.org/10.3390/biology12030427