
Phylogenomics and Biogeography of the Mammilloid Clade Revealed an Intricate Evolutionary History Arose in the Mexican Plateau

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
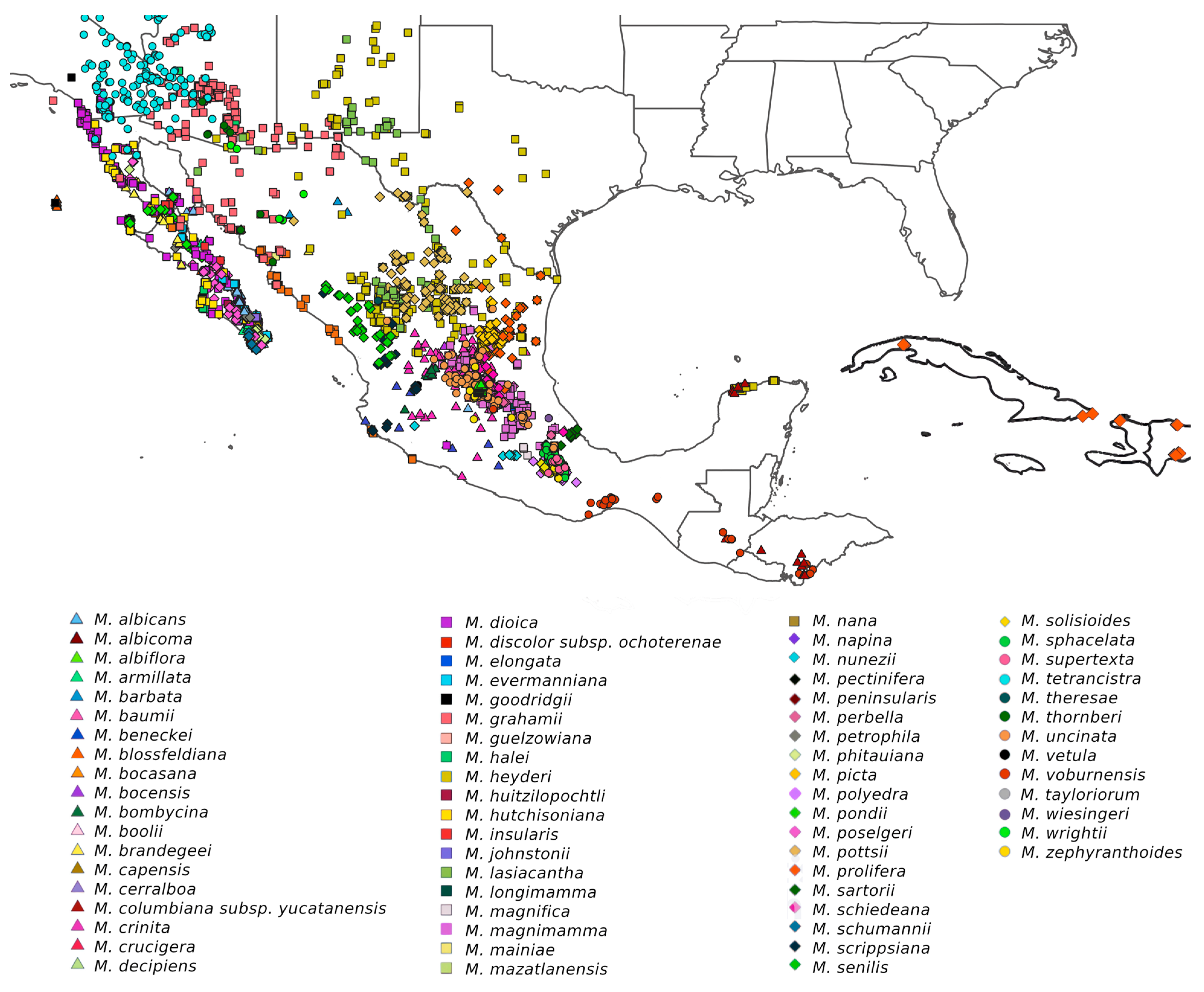
2.1. Taxon Sampling
2.2. DNA Extraction, cpDNA Enrichment, and High-Throughput Sequencing
2.3. De Novo Assembly of Chloroplast Genomes
2.4. Phylogenetic Relationships and Divergence Times
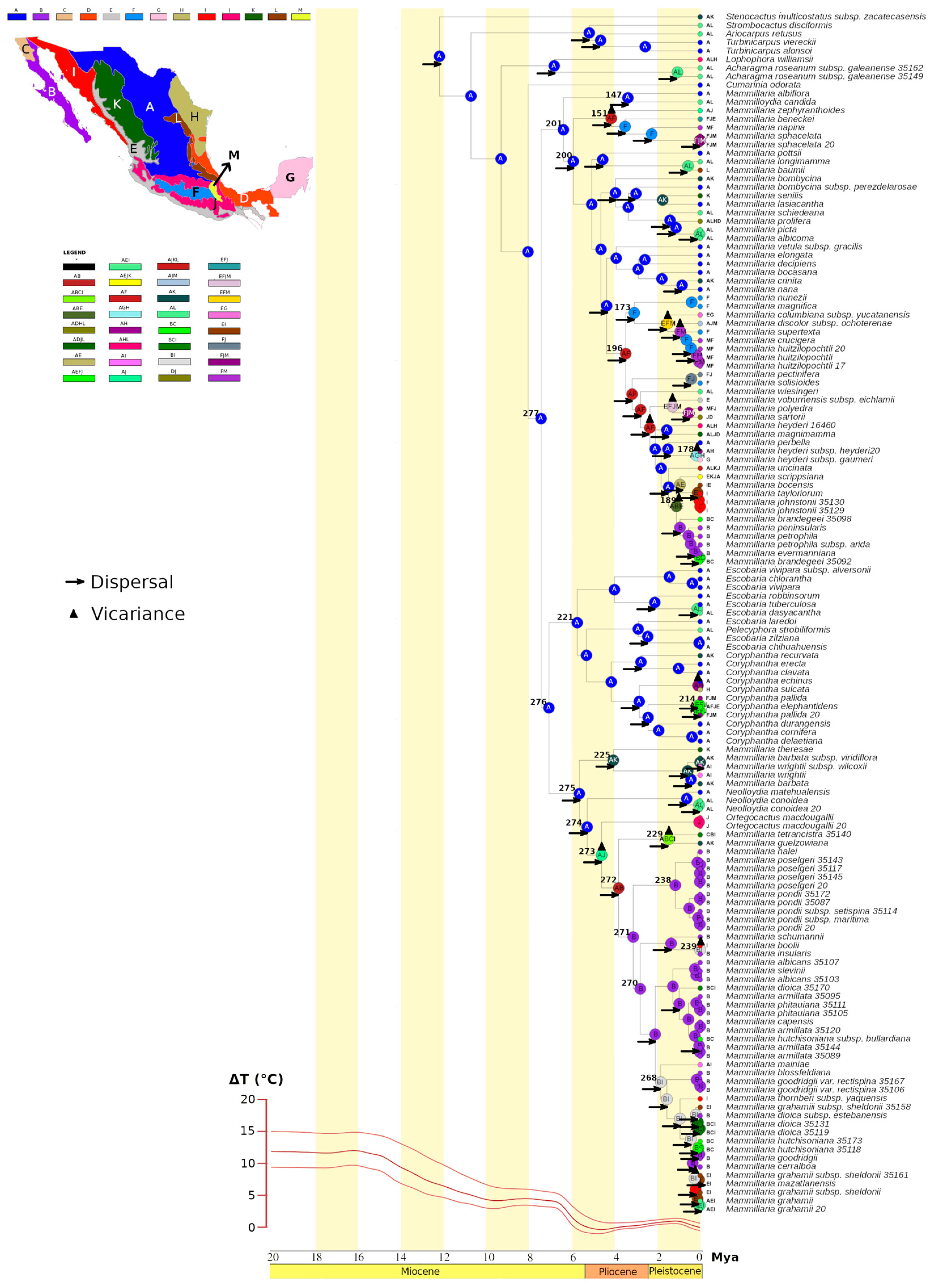
2.5. Biogeographical Analysis
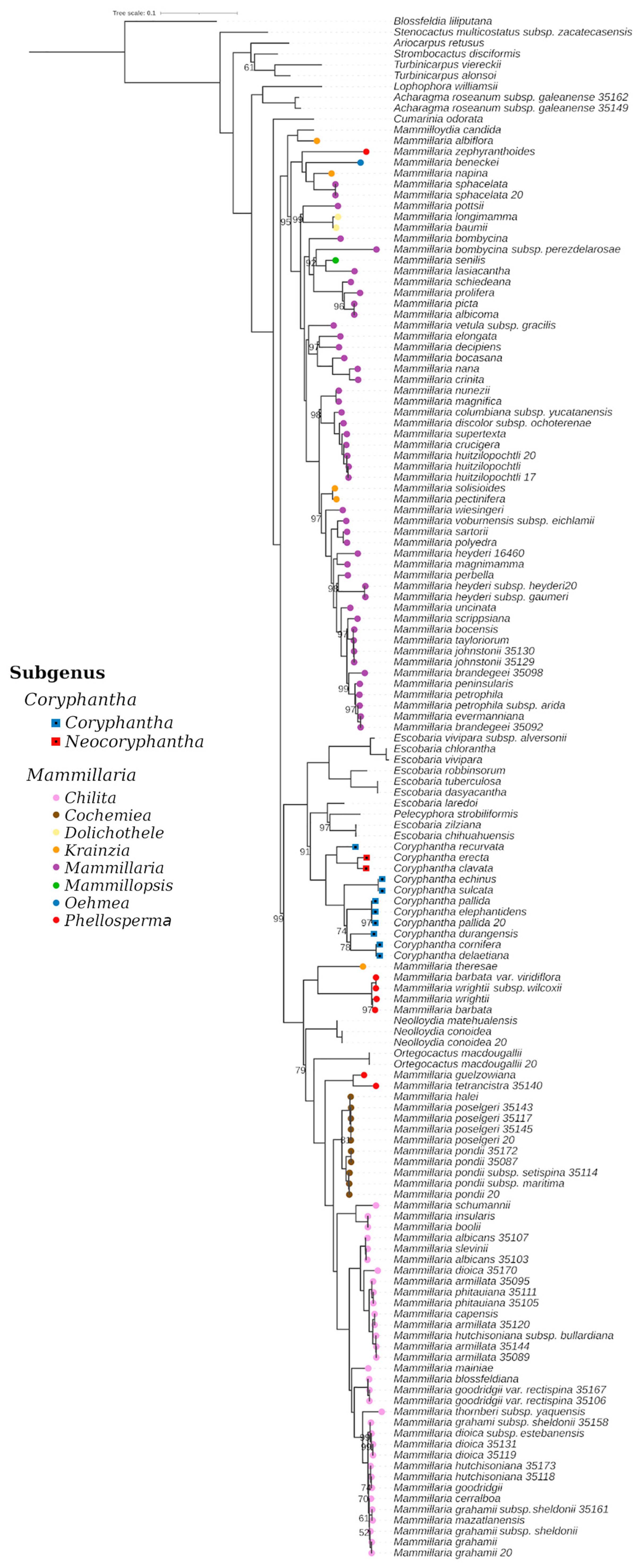
3. Results
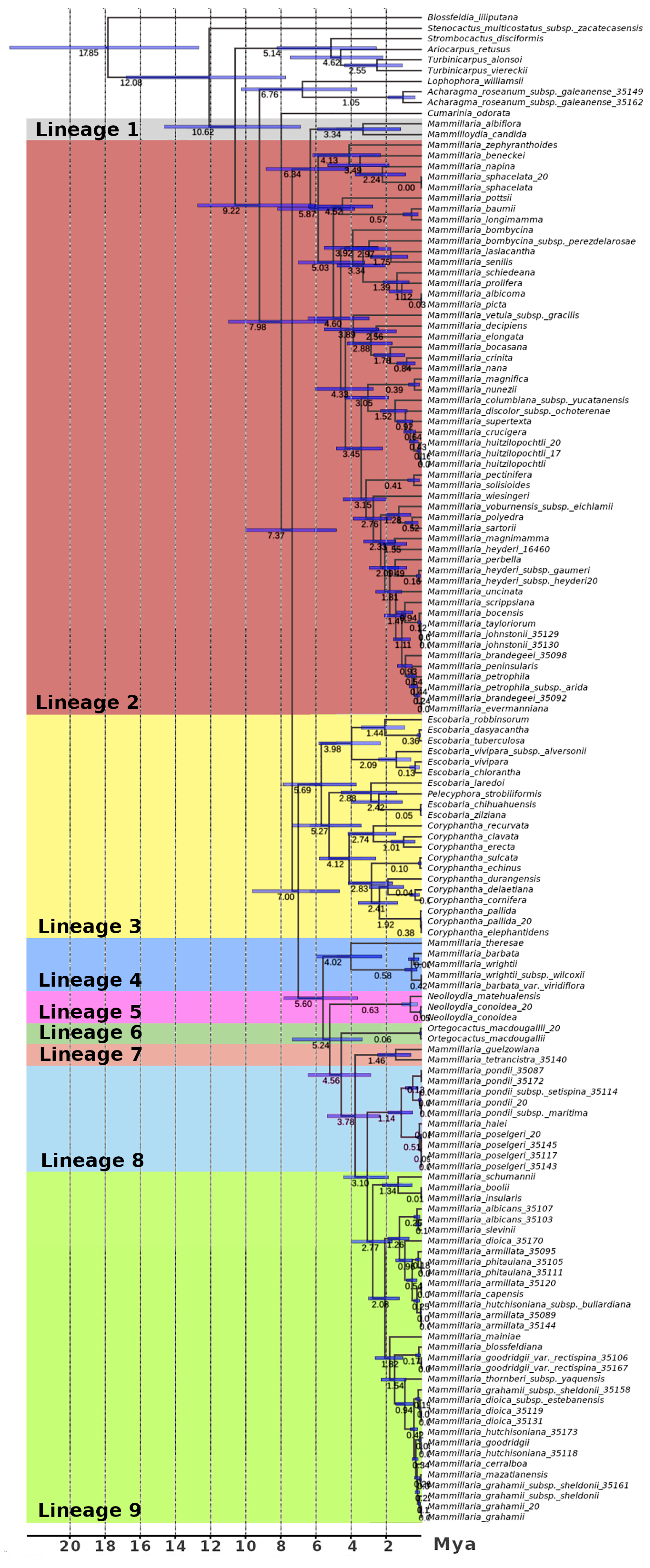
3.1. Evolutionary History of Cacti: Recent Divergence and Intricate Biogeography
3.2. Evolutionary History of the Nine Lineages
3.2.1. Evolutionary Lineage 1
3.2.2. Evolutionary Lineage 2
3.2.3. Evolutionary Lineage 3
3.2.4. Evolutionary Lineage 4
3.2.5. Evolutionary Lineage 5
3.2.6. Evolutionary Lineage 6
3.2.7. Evolutionary Lineage 7
3.2.8. Evolutionary Lineage 8
3.2.9. Evolutionary Lineage 9
4. Discussion
4.1. Origin and Diversification of the Mammilloid Clade
4.2. Taxonomic Contributions of the Phylogenetic Results
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ronquist, F.; Sanmartín, I. Phylogenetic methods in biogeography. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 441–464. [Google Scholar] [CrossRef] [Green Version]
- Xia, M.; Liu, Y.; Liu, J.; Chen, D.; Shi, Y.; Chen, Z.; Chen, D.; Jin, R.; Chen, H.; Peter, C.H.; et al. Out of the Himalaya-Hengduan Mountains: Phylogenomics, biogeography and diversification of Polygonatum Mill. (Asparagaceae) in the Northern Hemisphere. Mol. Phylogenet. Evol. 2022, 169, 107431. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.P.; Xiang, C.L.; Sytsma, K.J.; Drew, B.T. A timeframe for mint evolution: Towards a better understanding of trait evolution and historical biogeography in Lamiaceae. Bot. J. Linn. Soc. 2022, 200, 15–38. [Google Scholar] [CrossRef]
- Liu, S.Y.; Zhu, H.; Yang, J. A phylogenetic perspective on biogeographical divergence of the flora in Yunnan, Southwestern China. Sci. Rep. 2017, 7, 43032. [Google Scholar] [CrossRef]
- Qian, H.; Deng, T.; Ricklefs, R.E. Evolutionary assembly of the Arctic flora. Glob. Ecol. Biogeogr. 2022, 31, 396–404. [Google Scholar] [CrossRef]
- Barros-Souza, Y.; Borges, L.M. Spatial-and lineage-dependent processes underpin floristic assembly in the megadiverse Eastern South American mountains. J. Biogeogr. 2023, 50, 302–315. [Google Scholar] [CrossRef]
- Chen, L.Y.; Zhao, S.Y.; Mao, K.S.; Les, D.H.; Wang, Q.F.; Moody, M.L. Historical biogeography of Haloragaceae: An out-of-Australia hypothesis with multiple intercontinental dispersals. Mol. Phylogenet. Evol. 2014, 78, 87–95. [Google Scholar] [CrossRef]
- Nge, F.J.; Kellermann, J.; Biffin, E.; Waycott, M.; Thiele, K.R. Historical biogeography of Pomaderris (Rhamnaceae): Continental vicariance in Australia and repeated independent dispersals to New Zealand. Mol. Phylogenet. Evol. 2021, 158, 107085. [Google Scholar] [CrossRef]
- Liu, X.Q.; Xia, X.M.; Chen, L.; Wang, X.Q. Phylogeny and evolution of Cupressaceae: Updates on intergeneric relationships and new insights on ancient intergeneric hybridization. Mol. Phylogenet. Evol. 2022, 177, 107606. [Google Scholar] [CrossRef]
- Heinrichs, J.; Hentschel, J.; Feldberg, K.; Bombosch, A.; Schneider, H. Phylogenetic biogeography and taxonomy of disjunctly distributed bryophytes. J. Syst. Evol. 2009, 47, 497–508. [Google Scholar] [CrossRef]
- Oyston, J.W.; Wilkinson, M.; Ruta, M.; Wills, M.A. Molecular phylogenies map to biogeography better than morphological ones. Commun. Biol. 2022, 5, 521. [Google Scholar] [CrossRef] [PubMed]
- Svenning, J.-C.; Eiserhardt, W.L.; Normand, S.; Ordonez, A.; Sandel, B. The influence of paleoclimate on present-day patterns in biodiversity and ecosystems. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 551–572. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.Z.; Lehtonen, S.; Martins, K.; Gichira, A.W.; Wu, S.; Li, W.; Hu, G.W.; Liu, Y.; Zou, C.Y.; Wang, Q.F.; et al. Phylogenomics of the aquatic plant genus Ottelia (Hydrocharitaceae): Implications for historical biogeography. Mol. Phylogenet. Evol. 2020, 152, 106939. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.J.; Li, S.J.; Wang, W.M. Evolutionary trends in leaf morphology and biogeography of Altingiaceae based on fossil evidence. Palaeoworld 2018, 27, 415–422. [Google Scholar] [CrossRef]
- Ding, W.N.; Ree, R.H.; Spicer, R.A.; Xing, Y.W. Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science 2020, 369, 578–581. [Google Scholar] [CrossRef]
- Antonelli, A.; Nylander, J.A.A.; Persson, C.; Sanmartín, I. Tracing the impact of the Andean uplift on Neotropical plant evolution. Proc. Natl. Acad. Sci. USA 2009, 106, 9749–9754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitt, G.M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Hunt, D.R. The New Cactus Lexicon; DH Books: Milborne Port, UK, 2006. [Google Scholar]
- Nyffeler, R.; Eggli, U. Disintegrating Portulacaceae: A new familial classification of the suborder Portulacineae (Caryophyllales) based on molecular and morphological data. Taxon 2010, 59, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.F.; Yang, Y.; Feng, T.; Timoneda, A.; Mikenas, J.; Hutchison, V.; Edwards, C.; Wang, N.; Ahluwalia, S.; Olivieri, J.; et al. From cacti to carnivores: Improved phylotranscriptomic sampling and hierarchical homology inference provide further insight into the evolution of Caryophyllales. Am. J. Bot. 2018, 105, 446–462. [Google Scholar]
- Magallon, S.; Gomez-Acevedo, S.; Sanchez-Reyes, L.L.; Hernandez-Hernandez, T. A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity. New Phytol. 2015, 207, 437–453. [Google Scholar] [CrossRef]
- Hernandez-Hernandez, T.; Brown, J.W.; Schlumpberger, B.O.; Eguiarte, L.E.; Magallon, S. Beyond aridification: Multiple explanations for the elevated diversification of cacti in the New World Succulent Biome. New Phytol. 2014, 202, 1382–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakaki, M.; Christin, P.A.; Nyffeler, R.; Lendel, A.; Eggli, U.; Ogburn, M.; Spriggs, E.; Moore, M.J.; Edwards, E.J. Contemporaneous and recent radiations of the world’s major succulent plant lineages. Proc. Natl. Acad. Sci. USA 2011, 108, 8379–8384. [Google Scholar] [CrossRef] [Green Version]
- Pearson, P.N.; Palmer, M.R. Atmospheric carbon dioxide concentrations over the past 60 million years. Nature 2000, 406, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Tripati, A.K.; Roberts, C.D.; Eagle, R.A. Coupling of CO2 and Ice Sheet Stability Over Major Climate Transitions of the Last 20 Million Years. Science 2009, 326, 1394–1397. [Google Scholar] [CrossRef] [Green Version]
- Herbert, T.D.; Lawrence, K.T.; Tzanova, A.; Peterson, L.C.; Caballero-Gill, R.; Kelly, C.S. Late Miocene global cooling and the rise of modern ecosystems. Nat. Geosci. 2016, 9, 843. [Google Scholar] [CrossRef]
- Anderson, E.F. The Cactus Family; Timber: Portland, OR, USA, 2001. [Google Scholar]
- The IUCN Red List of Threatened Species. Version 2022-1. Available online: https://www.iucnredlist.org/search?taxonomies=111456&searchType=species (accessed on 1 November 2022).
- Haworth, A.H. Synopsis Plantarum Succulentarum; Richard Taylor: London, UK, 1812. [Google Scholar]
- Schumann, K. Gesamtbeschreibung der Kakteen (Monographia Cactearum); Verlag von J. Neumann: Neudamm, Germany, 1898; pp. 472–601. [Google Scholar]
- Hunt, D.R. Schumann and Buxbaum reconciled. Cact. Succ. J. Gr. Brit. 1971, 33, 53–72. [Google Scholar]
- Lüthy, J.M. Taxonomische Untersuchung der Gattung Mammillaria Haw. Ph.D. Thesis, Universität Bern, Bern, Switzerland, 1995. [Google Scholar]
- Hernández, H.M.; Gómez-Hinostrosa, C. Mapping the Cacti of Mexico Part II: Mammillaria; DH Books: Milborne Port, UK, 2015. [Google Scholar]
- Butterworth, C.A.; Cota-Sanchez, J.H.; Wallace, R.S. Molecular systematics of tribe Cacteae (Cactaceae: Cactoideae): A phylogeny based on rpl16 intron sequence variation. Syst. Bot. 2002, 27, 257–270. [Google Scholar]
- Butterworth, C.A.; Wallace, R.S. Phylogenetic studies of Mammillaria (Cactaceae)—Insights from chloroplast sequence variation and hypothesis testing using the parametric bootstrap. Am. J. Bot. 2004, 91, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Crozier, B.S. Systematics of Cactaceae Juss: Phylogeny, cpDNA Evolution, and Classification, with Emphasis on the Genus Mammillaria Haw. Ph.D. Thesis, University of Texas, Austin, TX, USA, 2005. [Google Scholar]
- Hunt, D.R. Revised classified list of the genus Mammillaria. Cact. Succ. J. Gr. Brit. 1981, 43, 41–48. [Google Scholar]
- Breslin, P.B.; Wojciechowski, M.F.; Majure, L.C. Molecular phylogeny of the Mammilloid clade (Cactaceae) resolves the monophyly of Mammillaria. Taxon 2021, 70, 308–323. [Google Scholar] [CrossRef]
- Solórzano, S.; Chincoya, D.A.; Sanchez-Flores, A.; Estrada, K.; Díaz-Velásquez, C.E.; González-Rodríguez, A.; Vaca-Paniagua, F.; Dávila, P.; Arias, S. De novo assembly discovered novel structures in genome of plastids and revealed divergent inverted repeats in Mammillaria (Cactaceae, Caryophyllales). Plants 2019, 8, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P.; Bjornson, R.; Weston, S. spThin: Functions for Spatial Thinning of Species Occurrence Records for Use in Ecological Models. 2019. Available online: https://cran.r-project.org/web/packages/spThin/index.html (accessed on 15 March 2023).
- Babraham Bioinformatics. Trim Galore. Available online: http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 21 April 2022).
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. Model Finder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 7, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Cruz, L.U.; Arias-Montes, S.; Dávila, P. Catálogo de Cactáceas Mexicanas; Universidad Nacional Autónoma de México, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, Mexico, 2007. [Google Scholar]
- Dicht, R.; Lüthy, A. Coryphantha: Cacti of Mexico and Southern USA; Springer: Berlin, Germany, 2005. [Google Scholar]
- Rzedowski, J. Vegetación de México; Limusa: Ciudad de México, Mexico, 1978. [Google Scholar]
- Matzke, N. Probabilistic historical biogeography: New models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Front. Biogeogr. 2015, 5, 242–248. [Google Scholar]
- Yu, Y.; Blair, C.; He, X. RASP 4: Ancestral state reconstruction tool for multiple genes and characters. Mol. Biol. Evol. 2020, 37, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.T.; Sosdian, S.; White, H.E.; Rosenthal, Y. North Atlantic climate evolution through the Plio-Pleistocene climate transitions. Earth Planet. Sci. Lett. 2010, 300, 329–342. [Google Scholar] [CrossRef]
- Vuilleumier, B.S. Pleistocene changes in the fauna and flora of South America. Science 1971, 173, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Hoag, C.; Svenning, J.C. African environmental change from the Pleistocene to the Anthropocene. Annu. Rev. Environ. Resour. 2017, 42, 27–54. [Google Scholar] [CrossRef] [Green Version]
- Janssens, S.B.; Knox, E.B.; Huysmans, S.; Smets, E.F.; Merckx, V.S. Rapid radiation of Impatiens (Balsaminaceae) during pliocene and pleistocene: Result of a global climate change. Mol. Phylogenet. Evol. 2009, 52, 806–824. [Google Scholar] [CrossRef]
- DeChaine, E.G.; Wendling, B.M.; Forester, B.R. Integrating environmental, molecular, and morphological data to unravel an ice-age radiation of arctic-alpine Campanula in western North America. Ecol. Evol. 2014, 4, 3940–3959. [Google Scholar] [CrossRef] [Green Version]
- Dorsey, B.L.; Gregory, T.J.; Sass, C.; Specht, C.D. Pleistocene diversification in an ancient lineage: A role for glacial cycles in the evolutionary history of Dioon Lindl. (Zamiaceae). Am. J. Bot. 2018, 105, 1512–1530. [Google Scholar] [CrossRef] [Green Version]
- Nevado, B.; Contreras-Ortiz, N.; Hughes, C.; Filatov, D.A. Pleistocene glacial cycles drive isolation, gene flow and speciation in the high-elevation Andes. New Phytol. 2018, 219, 779–793. [Google Scholar] [CrossRef] [Green Version]
- Cornejo-Romero, A.; Vargas-Mendoza, C.F.; Aguilar-Martínez, G.F.; Medina-Sánchez, J.; Rendón-Aguilar, B.; Valverde, P.L.; Zavala-Hurtado, J.A.; Serrato, A.; Rivas-Arancibia, S.; Pérez-Hernández, M.A.; et al. Alternative glacial-interglacial refugia demographic hypotheses tested on Cephalocereus columna-trajani (Cactaceae) in the intertropical Mexican drylands. PLoS ONE 2017, 12, e0175905. [Google Scholar] [CrossRef] [Green Version]
- Ossa, C.G.; Montenegro, P.; Larridon, I.; Pérez, F. Response of xerophytic plants to glacial cycles in southern South America. Ann. Bot. 2019, 124, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Bonatelli, I.A.; Perez, M.F.; Peterson, A.T.; Taylor, N.P.; Zappi, D.C.; Machado, M.C.; Koch, I.; Pires, A.H.C.; Moraes, E.M. Interglacial microrefugia and diversification of a cactus species complex: Phylogeography and palaeodistributional reconstructions for Pilosocereus aurisetus and allies. Mol. Ecol. 2014, 23, 3044–3063. [Google Scholar] [CrossRef] [PubMed]
- Ornelas, J.F.; Rodríguez-Gómez, F. Influence of Pleistocene glacial/interglacial cycles on the genetic structure of the Mistletoe Cactus Rhipsalis baccifera (Cactaceae) in Mesoamerica. J. Hered. 2015, 106, 196–210. [Google Scholar] [CrossRef] [Green Version]
- Morafka, D.J. A historical biogeography of the Chihuahuan herpetofauna. In A Biogeographical Analysis of the Chihuahuan Desert through Its Herpetofauna; Dr. W. Junk: The Hague, The Netherlands, 1977; pp. 157–215. [Google Scholar]
- UNESCO. ATLAS of Arid Zones in Latin America and the Caribbean; PHI UNESCO: Paris, France, 2010. [Google Scholar]
- Guerrero, P.C.; Durán, A.P.; Walter, H.E. Latitudinal and altitudinal patterns of the endemic cacti from the Atacama Desert to Mediterranean Chile. J. Environ. 2011, 75, 991–997. [Google Scholar] [CrossRef]
- Medellín-Leal, F. The Chihuahuan Desert. In Reference Handbook on the Deserts of North America; Bender, G.L., Ed.; Greenwood Press: Westfield, CT, USA, 1982; pp. 321–381. [Google Scholar]
- Jürgens, N.; Oncken, I.; Oldeland, J.; Gunter, F.; Rudolph, B. Welwitschia: Phylogeography of a living fossil, diversified within a desert refuge. Sci. Rep. 2021, 11, 114. [Google Scholar] [CrossRef]
- Puente, M.E.; Li, C.Y.; Bashan, Y. Rock-degrading endophytic bacteria in cacti. Environ. Exp. Bot. 2009, 66, 389–401. [Google Scholar] [CrossRef]
- Bregman, R. Forms of seed dispersal in Cactaceae. Acta Bot. Neerl. 1988, 37, 395–402. [Google Scholar] [CrossRef]
- Gómez-Tuena, A.; Orozco-Esquivel, M.T.; Ferrari, L. Igneous petrogenesis of the Trans-Mexican volcanic belt. Geo. Soc. Amer. Spec. 2007, 422, 129–181. [Google Scholar]
- Ledesma-Vázquez, J.; Johnson, M.E.; Gonzalez-Yajimovich, O.; Santamaría-del-Angel, E. Gulf of California geography, geological origins, oceanography, and sedimentation patterns. In Atlas of Coastal Ecosystems in the Western Gulf of California; Johnson, M.E., Ledesma-Vázquez, J., Eds.; University Arizona Press: Tucson, Arizona, 2009; pp. 1–10. [Google Scholar]
- Humphreys, A.M.; Linder, H.P. Concept versus data in delimitation of plant genera. Taxon 2009, 58, 1054–1074. [Google Scholar] [CrossRef]
- Sánchez, D.; Vázquez-Benítez, B.; Vázquez-Sánchez, M.; Aquino, D.; Arias, S. Phylogenetic relationships in Coryphantha and implications on Pelecyphora and Escobaria (Cacteae, Cactoideae, Cactaceae). PhytoKeys 2022, 188, 115–165. [Google Scholar] [CrossRef]
- Korotkova, N.; Aquino, D.; Arias, S.; Eggli, U.; Franck, A.; Gómez-Hinostrosa, C.; Guerrero, P.C.; Hernández, H.M.; Kohlbecker, A.; Köhler, M.; et al. Cactaceae at Caryophyllales.Org- A Dynamic Online Species-Level Taxonomic Backbone for the Family. Willdenowia 2021, 51, 251–270. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Subgenus | Number of Recognized Taxa | Total Number of Analyzed Taxa | Number of Genomes Analyzed | Number of Genomes de novo Sequenced |
|---|---|---|---|---|---|
| 1. Coryphantha | 42 | 10 | 11 | 3 | |
| 1.1 Coryphantha | 26 | 8 | 9 | 2 | |
| 1.2 Neocoryphantha | 15 | 2 | 2 | 1 | |
| 2. Escobaria | NA | 19 | 8 | 9 | 2 |
| 3. Mammillaria | 163 | 70 | 105 | 37 | |
| 3.1. Chilita * | 18 | 16 | 33 | 3 | |
| 3.2. Cochemiea * | 3 | 3 | 10 | 2 | |
| 3.3 Dolichothele | 6 | 2 | 2 | 2 | |
| 3.4. Krainzia | 12 | 5 | 5 | 2 | |
| 3.5. Mammillaria | 117 | 37 | 46 | 26 | |
| 3.6. Mammillopsis | 1 | 1 | 1 | 0 | |
| 3.7. Oehmea | 1 | 1 | 1 | 1 | |
| 3.8. Phellosperma | 5 | 5 | 7 | 1 | |
| 4. Mammilloydia | NA | 1 | 1 | 1 | 1 |
| 5. Neolloydia * | NA | 2 | 2 | 3 | 2 |
| 6. Ortegocactus * | NA | 1 | 1 | 2 | 1 |
| 7. Pelecyphora | NA | 2 | 1 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chincoya, D.A.; Arias, S.; Vaca-Paniagua, F.; Dávila, P.; Solórzano, S. Phylogenomics and Biogeography of the Mammilloid Clade Revealed an Intricate Evolutionary History Arose in the Mexican Plateau. Biology 2023, 12, 512. https://doi.org/10.3390/biology12040512
Chincoya DA, Arias S, Vaca-Paniagua F, Dávila P, Solórzano S. Phylogenomics and Biogeography of the Mammilloid Clade Revealed an Intricate Evolutionary History Arose in the Mexican Plateau. Biology. 2023; 12(4):512. https://doi.org/10.3390/biology12040512
Chicago/Turabian StyleChincoya, Delil A., Salvador Arias, Felipe Vaca-Paniagua, Patricia Dávila, and Sofía Solórzano. 2023. "Phylogenomics and Biogeography of the Mammilloid Clade Revealed an Intricate Evolutionary History Arose in the Mexican Plateau" Biology 12, no. 4: 512. https://doi.org/10.3390/biology12040512
APA StyleChincoya, D. A., Arias, S., Vaca-Paniagua, F., Dávila, P., & Solórzano, S. (2023). Phylogenomics and Biogeography of the Mammilloid Clade Revealed an Intricate Evolutionary History Arose in the Mexican Plateau. Biology, 12(4), 512. https://doi.org/10.3390/biology12040512