
Geometric Morphometric Versus Genomic Patterns in a Large Polyploid Plant Species Complex

 , , ,
, , , 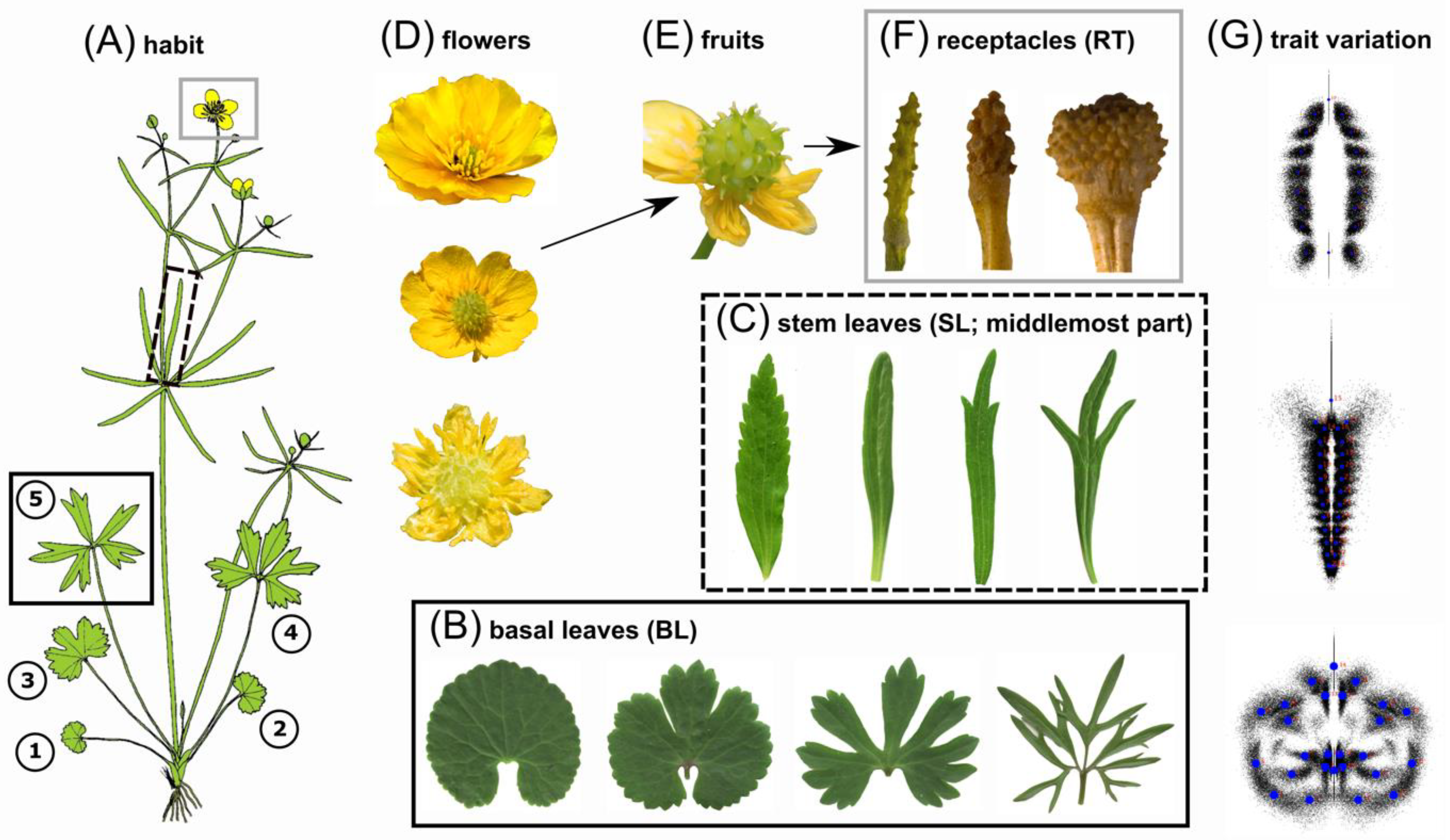{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Locations and Material Sampling
2.2. Genomic and Environmental Data Analysis
2.3. Geometric Morphometric (GM) Data Analysis
2.3.1. Data Collection and Preparation
2.3.2. Digitalization of Traits and Extraction of Shape Variables
2.3.3. Genomic Clusters and Morphological Groups
2.3.4. Covariation of Traits, Taxonomic Resolution, and Shape Changes along Genomic Gradients within Clusters
2.3.5. Shape-Environment and Shape-Genomics Association Models
2.3.6. Ancestral Shape Reconstruction
2.3.7. Inferring Morphological and Genomic Differentiation in an Ecological Context, and Intermediary Versus Transgressive Hybrid Patterns
3. Results
3.1. Morphological Clustering with Genomic Background (RAD-Seq)

3.2. Comparison of Morphological Clustering Concerning Different Genomic Backgrounds

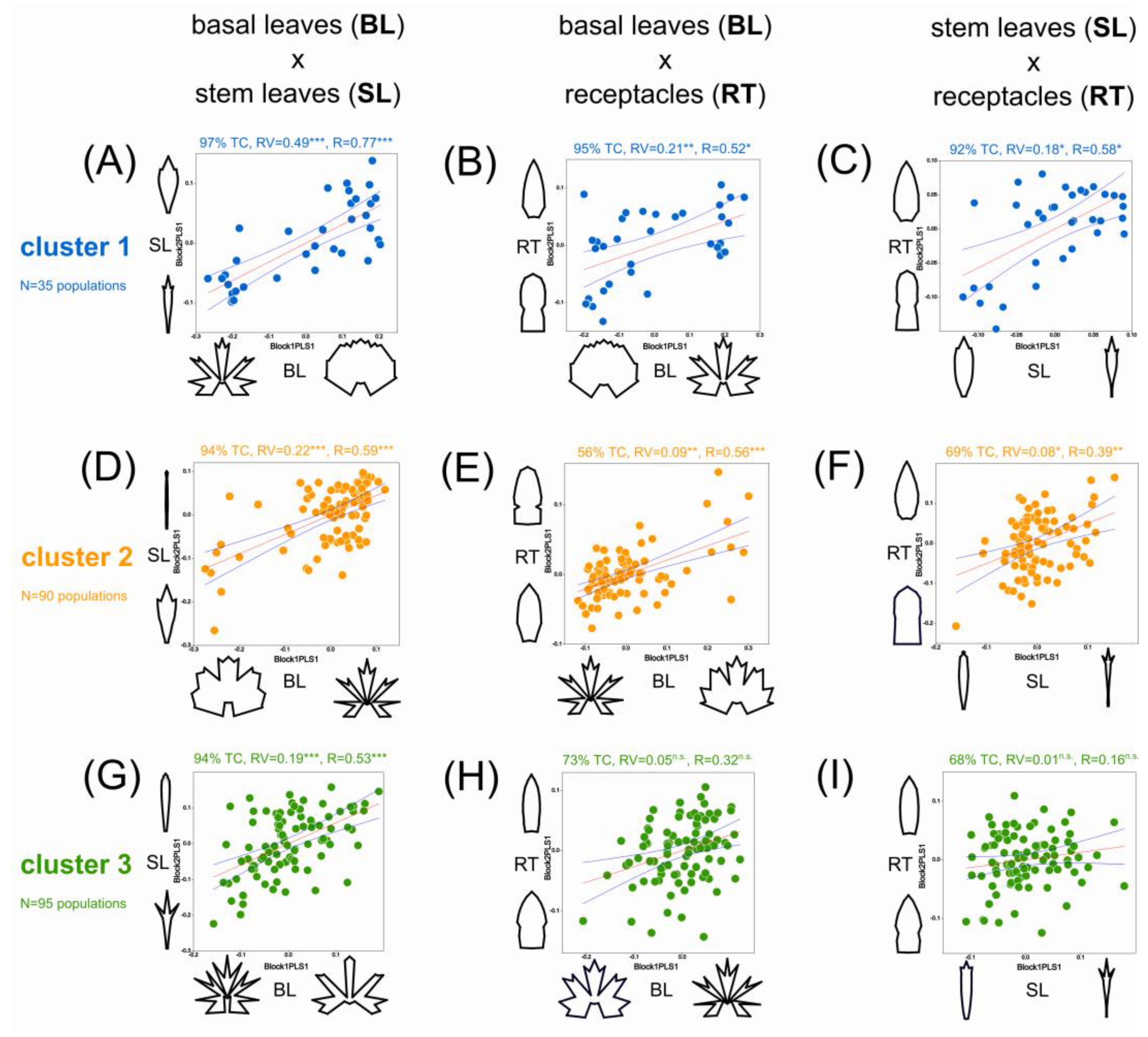
3.3. Covariation of Traits

3.4. Morphological Clustering of Polyploid Apomictic Nothotaxa

3.5. Subgenome Contributions from Sexual Progenitors with Associated Morphotypes of Polyploid Apomicts, and Ancestral Morphotype Reconstruction

3.6. Environmental and Genomic Variables Associated with Phenotypic Variation
3.7. Morphologically and Ecologically Intermediate to Transgressive Polyploid Hybrids
4. Discussion
4.1. GM Methodology
4.2. Congruence of Genetic and Morphological Clustering, and Taxonomical Implications
4.3. Sexual Species and Apomictic Derivative Taxa in Relation to Morphospace and Ecology, and Taxonomic Implications
4.4. Perspectives of GM and Species Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hörandl, E. Novel Approaches for Species Concepts and Delimitation in Polyploids and Hybrids. Plants 2022, 11, 204. [Google Scholar] [CrossRef] [PubMed]
- Soltis, P.S.; Soltis, D.E. The Role of Genetic and Genomic Attributes in the Success of Polyploids. Proc. Natl. Acad. Sci. USA 2000, 97, 7051–7057. [Google Scholar] [CrossRef] [PubMed]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and Genome Evolution in Plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef]
- Leebens-Mack, J.H.; Barker, M.S.; Carpenter, E.J.; Deyholos, M.K.; Gitzendanner, M.A.; Graham, S.W.; Grosse, I.; Li, Z.; Melkonian, M.; Mirarab, S.; et al. One Thousand Plant Transcriptomes and the Phylogenomics of Green Plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. The Role of Hybridization in Plant Speciation. Annu. Rev. Plant Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef]
- Blischak, P.D.; Mabry, M.E.; Conant, G.C.; Pires, J.C. Integrating Networks, Phylogenomics, and Population Genomics for the Study of Polyploidy. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 253–278. [Google Scholar] [CrossRef]
- Comai, L. The Advantages and Disadvantages of Being Polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Van de Peer, Y.; Ashman, T.-L.; Soltis, P.S.; Soltis, D.E. Polyploidy: An Evolutionary and Ecological Force in Stressful Times. Plant Cell 2021, 33, 11–26. [Google Scholar] [CrossRef]
- Hörandl, E. The Complex Causality of Geographical Parthenogenesis. New Phytol. 2006, 171, 525–538. [Google Scholar] [CrossRef]
- Rice, A.; Šmarda, P.; Novosolov, M.; Drori, M.; Glick, L.; Sabath, N.; Meiri, S.; Belmaker, J.; Mayrose, I. The Global Biogeography of Polyploid Plants. Nat. Ecol. Evol. 2019, 3, 265–273. [Google Scholar] [CrossRef]
- Qiu, T.; Liu, Z.; Liu, B. The Effects of Hybridization and Genome Doubling in Plant Evolution via Allopolyploidy. Mol. Biol. Rep. 2020, 47, 5549–5558. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization as an Invasion of the Genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Barke, B.H.; Karbstein, K.; Daubert, M.; Hörandl, E. The Relation of Meiotic Behaviour to Hybridity, Polyploidy and Apomixis in the Ranunculus auricomus Complex (Ranunculaceae). BMC Plant Biol. 2020, 20, 523. [Google Scholar] [CrossRef]
- Izmailow, R. Observations in Embryo and Endosperm Development in Various Chromosomic Types of Apomictic Species Ranunculus cassubicus L. Acta Biol. Cracoviensia Ser. Bot. 1967, 10, 99–111. [Google Scholar]
- Ainouche, M.L.; Baumel, A.; Salmon, A.; Yannic, G. Hybridization, Polyploidy and Speciation in Spartina (Poaceae). New Phytol. 2004, 161, 165–172. [Google Scholar] [CrossRef]
- Barke, B.H.; Daubert, M.; Hörandl, E. Establishment of Apomixis in Diploid F2 Hybrids and Inheritance of Apospory from F1 to F2 Hybrids of the Ranunculus auricomus Complex. Front. Plant Sci. 2018, 9, 1111. [Google Scholar] [CrossRef]
- Hojsgaard, D.; Klatt, S.; Baier, R.; Carman, J.G.; Hörandl, E. Taxonomy and Biogeography of Apomixis in Angiosperms and Associated Biodiversity Characteristics. CRC Crit. Rev. Plant Sci. 2014, 33, 414–427. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, H.; Hörandl, E.; de Oliveira Franca, R.; Wang, L.; Hao, J. Autonomous Apomixis in Praxelis clematidea (Asteraceae: Eupatorieae), an Invasive Alien Plant. AoB Plants 2021, 13, plab007. [Google Scholar] [CrossRef]
- Pellino, M.; Hojsgaard, D.; Schmutzer, T.; Scholz, U.; Hörandl, E.; Vogel, H.; Sharbel, T.F. Asexual Genome Evolution in the Apomictic Ranunculus auricomus Complex: Examining the Effects of Hybridization and Mutation Accumulation. Mol. Ecol. 2013, 22, 5908–5921. [Google Scholar] [CrossRef]
- Hojsgaard, D.; Hörandl, E. The Rise of Apomixis in Natural Plant Populations. Front. Plant Sci. 2019, 10, 358. [Google Scholar] [CrossRef]
- Nogler, G.A. Genetics of Apospory in Apomictic Ranunculus auricomus. V. Conclusion. Bot. Helv. 1984, 94, 411–422. [Google Scholar]
- Ozias-Akins, P.; van Dijk, P.J. Mendelian Genetics of Apomixis in Plants. Annu. Rev. Genet. 2007, 41, 509–537. [Google Scholar] [CrossRef]
- Asker, S.; Jerling, L. Apomixis in Plants; CRC press: Boca Raton, FL, USA, 1992; ISBN 0849345456. [Google Scholar]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Lorberg, E.; Daubert, M.; Hörandl, E. Moving beyond Assumptions: Polyploidy and Environmental Effects Explain a Geographical Parthenogenesis Scenario in European Plants. Mol. Ecol. 2021, 30, 2659–2675. [Google Scholar] [CrossRef] [PubMed]
- Stuessy, T.F.; Crawford, D.J.; Soltis, D.E.; Soltis, P.S. Plant Systematics: The Origin, Interpretation, and Ordering of Plant Biodiversity. In Regnum Vegetabile; Koeltz Scientific Books: Königsstein (Taunus), Germany, 2014; p. 425. ISBN 3874294528. [Google Scholar]
- Miralles, A.; Bruy, T.; Wolcott, K.; Scherz, M.D.; Begerow, D.; Beszteri, B.; Bonkowski, M.; Felden, J.; Gemeinholzer, B.; Glaw, F.; et al. Repositories for Taxonomic Data: Where we are and what is Missing. Syst. Biol. 2020, 69, 1231–1253. [Google Scholar] [CrossRef] [PubMed]
- Small, E. Systematics of Biological Systematics (Or, Taxonomy of Taxonomy). Taxon 1989, 38, 335–356. [Google Scholar] [CrossRef]
- Bebber, D.P.; Carine, M.A.; Wood, J.R.I.; Wortley, A.H.; Harris, D.J.; Prance, G.T.; Davidse, G.; Paige, J.; Pennington, T.D.; Robson, N.K.B.; et al. Herbaria Are a Major Frontier for Species Discovery. Proc. Natl. Acad. Sci. USA 2010, 107, 22169–22171. [Google Scholar] [CrossRef] [PubMed]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-1. Available online: https://www.iucnredlist.org (accessed on 7 March 2023).
- Larsen, B.B.; Miller, E.C.; Rhodes, M.K.; Wiens, J.J. Inordinate Fondness Multiplied and Redistributed: The Number of Species on Earth and the New Pie of Life. Q. Rev. Biol. 2017, 92, 229–265. [Google Scholar] [CrossRef]
- Stuessy, T.F. Plant Taxonomy: The Systematic Evaluation of Comparative Data, 2nd ed.; Columbia University Press: New York, NY, USA, 2009; ISBN 0231518641. [Google Scholar]
- Mallet, J. A Species Definition for the Modern Synthesis. Trends Ecol. Evol. 1995, 10, 294–299. [Google Scholar] [CrossRef]
- De Queiroz, K. Species Concepts and Species Delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef]
- Sukumaran, J.; Knowles, L.L. Multispecies Coalescent Delimits Structure, Not Species. Proc. Natl. Acad. Sci. USA 2017, 114, 1607–1612. [Google Scholar] [CrossRef]
- Sukumaran, J.; Holder, M.T.; Knowles, L.L. Incorporating the Speciation Process into Species Delimitation. PLOS Comput. Biol. 2021, 17, e1008924. [Google Scholar] [CrossRef]
- Ott, T.; Schall, M.; Vogt, R.; Oberprieler, C. The Warps and Wefts of a Polyploidy Complex: Integrative Species Delimitation of the Diploid Leucanthemum (Compositae, Anthemideae) Representatives. Plants 2022, 11, 1878. [Google Scholar] [CrossRef] [PubMed]
- Freudenstein, J.V.; Broe, M.B.; Folk, R.A.; Sinn, B.T. Biodiversity and the Species Concept—Lineages Are Not Enough. Syst. Biol. 2017, 66, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Hörandl, E. The Classification of Asexual Organisms: Old Myths, New Facts, and a Novel Pluralistic Approach. Taxon 2018, 67, 1066–1081. [Google Scholar] [CrossRef]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Dunkel, F.G.; Daubert, M.; Hörandl, E. Phylogenomics Supported by Geometric Morphometrics Reveals Delimitation of Sexual Species within the Polyploid Apomictic Ranunculus auricomus Complex (Ranunculaceae). Taxon 2020, 69, 1191–1220. [Google Scholar] [CrossRef]
- Vences, M.; Miralles, A.; Brouillet, S.; Ducasse, J.; Fedosov, A.; Kharchev, V.; Kostadinov, I.; Kumari, S.; Patmanidis, S.; Scherz, M.D.; et al. ITaxoTools 0.1: Kickstarting a Specimen-Based Software Toolkit for Taxonomists. Megataxa 2021, 6, 77–92. [Google Scholar] [CrossRef]
- Irisarri, I.; Darienko, T.; Pröschold, T.; Fürst-Jansen, J.M.R.; Jamy, M.; De Vries, J. Unexpected Cryptic Species among Streptophyte Algae Most Distant to Land Plants. Proc. R. Soc. B Biol. Sci. 2021, 288, 20212168. [Google Scholar] [CrossRef]
- Chambers, E.A.; Hillis, D.M. The Multispecies Coalescent Over-Splits Species in the Case of Geographically Widespread Taxa. Syst. Biol. 2020, 69, 184–193. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization, Ecological Races and the Nature of Species: Empirical Evidence for the Ease of Speciation. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2971–2986. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative Taxonomy: A Multisource Approach to Exploring Biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef]
- Pinheiro, F.; Dantas-Queiroz, M.V.; Palma-Silva, C. Plant Species Complexes as Models to Understand Speciation and Evolution: A Review of South American Studies. CRC Crit. Rev. Plant Sci. 2018, 37, 54–80. [Google Scholar] [CrossRef]
- Dayrat, B. Towards Integrative Taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- Oberprieler, C. The Wettstein Tesseract: A Tool for Conceptualising Species-rank Decisions and Illustrating Speciation Trajectories. Taxon 2023, 72, 1–7. [Google Scholar] [CrossRef]
- Ennos, R.; French, G.; Hollingsworth, P. Conserving Taxonomic Complexity. Trends Ecol. Evol. 2005, 20, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Federici, S.; Galimberti, A.; Bartolucci, F.; Bruni, I.; De mattia, F.; Cortis, P.; Labra, M. DNA Barcoding to Analyse Taxonomically Complex Groups in Plants: The Case of Thymus (Lamiaceae). Bot. J. Linn. Soc. 2013, 171, 687–699. [Google Scholar] [CrossRef]
- Karbstein, K. Untying Gordian Knots: The Evolution and Biogeography of the Large European Apomictic Polyploid Ranunculus auricomus Plant Complex. Ph.D. Thesis, University of Göttingen, Göttingen, Germany, 2021. [Google Scholar]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Wagner, N.; Marinček, P.; Barke, B.H.; Paetzold, C.; Hörandl, E. Untying Gordian Knots: Unraveling Reticulate Polyploid Plant Evolution by Genomic Data Using the Large Ranunculus auricomus Species Complex. New Phytol. 2022, 235, 2081–2098. [Google Scholar] [CrossRef] [PubMed]
- Sochor, M.; Vašut, R.J.; Sharbel, T.F.; Trávníček, B. How Just a Few Makes a Lot: Speciation via Reticulation and Apomixis on Example of European Brambles (Rubus Subgen. Rubus, Rosaceae). Mol. Phylogenet. Evol. 2015, 89, 13–27. [Google Scholar] [CrossRef]
- Wagner, F.; Ott, T.; Zimmer, C.; Reichhart, V.; Vogt, R.; Oberprieler, C. ‘At the Crossroads towards Polyploidy’: Genomic Divergence and Extent of Homoploid Hybridization Are Drivers for the Formation of the Ox-Eye Daisy Polyploid Complex (Leucanthemum, Compositae-Anthemideae). New Phytol. 2019, 223, 2039–2053. [Google Scholar] [CrossRef]
- Burgess, M.B.; Cushman, K.R.; Doucette, E.T.; Frye, C.T.; Campbell, C.S. Understanding Diploid Diversity: A First Step in Unraveling Polyploid, Apomictic Complexity in Amelanchier. Am. J. Bot. 2015, 102, 2041–2057. [Google Scholar] [CrossRef]
- Chen, Z.J. Genetic and Epigenetic Mechanisms for Gene Expression and Phenotypic Variation in Plant Polyploids. Annu. Rev. Plant Biol. 2007, 58, 377–406. [Google Scholar] [CrossRef]
- Wendel, J.F. The Wondrous Cycles of Polyploidy in Plants. Am. J. Bot. 2015, 102, 1753–1756. [Google Scholar] [CrossRef]
- Spoelhof, J.P.; Soltis, P.S.; Soltis, D.E. Pure Polyploidy: Closing the Gaps in Autopolyploid Research. J. Syst. Evol. 2017, 55, 340–352. [Google Scholar] [CrossRef]
- Liu, B.; Wendel, J.F. Epigenetic Phenomena and the Evolution of Plant Allopolyploids. Mol. Phylogenet. Evol. 2003, 29, 365–379. [Google Scholar] [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and Speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Fox, D.T.; Soltis, D.E.; Soltis, P.S.; Ashman, T.L.; Van de Peer, Y. Polyploidy: A Biological Force from Cells to Ecosystems. Trends Cell Biol. 2020, 30, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Barker, M.S.; Arrigo, N.; Baniaga, A.E.; Li, Z.; Levin, D.A. On the Relative Abundance of Autopolyploids and Allopolyploids. New Phytol. 2016, 210, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Hörandl, E. Morphological Differentiation within the Ranunculus cassubicus Group Compared to Variation of Isozymes, Ploidy Levels, and Reproductive Systems: Implications for Taxonomy. Plant Syst. Evol. 2002, 233, 65–78. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Regnum Vegetabile 159; Koeltz Botanical Books: Glashütten, Germany, 2018; Volume 159, ISBN 9783946583165. [Google Scholar]
- Hörandl, E.; Greilhuber, J.; Klímová, K.; Paun, O.; Temsch, E.; Emadzade, K.; Hodálová, I. Reticulate Evolution and Taxonomic Concepts in the Ranunculus auricomus Complex (Ranunculaceae): Insights from Analysis of Morphological, Karyological and Molecular Data. Taxon 2009, 58, 1194–1215. [Google Scholar] [CrossRef]
- Beck, J.B.; Alexander, P.J.; Allphin, L.; Al-Shehbaz, I.A.; Rushworth, C.; Bailey, C.D.; Windham, M.D. Does Hybridization Drive the Transition to Asexuality in Diploid Boechera? Evolution 2012, 66, 985–995. [Google Scholar] [CrossRef]
- Cushman, K.R.; Burgess, M.B.; Doucette, E.T.; Nelson, G.A.; Campbell, C.S. Species Delimitation in Tetraploid, Apomictic Amelanchier (Rosaceae). Syst. Bot. 2017, 42, 234–256. [Google Scholar] [CrossRef]
- Rothfels, C.J. Polyploid Phylogenetics. New Phytol. 2021, 230, 66–72. [Google Scholar] [CrossRef]
- Gehrke, B.; Bräuchler, C.; Romoleroux, K.; Lundberg, M.; Heubl, G.; Eriksson, T. Molecular Phylogenetics of Alchemilla, Aphanes and Lachemilla (Rosaceae) Inferred from Plastid and Nuclear Intron and Spacer DNA Sequences, with Comments on Generic Classification. Mol. Phylogenet. Evol. 2008, 47, 1030–1044. [Google Scholar] [CrossRef]
- Wagner, N.D.; He, L.; Hörandl, E. Phylogenomic Relationships and Evolution of Polyploid Salix Species Revealed by RAD Sequencing Data. Front. Plant Sci. 2020, 11, 36–41. [Google Scholar] [CrossRef]
- Carter, K.A.; Liston, A.; Bassil, N.V.; Alice, L.A.; Bushakra, J.M.; Sutherland, B.L.; Mockler, T.C.; Bryant, D.W.; Hummer, K.E. Target Capture Sequencing Unravels Rubus Evolution. Front. Plant Sci. 2019, 10, 1615. [Google Scholar] [CrossRef] [PubMed]
- Wagner, F.; Ott, T.; Schall, M.; Lautenschlager, U.; Vogt, R.; Oberprieler, C. Taming the Red Bastards: Hybridisation and Species Delimitation in the Rhodanthemum arundanum-Group (Compositae, Anthemideae). Mol. Phylogenet. Evol. 2020, 144, 106702. [Google Scholar] [CrossRef]
- Pätzold, C.; Wood, K.R.; Eaton, D.A.R.; Wagner, W.L.; Appelhans, M.S. Phylogeny of Hawaiian Melicope (Rutaceae): RAD-Seq Resolves Species Relationships and Reveals Ancient Introgression. Front. Plant Sci. 2019, 10, 1074. [Google Scholar] [CrossRef] [PubMed]
- Šlenker, M.; Kantor, A.; Marhold, K.; Schmickl, R.; Mandáková, T.; Lysak, M.A.; Perný, M.; Caboňová, M.; Slovák, M.; Zozomová-Lihová, J. Allele Sorting as a Novel Approach to Resolving the Origin of Allotetraploids Using Hyb-Seq Data: A Case Study of the Balkan Mountain Endemic Cardamine barbaraeoides. Front. Plant Sci. 2021, 12, 659275. [Google Scholar] [CrossRef]
- Gulyaev, S.; Cai, X.-J.; Guo, F.-Y.; Kikuchi, S.; Applequist, W.L.; Zhang, Z.-X.; Hörandl, E.; He, L. The Phylogeny of Salix Revealed by Whole Genome Re-Sequencing Suggests Different Sex-Determination Systems in Major Groups of the Genus. Ann. Bot. 2022, 129, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Stuessy, T.F.; Mayer, V.; Hörandl, E. (Eds.) Deep Morphology: Toward a Renaissance of Morphology in Plant Systematics; Koeltz Botanical Books; Gantner: Ruggel, Liechtenstein, 2003; ISBN 3-906166-07-4. [Google Scholar]
- Jalas, J. Thymus L. In Flora Europaea, 3; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Eds.; Cambridge University Press: Cambridge, UK, 1972; pp. 172–182. [Google Scholar]
- Morales, R. The History, Botany and Taxonomy of the Genus Thymus. In Thyme—The genus Thymus; Stahl-Biskup, E., Sáez, F., Eds.; Taylor & Francis: London, UK; New York, NY, USA, 2002; pp. 1–43. ISBN 978-0-203-29389-8. [Google Scholar]
- Stace, C.A. Species Recognition in Agamosperms—The Need for a Pragmatic Approach. Folia Geobot. 1998, 33, 319–326. [Google Scholar] [CrossRef]
- Haveman, R. Freakish Patterns—Species and Species Concepts in Apomicts. Nord. J. Bot. 2013, 31, 257–269. [Google Scholar] [CrossRef]
- Scoppola, A.; Angeloni, D.; Franceschini, C. Classical Morphometrics in V. arvensis and V. kitaibeliana (Viola Sect. Melanium) Reveals Intraspecific Variation with Implications for Species Delimitation: Inferences from a Case Study in Central Italy. Plants 2022, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Gratani, L. Plant Phenotypic Plasticity in Response to Environmental Factors. Adv. Bot. 2014, 2014, 208747. [Google Scholar] [CrossRef]
- Karbstein, K.; Tomasello, S.; Prinz, K. Desert-like Badlands and Surrounding (Semi-)Dry Grasslands of Central Germany Promote Small-scale Phenotypic and Genetic Differentiation in Thymus praecox. Ecol. Evol. 2019, 9, 14066–14084. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C. Analyzing Fluctuating Asymmetry with Geometric Morphometrics: Concepts, Methods, and Applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Karbstein, K.; Gockel, S.; Frischbier, N.; Kahlert, K.; Konnert, M.; Profft, I. “High-Altitude Spruces” in Central Europe—A Summarizing Contribution to Phenotypic and (Epi)Genetic Differentiation within Picea abies (L.) H.KARST. Allg. Forst- und Jagdzeitschrift 2021, 191, 197–211. [Google Scholar] [CrossRef]
- Wallace, M.; Bonhomme, V.; Russell, J.; Stillman, E.; George, T.S.; Ramsay, L.; Wishart, J.; Timpany, S.; Bull, H.; Booth, A.; et al. Searching for the Origins of Bere Barley: A Geometric Morphometric Approach to Cereal Landrace Recognition in Archaeology. J. Archaeol. Method Theory 2019, 26, 1125–1142. [Google Scholar] [CrossRef]
- Cardini, A. Geometric Morphometrics. In Encyclopedia of life support systems (EOLSS), Developed under the Auspices of the UNESCO; Eolss Publishers: Paris, France, 2013; Available online: http://www.eolss.net (accessed on 31 January 2023).
- Hodač, L.; Scheben, A.P.; Hojsgaard, D.; Paun, O.; Hörand, E. ITS Polymorphisms Shed Light on Hybrid Evolution in Apomictic Plants: A Case Study on the Ranunculus auricomus Complex. PLoS ONE 2014, 9, 28–30. [Google Scholar] [CrossRef]
- Hodač, L.; Barke, B.H.; Hörandl, E. Mendelian Segregation of Leaf Phenotypes in Experimental F2 Hybrids Elucidates Origin of Morphological Diversity of the Apomictic Ranunculus auricomus Complex. Taxon 2018, 67, 1082–1092. [Google Scholar] [CrossRef]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1997; ISBN 0521585988. [Google Scholar]
- Rohlf, F.J. Morphometric Spaces, Shape Components and the Effects of Linear Transformations. In Advances in Morphometrics; Springer: Boston, MA, USA, 1996; pp. 117–129. [Google Scholar]
- Cooke, S.B.; Terhune, C.E. Form, Function, and Geometric Morphometrics. Anat. Rec. 2015, 298, 5–28. [Google Scholar] [CrossRef]
- Jensen, R.J.; Ciofani, K.M.; Miramontes, L.C. Lines, Outlines, and Landmarks: Morphometric Analyses of Leaves of Acer rubrum, Acer saccharinum (Aceraceae) and Their Hybrid. Taxon 2002, 51, 475–492. [Google Scholar] [CrossRef]
- van der Niet, T.; Zollikofer, C.P.E.; de León, M.S.P.; Johnson, S.D.; Linder, H.P. Three-Dimensional Geometric Morphometrics for Studying Floral Shape Variation. Trends Plant Sci. 2010, 15, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Leménager, M.; Burkiewicz, J.; Schoen, D.J.; Joly, S. Studying Flowers 3D Using Photogrammetry. New Phytol. 2022, 237, 1922–1933. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Evolution and Development of Shape: Integrating Quantitative Approaches. Nat. Rev. Genet. 2010, 11, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Viscosi, V. Geometric Morphometrics and Leaf Phenotypic Plasticity: Assessing Fluctuating Asymmetry and Allometry in European White Oaks (Quercus). Bot. J. Linn. Soc. 2015, 179, 335–348. [Google Scholar] [CrossRef]
- Viscosi, V.; Fortini, P.; Slice, D.E.; Loy, A.; Blasi, C. Geometric Morphometric Analyses of Leaf Variation in Four Oak Species of the Subgenus Quercus (Fagaceae). Plant Biosyst. 2009, 143, 575–587. [Google Scholar] [CrossRef]
- Stojnić, S.; Viscosi, V.; Marković, M.; Ivanković, M.; Orlović, S.; Tognetti, R.; Cocozza, C.; Vasić, V.; Loy, A. Spatial Patterns of Leaf Shape Variation in European Beech (Fagus sylvatica L.) Provenances. Trees 2022, 36, 497–511. [Google Scholar] [CrossRef]
- Kilian, N.; Henning, T.; Plitzner, P.; Müller, A.; Güntsch, A.; Stöver, B.C.; Müller, K.F.; Berendsohn, W.G.; Borsch, T. Sample Data Processing in an Additive and Reproducible Taxonomic Workflow by Using Character Data Persistently Linked to Preserved Individual Specimens. Database 2015, 2015, bav094. [Google Scholar] [CrossRef]
- Piedra-Malagón, E.M.; Albarrán-Lara, A.L.; Rull, J.; Piñero, D.; Sosa, V. Using Multiple Sources of Characters to Delimit Species in the Genus Crataegus (Rosaceae): The Case of the Crataegus rosei Complex. Syst. Biodivers. 2016, 14, 244–260. [Google Scholar] [CrossRef]
- Lexer, C.; Joseph, J.; Van Loo, M.; Prenner, G.; Heinze, B.; Chase, M.W.; Kirkup, D. The Use of Digital Image-Based Morphometrics to Study the Phenotypic Mosaic in Taxa with Porous Genomes. Taxon 2009, 58, 349–364. [Google Scholar] [CrossRef]
- Thompson, K.A.; Urquhart-Cronish, M.; Whitney, K.D.; Rieseberg, L.H.; Schluter, D. Patterns, Predictors, and Consequences of Dominance in Hybrids. Am. Nat. 2021, 197, E72–E88. [Google Scholar] [CrossRef]
- Andrews, K.R.; Good, J.M.; Miller, M.R.; Luikart, G.; Hohenlohe, P.A. Harnessing the Power of RADseq for Ecological and Evolutionary Genomics. Nat. Rev. Genet. 2016, 17, 81–92. [Google Scholar] [CrossRef] [PubMed]
- McKain, M.R.; Johnson, M.G.; Uribe-Convers, S.; Eaton, D.; Yang, Y. Practical Considerations for Plant Phylogenomics. Appl. Plant Sci. 2018, 6, e1038. [Google Scholar] [CrossRef]
- Dauphin, B.; Grant, J.R.; Farrar, D.R.; Rothfels, C.J. Rapid Allopolyploid Radiation of Moonwort Ferns (Botrychium; Ophioglossaceae) Revealed by PacBio Sequencing of Homologous and Homeologous Nuclear Regions. Mol. Phylogenet. Evol. 2018, 120, 342–353. [Google Scholar] [CrossRef]
- Eriksson, J.S.; de Sousa, F.; Bertrand, Y.J.K.; Antonelli, A.; Oxelman, B.; Pfeil, B.E. Allele Phasing Is Critical to Revealing a Shared Allopolyploid Origin of Medicago arborea and M. strasseri (Fabaceae). BMC Evol. Biol. 2018, 18, 9. [Google Scholar] [CrossRef]
- Lautenschlager, U.; Wagner, F.; Oberprieler, C. AllCoPol: Inferring Allele Co-Ancestry in Polyploids. BMC Bioinform. 2020, 21, 441. [Google Scholar] [CrossRef] [PubMed]
- Rannala, B.; Yang, Z. Bayes Estimation of Species Divergence Times and Ancestral Population Sizes Using DNA Sequences from Multiple Loci. Genetics 2003, 164, 1645–1656. [Google Scholar] [CrossRef]
- Yang, Z.; Rannala, B. Unguided Species Delimitation Using DNA Sequence Data from Multiple Loci. Mol. Biol. Evol. 2014, 31, 3125–3135. [Google Scholar] [CrossRef]
- Weitemier, K.; Straub, S.C.K.; Cronn, R.C.; Fishbein, M.; Schmickl, R.; McDonnell, A.; Liston, A. Hyb-Seq: Combining Target Enrichment and Genome Skimming for Plant Phylogenomics. Appl. Plant Sci. 2014, 2, 1400042. [Google Scholar] [CrossRef]
- Folk, R.A.; Mandel, J.R.; Freudenstein, J.V. A Protocol for Targeted Enrichment of Intron-Containing Sequence Markers for Recent Radiations: A Phylogenomic Example from Heuchera (Saxifragaceae). Appl. Plant Sci. 2015, 3, 1500039. [Google Scholar] [CrossRef] [PubMed]
- Stull, G.W.; Soltis, P.S.; Soltis, D.E.; Gitzendanner, M.A.; Smith, S.A. Nuclear Phylogenomic Analyses of Asterids Conflict with Plastome Trees and Support Novel Relationships among Major Lineages. Am. J. Bot. 2020, 107, 790–805. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.I.; Hefer, C.A.; Kolosova, N.; Douglas, C.J.; Cronk, Q.C.B. Whole Plastome Sequencing Reveals Deep Plastid Divergence and Cytonuclear Discordance between Closely Related Balsam Poplars, Populus balsamifera and P. trichocarpa (Salicaceae). N. Phytol. 2014, 204, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Paetzold, C.; Barke, B.H.; Hörandl, E. Evolution of Transcriptomes in Early-Generation Hybrids of the Apomictic Ranunculus auricomus Complex (Ranunculaceae). Int. J. Mol. Sci. 2022, 23, 13881. [Google Scholar] [CrossRef] [PubMed]
- Paun, O.; Stuessy, T.F.; Hörandl, E. The Role of Hybridization, Polyploidization and Glaciation in the Origin and Evolution of the Apomictic Ranunculus cassubicus Complex. New Phytol. 2006, 171, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Emadzade, K.; Gehrke, B.; Peter Linder, H.; Hörandl, E. The Biogeographical History of the Cosmopolitan Genus ranunculus L. (Ranunculaceae) in the Temperate to Meridional Zones. Mol. Phylogenet. Evol. 2011, 58, 4–21. [Google Scholar] [CrossRef]
- Emadzade, K.; Lebmann, M.J.; Hoffmann, M.H.; Tkach, N.; Lone, F.A.; Hörandl, E. Phylogenetic Relationships and Evolution of High Mountain Buttercups (Ranunculus) in North America and Central Asia. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 131–141. [Google Scholar] [CrossRef]
- Jalas, J.; Suominen, J. Atlas Florae Europaeae, Vol. 8, Nymphaeaceae to Ranunculaceae; The committee for mapping the flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1989. [Google Scholar]
- GBIF. Secretariat 2017. Ranunculus auricomus L; GBIF Backbone Taxonomy. Checklist Dataset. Available online: https://doi.org/10.15468/39omei (accessed on 21 November 2018).
- Melzheimer, V.; Hörandl, E. Die Ranunculaceae der Flora von Zentraleuropa: Ranunculus sect. Auricomus; Universitätsbibliothek Johann Christian Senckenberg: Frankfurt, Germany, 2022. [Google Scholar]
- Borchers-Kolb, E. Ranunculus sect. Auricomus in Bayern und den angrenzenden Gebieten II. Spezieller Teil. Mitt. Bot. München 1985, 21, 49–300. [Google Scholar]
- Marklund, G. Der Ranunculus auricomus-Komplex in Finnland I. Diagnosen und Fundortslisten einiger Sippen des R. auricomus L. Coll. (s. str.). Flora Fenn. 1961, 3, 1–128. [Google Scholar]
- Jalas, J.; Kvist, G. Atlas Florae Europaeae Notes. 7. Taxa of the Ranunculus auricomus Group Recognized as Species by Marklund in 1940. Ann. Bot. Fenn. 1987, 12, 221–223. [Google Scholar]
- Hörandl, E.; Raab-Straube, E. Ranunculaceae. Available online: http://ww2.bgbm.org/EuroPlusMed (accessed on 7 March 2023).
- Leslie, A.C. Ranunculaceae. Section 2. Auricomus. In Flora of Great Britain and Ireland; Sell, P., Murrell, G., Eds.; Cambridge University Press: Cambridge, UK, 2018; pp. 671–677. [Google Scholar]
- Julin, E. Ranunculus auricomus L. in Södermanland, East-Central Sweden. Opera Bot. 1980, 57, 1–145. [Google Scholar]
- Hörandl, E.; Gutermann, W. Der Ranunculus auricomus-Komplex in Österreich 2. Die R. cassubicus-, R. monophyllus- und R. fallax-Sammelgruppe. Bot. Jahrb. Syst. 1998, 120, 545–598. [Google Scholar]
- Dunkel, F.G.; Gregor, T.; Paule, J. New Diploid Species in the Ranunculus auricomus Complex (Ranunculaceae) from W and SE Europe. Willdenowia 2018, 48, 227–257. [Google Scholar] [CrossRef]
- Dunkel, F.G. Contribution to the Knowledge of the Ranunculus auricomus Complex (Ranunculaceae) in Spain. Stapfia 2021, 112, 5–59. [Google Scholar]
- Linnaeus, C. Species Plantarum; Holmiae: Uppsala, Sweden, 1753. [Google Scholar]
- Kvist, G. Identity of the Material of the Ranunculus auricomus Group in the Linnaean Herbarium. Ann. Bot. Fenn. 1987, 24, 73–77. [Google Scholar]
- Sloboda, D. Rostlinnictví: Čili návod k snadnému určení a pojmenování rostlin v Čechách, Moravě a jiných zemích Rakouského Mocnářství domácích; V kommissí u Fr. Rivnáce: Prague, Bohemia (Austrian Empire), 1852. [Google Scholar]
- Ovchinnikov, P. Ranunculus monophyllus Ovcz. in Bot. Mater. Gerb. Glavn. Bot. Sada RSFSR. 1922, 3, 54. Available online: https://www.europlusmed.org/cdm_dataportal/taxon/c2da9f70-7f7d-4320-95bd-425566191fa8 (accessed on 31 January 2023).
- Marklund, G. Der Ranunculus auricomus-Komplex in Finnland II: Diagnosen und Fundortslisten einiger Sippen von R. fallax (W. et Gr.) Schur, R. monophyllus Ovcz. und R. cassubicus L. Flora Fenn. 1965, 4, 1–198. [Google Scholar]
- Soó, R. Die Ranunculus auricomus L. Emend. Korsh. Artengruppe in der Flora Ungarns und der Karpaten I. Acta Bot. Acad. Sci. Hung 1964, 10, 221–237. [Google Scholar]
- Schwarz, O. Beiträge zur Kenntnis kritischer Formenkreise im Gebiet der Flora von Thüringen. Mitteilungen der Thüringischen Bot. Gesellschaft 1949, 1, 120–143. [Google Scholar]
- Hörandl, E.; Gutermann, W. Zur Kenntnis des Ranunculus auricomus-Komplexes in Österreich: Die Arten der R. phragmiteti- und R. indecorus-Gruppe. Phyton 1998, 37, 263–320. [Google Scholar]
- Hörandl, E.; Gutermann, W. Der Ranunculus auricomus-Komplex in Österreich und benachbarten Gebieten 3. Die Arten der R. latisectus-, R. puberulus-, R. stricticaulis-und R. argoviensis (R. auricomus-Sammelgruppe). Bot. Jahrb. Syst. 1999, 121, 99–138. [Google Scholar]
- Hörandl, E. Species Concepts in Agamic Complexes: Applications in the Ranunculus auricomus Complex and General Perspectives. Folia Geobot. 1998, 33, 335–348. [Google Scholar] [CrossRef]
- Ericsson, S. The Microspecies of the Ranunculus auricomus Complex Treated at the Species Level. Ann. Bot. Fenn. 1992, 29, 123–158. [Google Scholar]
- Jäger, E. Ranunculus L. In Rothmaler Exkursionsflora von Deutschland. Gefäßpflanzen: Grundband 2; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Borchers-Kolb, E. Ranunculus sect. Auricomus in Bayern und den angrenzenden Gebieten—I. Allgemeiner Teil. Mitt. Bot. München 1983, 19, 363–429. [Google Scholar]
- Tomasello, S.; Karbstein, K.; Hodač, L.; Pätzold, C.; Hörandl, E. Phylogenomics Unravels Quaternary Vicariance and Allopatric Speciation Patterns in Temperate-montane Plant Species: A Case Study on the Ranunculus auricomus Species Complex. Mol. Ecol. 2020, 29, 2031–2049. [Google Scholar] [CrossRef]
- Hörandl, E.; Gutermann, W. Die Bearbeitung der Ranunculus auricomus-Gruppe für die “Flora von Österreich”–Ein Werkstattbericht. Fl. Austr. Novit 1995, 2, 12–27. [Google Scholar] [CrossRef]
- Paun, O.; Greilhuber, J.; Temsch, E.M.; Hörandl, E. Patterns, Sources and Ecological Implications of Clonal Diversity in Apomictic Ranunculus carpaticola (Ranunculus auricomus Complex, Ranunculaceae). Mol. Ecol. 2006, 15, 897–910. [Google Scholar] [CrossRef]
- Reichenbach, L. Flora Germanica excurso-ria […]; Cnobloch, C., Ed.; Carolum Cnobloch: Leipzig, Germany, 1832. [Google Scholar]
- Grau, J. Vorläufige Übersicht der iberischen Vertreter von Ranunculus sect. auricomus. Mitt. Bot. München 1984, 20, 11–28. [Google Scholar]
- Masci, S.; Miho, A.; Marchi, P. Ranunculus auricomus L. Aggr. (Ranunculaceae) in Italy. I. Sexual Tetraploids on the Apennines. Caryologia 1994, 47, 97–108. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Willis, J.H. Plant Speciation. Science 2007, 317, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Dittrich-Reed, D.R.; Fitzpatrick, B.M. Transgressive Hybrids as Hopeful Monsters. Evol. Biol. 2013, 40, 310–315. [Google Scholar] [CrossRef]
- Frichot, E.; Mathieu, F.; Trouillon, T.; Bouchard, G.; François, O. Fast and Efficient Estimation of Individual Ancestry Coefficients. Genetics 2014, 196, 973–983. [Google Scholar] [CrossRef]
- Frichot, E.; François, O. LEA: An R Package for Landscape and Ecological Association Studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Malinsky, M.; Trucchi, E.; Lawson, D.J.; Falush, D. RADpainter and FineRADstructure: Population Inference from RADseq Data. Mol. Biol. Evol. 2018, 35, 1284–1290. [Google Scholar] [CrossRef]
- Jones, G. Algorithmic Improvements to Species Delimitation and Phylogeny Estimation under the Multispecies Coalescent. J. Math. Biol. 2017, 74, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Taiyun, W.; Simko, V. Corrplot—Visualization of a Correlation Matrix. 2022. Available online: https://cran.r-project.org/web/packages/corrplot/corrplot (accessed on 31 January 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.r-project.org (accessed on 7 March 2023).
- Volkova, P.A.; Kasatskaya, S.A.; Boiko, A.A.; Shipunov, A.B. Stability of Leaf Form and Size during Specimen Preparation of Herbarium Specimens. Feddes Repert. 2010, 121, 219–225. [Google Scholar] [CrossRef]
- Lohwasser, U. Biosystematische Untersuchungen an Ranunculus auricomus L. (Ranunculaceae) in Deutschland. Diss. Bot. 2001, 343, 220. [Google Scholar]
- Hörandl, E.; Gutermann, W. Der Ranunculus auricomus-Komplex in Österreich. 1. Methodik. Gruppierung der mitteleuropäischen Sippen. Bot. Jahrb. Syst. 1998, 120, 1–44. [Google Scholar]
- Rohlf, F.J. The Tps Series of Software. Hystrix 2015, 26, 9–12. [Google Scholar] [CrossRef]
- Klingenberg, C.P. MorphoJ: An Integrated Software Package for Geometric Morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar] [CrossRef]
- Karbstein, K.; Prinz, K.; Hellwig, F.; Römermann, C. Plant Intraspecific Functional Trait Variation Is Related to Within-habitat Heterogeneity and Genetic Diversity in Trifolium montanum L. Ecol. Evol. 2020, 10, 5015–5033. [Google Scholar] [CrossRef] [PubMed]
- Goslee, S.C.; Urban, D.L. The Ecodist Package for Dissimilarity-Based Analysis of Ecological Data. J. Stat. Softw. 2007, 22, 1–19. [Google Scholar] [CrossRef]
- Maddison, W.P. Squared-Change Parsimony Reconstructions of Ancestral States for Continuous-Valued Characters on a Phylogenetic Tree. Syst. Biol. 1991, 40, 304–314. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Gidaszewski, N.A. Testing and Quantifying Phylogenetic Signals and Homoplasy in Morphometric Data. Syst. Biol. 2010, 59, 245–261. [Google Scholar] [CrossRef] [PubMed]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. arXiv 2018, arXiv:1802.03426. [Google Scholar]
- Hörandl, E.; Jakubowsky, G.; Dobeš, C. Isozyme and Morphological Diversity within Apomictic and Sexual Taxa of the Ranunculus auricomus Complex. Plant Syst. Evol. 2001, 226, 165–185. [Google Scholar] [CrossRef]
- Jiang, X.; Hipp, A.L.; Deng, M.; Su, T.; Zhou, Z.; Yan, M. East Asian Origins of European Holly Oaks (Quercus Section Ilex Loudon) via the Tibet-Himalaya. J. Biogeogr. 2019, 46, 2203–2214. [Google Scholar] [CrossRef]
- Kirschner, J.; Záveská Drábková, L.; Štěpánek, J.; Uhlemann, I. Towards a Better Understanding of the Taraxacum Evolution (Compositae–Cichorieae) on the Basis of nrDNA of Sexually Reproducing Species. Plant Syst. Evol. 2015, 301, 1135–1156. [Google Scholar] [CrossRef]
- Alger, E.I.; Edger, P.P. One Subgenome to Rule Them All: Underlying Mechanisms of Subgenome Dominance. Curr. Opin. Plant Biol. 2020, 54, 108–113. [Google Scholar] [CrossRef]
- Ericsson, S. Microspecies within the Ranunculus auricomus Complex. In Flora Nordica 2; Jonsell, B., Ed.; Bergius Foundation: Stockholm, Sweden, 2001; pp. 382–397. [Google Scholar]
- Kürbs, S. Vergleichend-Entwicklungsgeschichtliche Studien an Ranunculaceaen-Fiederblättern. Teil I, II. Bot. Jahrb. Syst. 1973, 93, 130–167. [Google Scholar]
- Grant, V. Plant Speciation; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Hörandl, E. The Evolution of Self-Fertility in Apomictic Plants. Sex. Plant Reprod. 2010, 23, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Hörandl, E.; Greilhuber, J. Diploid and Autotetraploid Sexuals and Their Relationships to Apomicts in the Ranunculus cassubicus Group: Insights from DNA Content and Isozyme Variation. Plant Syst. Evol. 2002, 234, 85–100. [Google Scholar] [CrossRef]
- Sailer, C.; Schmid, B.; Grossniklaus, U. Apomixis Allows the Transgenerational Fixation of Phenotypes in Hybrid Plants. Curr. Biol. 2016, 26, 331–337. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Li, M.; Chitwood, D.H.; Myles, S. Morphometrics Reveals Complex and Heritable Apple Leaf Shapes. Front. Plant Sci. 2018, 8, 2185. [Google Scholar] [CrossRef]
- Tsukaya, H. Leaf Shape: Genetic Controls and Environmental Factors. Int. J. Dev. Biol. 2005, 49, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Peppe, D.J.; Royer, D.L.; Cariglino, B.; Oliver, S.Y.; Newman, S.; Leight, E.; Enikolopov, G.; Fernandez-Burgos, M.; Herrera, F.; Adams, J.M.; et al. Sensitivity of Leaf Size and Shape to Climate: Global Patterns and Paleoclimatic Applications. New Phytol. 2011, 190, 724–739. [Google Scholar] [CrossRef]
- Wang, H.; Wang, R.; Harrison, S.P.; Prentice, I.C. Leaf Morphological Traits as Adaptations to Multiple Climate Gradients. J. Ecol. 2022, 110, 1344–1355. [Google Scholar] [CrossRef]
- Halamski, A.T.; Dunkel, F.G.; Temsch, E.M. Ranunculus kobendzae, a New Species of the R. auricomus Complex (Ranunculaceae) from Poland. BioOne 2019, 56, 123–134. [Google Scholar] [CrossRef]
- Hörandl, E. Geographical Parthenogenesis in Alpine and Arctic Plants. Plants 2023, 12, 844. [Google Scholar] [CrossRef]
- Wäldchen, J.; Mäder, P. Plant Species Identification Using Computer Vision Techniques: A Systematic Literature Review. Arch. Comput. Methods Eng. 2018, 25, 507–543. [Google Scholar] [CrossRef]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep Learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Rzanny, M.; Mäder, P.; Deggelmann, A.; Chen, M.; Wäldchen, J. Flowers, Leaves or Both? How to Obtain Suitable Images for Automated Plant Identification. Plant Methods 2019, 15, 77. [Google Scholar] [CrossRef] [PubMed]
- Mäder, P.; Boho, D.; Rzanny, M.; Seeland, M.; Wittich, H.C.; Deggelmann, A.; Wäldchen, J. The Flora Incognita App—Interactive Plant Species Identification. Methods Ecol. Evol. 2021, 12, 1335–1342. [Google Scholar] [CrossRef]
- Weaver, W.N.; Ng, J.; Laport, R.G. LeafMachine: Using Machine Learning to Automate Leaf Trait Extraction from Digitized Herbarium Specimens. Appl. Plant Sci. 2020, 8, e11367. [Google Scholar] [CrossRef] [PubMed]
- Katal, N.; Rzanny, M.; Mäder, P.; Wäldchen, J. Deep Learning in Plant Phenological Research: A Systematic Literature Review. Front. Plant Sci. 2022, 13, 805738. [Google Scholar] [CrossRef]
- Cintas, C.; Quinto-Sánchez, M.; Acuña, V.; Paschetta, C.; de Azevedo, S.; Cesar Silva de Cerqueira, C.; Ramallo, V.; Gallo, C.; Poletti, G.; Bortolini, M.C.; et al. Automatic Ear Detection and Feature Extraction Using Geometric Morphometrics and Convolutional Neural Networks. IET Biom. 2017, 6, 211–223. [Google Scholar] [CrossRef]
- Bellin, N.; Calzolari, M.; Callegari, E.; Bonilauri, P.; Grisendi, A.; Dottori, M.; Rossi, V. Geometric Morphometrics and Machine Learning as Tools for the Identification of Sibling Mosquito Species of the Maculipennis complex (Anopheles). Infect. Genet. Evol. 2021, 95, 105034. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Yravedra, J.; Huguet, R.; Aramendi, J.; Maté-González, M.Á.; González-Aguilera, D.; Arriaza, M.C. Combining Machine Learning Algorithms and Geometric Morphometrics: A Study of Carnivore Tooth Marks. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 522, 28–39. [Google Scholar] [CrossRef]
- Le, V.-L.; Beurton-Aimar, M.; Zemmari, A.; Marie, A.; Parisey, N. Automated Landmarking for Insects Morphometric Analysis Using Deep Neural Networks. Ecol. Inform. 2020, 60, 101175. [Google Scholar] [CrossRef]
- Rzanny, M.; Wittich, H.C.; Mäder, P.; Deggelmann, A.; Boho, D.; Wäldchen, J. Image-Based Automated Recognition of 31 Poaceae Species: The Most Relevant Perspectives. Front. Plant Sci. 2022, 12, 3315. [Google Scholar] [CrossRef] [PubMed]
- Selvaraju, R.R.; Cogswell, M.; Das, A.; Vedantam, R.; Parikh, D.; Batra, D. Grad-CAM: Visual Explanations from Deep Networks via Gradient-Based Localization. In Proceedings of the IEEE International Conference on Computer Vision, Venice, Italy, 22–29 October 2017; pp. 618–626. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodač, L.; Karbstein, K.; Tomasello, S.; Wäldchen, J.; Bradican, J.P.; Hörandl, E. Geometric Morphometric Versus Genomic Patterns in a Large Polyploid Plant Species Complex. Biology 2023, 12, 418. https://doi.org/10.3390/biology12030418
Hodač L, Karbstein K, Tomasello S, Wäldchen J, Bradican JP, Hörandl E. Geometric Morphometric Versus Genomic Patterns in a Large Polyploid Plant Species Complex. Biology. 2023; 12(3):418. https://doi.org/10.3390/biology12030418
Chicago/Turabian StyleHodač, Ladislav, Kevin Karbstein, Salvatore Tomasello, Jana Wäldchen, John Paul Bradican, and Elvira Hörandl. 2023. "Geometric Morphometric Versus Genomic Patterns in a Large Polyploid Plant Species Complex" Biology 12, no. 3: 418. https://doi.org/10.3390/biology12030418
APA StyleHodač, L., Karbstein, K., Tomasello, S., Wäldchen, J., Bradican, J. P., & Hörandl, E. (2023). Geometric Morphometric Versus Genomic Patterns in a Large Polyploid Plant Species Complex. Biology, 12(3), 418. https://doi.org/10.3390/biology12030418