
Admittance to Wildlife Rehabilitation Centres Points to Adverse Effects of Climate Change on Insectivorous Bats

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
- In summer, the number of individual bats from urban colonies admitted to WRCs will be influenced by the mean maximum temperature of the week they were rescued, and higher numbers are predicted in hotter weeks.
- Being more sensitive and less capable of moving autonomously inside a roost to select lower temperatures, young bats will be more exposed to heat stress than adults, so we predict that admittance data will be dominated by pups.
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perkins, S.E.; Alexander, L.V. On the measurement of heat waves. J. Clim. 2013, 26, 4500–4517. [Google Scholar] [CrossRef]
- Montoya, J.M.; Raffaelli, D. Climate change, biotic interactions and ecosystem services. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2013–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, J.; Møller, A.P. Extreme climatic events in relation to global change and their impact on life histories. Curr. Zool. 2011, 57, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festa, F.; Ancillotto, L.; Santini, L.; Pacifici, M.; Rocha, R.; Toshkova, N.; Amorim, F.; Benitez-Lopez, A.; Domer, A.; Hamidovic, D.; et al. Bat responses to climate change: A systematic review. Biol. Rev. 2022, 98, 19–33. [Google Scholar] [CrossRef]
- Ancillotto, L.; Santini, L.; Ranc, N.; Maiorano, L.; Russo, D. Extraordinary range expansion in a common bat: The potential roles of climate change and urbanisation. Sci. Nat. 2016, 103, 1–8. [Google Scholar] [CrossRef]
- Ancillotto, L.; Budinski, I.; Nardone, V.; Di Salvo, I.; Della Corte, M.; Bosso, L.; Conti, P.; Russo, D. What is driving range expansion in a common bat? Hints from thermoregulation and habitat selection. Behav. Process. 2018, 157, 540–546. [Google Scholar] [CrossRef]
- Sherwin, H.A.; Montgomery, W.I.; Lundy, M.G. The impact and implications of climate change for bats. Mamm. Rev. 2013, 43, 171–182. [Google Scholar] [CrossRef]
- Geiser, F. Evolution of daily torpor and hibernation in birds and mammals: Importance of body size. Clin. Exp. Pharmacol. Physiol. 1998, 25, 736–740. [Google Scholar] [CrossRef]
- Altringham, J.D. Bats: From Evolution to Conservation; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Welman, S.; Tuen, A.A.; Lovegrove, B.G. Using thermoregulatory profiles to assess climate change vulnerability in an arboreal tropical bat: Heterothermy may be a pre-adaptive advantage. Clim. Res. 2017, 74, 161–170. [Google Scholar] [CrossRef]
- Korine, C.; Adams, R.; Russo, D.; Fisher-Phelps, M.; Jacobs, D. Bats and water: Anthropogenic alterations threaten global bat populations. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer: Cham, Switzerland, 2016; pp. 215–241. [Google Scholar]
- Salinas-Ramos, V.B.; Ancillotto, L.; Bosso, L.; Sánchez-Cordero, V.; Russo, D. Interspecific competition in bats: State of knowledge and research challenges. Mamm. Rev. 2020, 50, 68–81. [Google Scholar] [CrossRef]
- O’Shea, T.J.; Cryan, P.M.; Hayman, D.T.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mamm. Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Welbergen, J.A.; Klose, S.M.; Markus, N.; Eby, P. Climate change and the effects of temperature extremes on Australian flying foxes. Proc. R. Soc. B 2008, 275, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Hale, R.; Swearer, S.E. Ecological traps: Current evidence and future directions. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruvot, M.; Cappelle, J.; Furey, N.; Hul, V.; Heng, H.S.; Duong, V.; Dussart, P.; Horwood, P. Extreme temperature event and mass mortality of insectivorous bats. Eur. J. Wildl. Res. 2019, 65, 41. [Google Scholar] [CrossRef]
- Kerth, G.; Weissmann, K.; Konig, B. Day roost selection in female Bechstein’s bats (Myotis bechsteinii): A field experiment to determine the influence of roost temperature. Oecologia 2001, 126, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lausen, C.L.; Barclay, R.M.R. Benefits of living in a building: Big brown bats (Eptesicus fuscus) in rocks versus buildings. J. Mammal. 2006, 87, 362–370. [Google Scholar] [CrossRef] [Green Version]
- Ransome, R.D. The Impact of Maternity Roost Conditions on Populations of Greater Horseshoe Bats; English Nature: Peterborough, UK, 1998. [Google Scholar]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef]
- Martin Bideguren, G.; López-Baucells, A.; Puig-Montserrat, X.; Mas, M.; Porres, X.; Flaquer, C. Bat boxes and climate change: Testing the risk of over-heating in the Mediterranean region. Biodivers. Conserv. 2019, 28, 21–35. [Google Scholar] [CrossRef]
- Bat Conservation Trust. Available online: https://www.bats.org.uk/news/2020/07/helpline-heroes-rescue-63-baby-bats-struck-by-heat-wave (accessed on 4 November 2022).
- The Guardian. Groc, I. 2002. Available online: https://www.theguardian.com/environment/2021/feb/24/race-to-save-bats-flying-foxes-from-overheating-as-temperatures-rise-aoe (accessed on 4 November 2022).
- Ancillotto, L.; Fichera, G.; Pidinchedda, E.; Veith, M.; Kiefer, A.; Mucedda, M.; Russo, D. Wildfires, heatwaves and human disturbance threaten insular endemic bats. Biodivers. Conserv. 2021, 30, 4401–4416. [Google Scholar] [CrossRef]
- Czenze, Z.J.; Noakes, M.J.; Wojciechowski, M.S. Home is where the heat is: Thermoregulation of European bats inhabiting artificial roosts and the threat of heat waves. J. Appl. Ecol. 2022, 59, 2179–2188. [Google Scholar] [CrossRef]
- Bondarenco, A.; Körtner, G.; Geiser, F. Hot bats: Extreme thermal tolerance in a desert heat wave. Naturwissenschaften 2014, 101, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, G.S.; Brunet-Rossinni, A.K. Methods for age estimation and the study of senescence in bats. In Ecological and Behavioral Methods for the Study of Bats; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 315–325. [Google Scholar]
- Smeraldo, S.; Bosso, L.; Salinas-Ramos, V.B.; Ancillotto, L.; Sánchez-Cordero, V.; Gazaryan, S.; Russo, D. Generalists yet different: Distributional responses to climate change may vary in opportunistic bat species sharing similar ecological traits. Mamm. Rev. 2021, 51, 571–584. [Google Scholar] [CrossRef]
- Moretti, M.; Maddalena, T.; Pierallini, R.; Violani, C. What type of attention should be paid to the most abundant anthropophilous species? Considerations on the example of Pipistrellus c. pipistrellus, P. kuhlii and Hypsugo savii on the southern slopes of the Alps (Ticino, Switzerland). In Atti del I Convegno Italiano sui Chirotteri; Dondini, G., Papalini, O., Vergari, S., Eds.; Castell’Azzara: Grosseto, Italy, 1998; pp. 211–222. [Google Scholar]
- Andreani, G.; Cannavacciuolo, A.; Menotta, S.; Spallucci, V.; Fedrizzi, G.; Carpene, E.; Isani, G. Environmental exposure to non-essential trace elements in two bat species from urbanised (Tadarida teniotis) and open land (Miniopterus schreibersii) areas in Italy. Environ. Pollut. 2019, 254, 113034. [Google Scholar] [CrossRef] [PubMed]
- Flaquer, C.; Puig-Montserrat, X.; López-Baucells, A.; Torre, I.; Freixas, L.; Mas, M.; Porres, X.; Arrizabalaga, A. Could overheating turn bat boxes into death traps? Barbastella 2014, 7, 46–53. [Google Scholar] [CrossRef]
- Korine, C.; Cohen, Y.; Kahnonitch, I. Insect Pest Pheromone Lures May Enhance the Activity of Insectivorous Bats in Mediterranean Vineyards and Apple Orchards. Sustainability 2022, 14, 16566. [Google Scholar] [CrossRef]
- Lausen, C.L.; Barclay, R.M. Roosting behaviour and roost selection of female big brown bats (Eptesicus fuscus) roosting in rock crevices in southeastern Alberta. Can. J. Zool. 2002, 80, 1069–1076. [Google Scholar] [CrossRef]
- Lewis, S.E. Roost fidelity of bats: A review. J. Mammal. 1995, 76, 481–496. [Google Scholar] [CrossRef]
- Racey, P.A.; Swift, S.M. Variations in gestation length in a colony of pipistrelle bats (Pipistrellus pipistrellus) from year to year. Reproduction 1981, 61, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Wilde, C.J.; Knight, C.H.; Racey, P.A. Influence of torpor on milk protein composition and secretion in lactating bats. J. Exp. Zool. 1999, 284, 35–41. [Google Scholar] [CrossRef]
- McAllan, B.M.; Geiser, F. Torpor during reproduction in mammals and birds: Dealing with an energetic conundrum. ICB 2014, 54, 516–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serangeli, M.T.; Cistrone, L.; Ancillotto, L.; Tomassini, A.; Russo, D. The post-release fate of hand-reared orphaned bats: Survival and habitat selection. Anim. Welf. UFAW J. 2012, 21, 9. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.; Coleman, J.L.; Ancillotto, L.; Korine, C. Ecosystem Services by Bats in Urban Areas. In Urban Bats, Fascinating Life Sciences; Moretto, L., Coleman, J.L., Davy, C.M., Fenton, M.B., Korine, C., Patriquin, K.J., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 167–181. [Google Scholar]



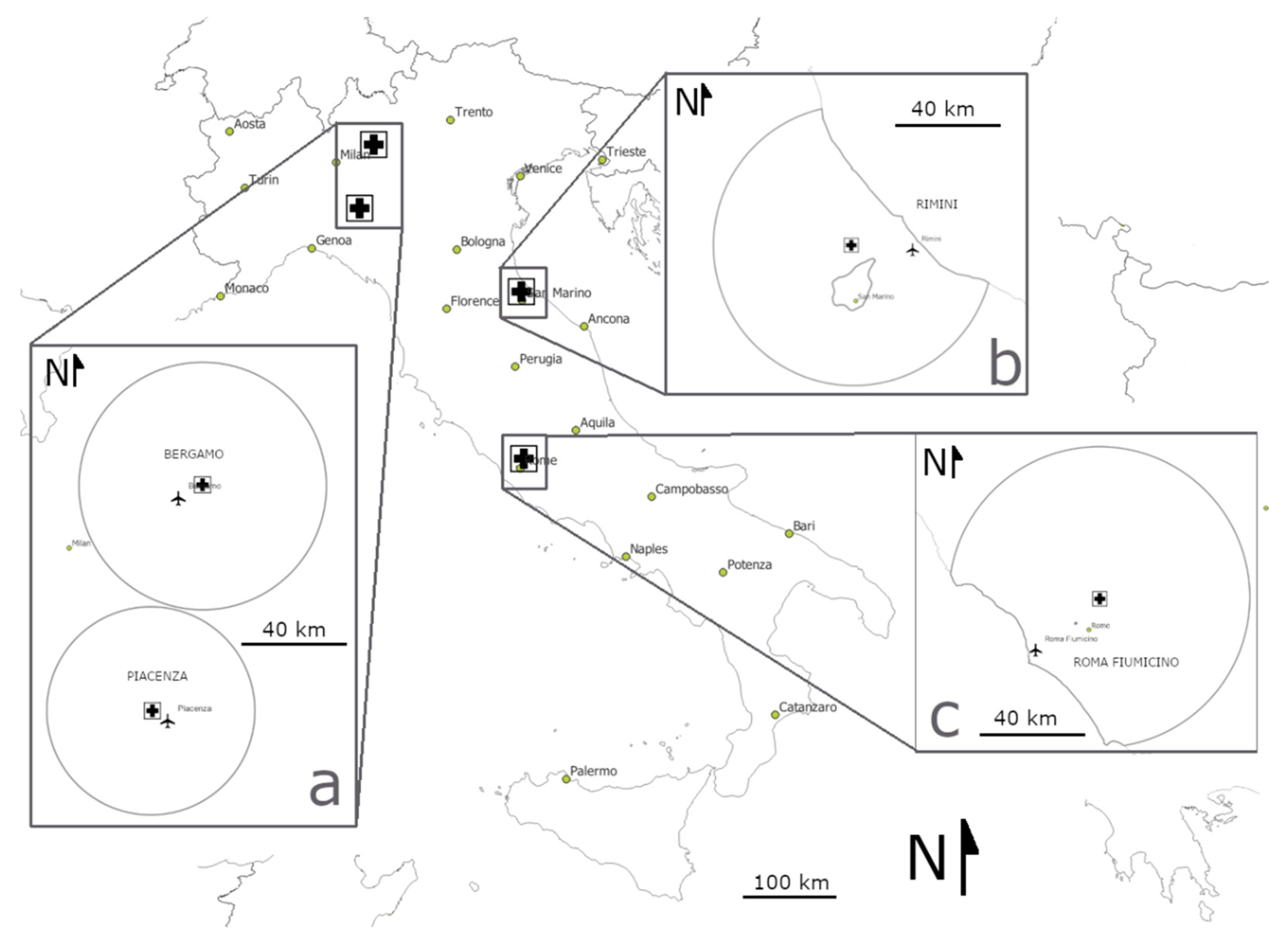{kind=link}
{kind=link}
| Total n Bats (n = 5842) | Young Bats (n = 4361) | Adults (n = 1481) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | d.f. | F | P | R2 (Adj) | F | P | R2 (Adj) | F | P | R2 (Adj) | |
| Year | 1 | 2.87 | 0.091 | 8.16 | 0.004 | 31.43 | 0.000 | ||||
| Centre | 3 | 45.54 | 0.000 | 30.99 | 0.000 | 45.27 | 0.000 | ||||
| Temperature | 1 | 9.34 | 0.002 | 6.09 | 0.014 | 10.11 | 0.002 | ||||
| Error | 736 | ||||||||||
| 16.53% | 12.19% | 19.43% | |||||||||
| Pipistrellus kuhlii | ||||||||||
| Total n Bats (n =900) | Young Bats (n =513) | Adults (n =387) | ||||||||
| Source | d.f. | F | P | R2 (Adj) | F | P | R2 (Adj) | F | P | R2 (Adj) |
| Year | 1 | 6.18 | 0.013 | 0.36 | 0.547 | 23.87 | 0.000 | |||
| Centre | 3 | 34.90 | 0.000 | 20.09 | 0.000 | 29.30 | 0.000 | |||
| Heatwaves | 1 | 7.87 | 0.005 | 5.50 | 0.019 | 4.51 | 0.034 | |||
| Error | 736 | |||||||||
| 14.12% | 7.94% | 14.29% | ||||||||
| Pipistrellus pipistrellus s.l. | ||||||||||
| Total n Bats (n = 60) | Young Bats (n = 34) | Adults (n = 26) | ||||||||
| Source | d.f. | F | P | R2 (Adj) | F | P | R2 (Adj) | F | P | R2 (Adj) |
| Year | 1 | 0.04 | 0.847 | 0.30 | 0.586 | 0.25 | 0.619 | |||
| Centre | 3 | 12.77 | 0.000 | 6.70 | 0.000 | 9.23 | 0.000 | |||
| Heatwaves | 1 | 0.68 | 0.411 | 0.02 | 0.877 | 2.02 | 0.155 | |||
| Error | 737 | |||||||||
| 4.31% | 2.01% | 3.16% | ||||||||
| Hypsugo savii | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total n Bats (n =1068) | Young Bats (n = 556) | Adults (n = 513) | ||||||||
| Source | d.f. | F | P | R2 (Adj) | F | P | R2 (Adj) | F | P | R2 (Adj) |
| Year | 1 | 6.34 | 0.012 | 1.16 | 0.283 | 9.18 | 0.003 | |||
| Centre | 3 | 35.27 | 0.000 | 15.90 | 0.000 | 30.60 | 0.000 | |||
| Heatwaves | 1 | 15.00 | 0.000 | 6.60 | 0.010 | 11.96 | 0.001 | |||
| Error | 737 | 15.38% | 7.20% | 13.45% | ||||||
| Tadarida teniotis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total n Bats (n = 2150) | Young Bats (n = 2066) | Adults (n = 84) | ||||||||
| Source | d.f. | F | P | R2 (Adj) | F | P | R2 (Adj) | F | P | R2 (Adj) |
| Year | 1 | 10.08 | 0.002 | 10.71 | 0.001 | 3.56 | 0.059 | |||
| Centre | 3 | 12.70 | 0.000 | 11.80 | 0.000 | 8.18 | 0.000 | |||
| Heatwaves | 1 | 2.84 | 0.092 | 3.06 | 0.081 | 1.65 | 0.199 | |||
| Error | 737 | 5.85% | 5.62% | 3.14% | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salinas-Ramos, V.B.; Tomassini, A.; Ferrari, F.; Boga, R.; Russo, D. Admittance to Wildlife Rehabilitation Centres Points to Adverse Effects of Climate Change on Insectivorous Bats. Biology 2023, 12, 543. https://doi.org/10.3390/biology12040543
Salinas-Ramos VB, Tomassini A, Ferrari F, Boga R, Russo D. Admittance to Wildlife Rehabilitation Centres Points to Adverse Effects of Climate Change on Insectivorous Bats. Biology. 2023; 12(4):543. https://doi.org/10.3390/biology12040543
Chicago/Turabian StyleSalinas-Ramos, Valeria B., Alessandra Tomassini, Fabiana Ferrari, Rita Boga, and Danilo Russo. 2023. "Admittance to Wildlife Rehabilitation Centres Points to Adverse Effects of Climate Change on Insectivorous Bats" Biology 12, no. 4: 543. https://doi.org/10.3390/biology12040543
APA StyleSalinas-Ramos, V. B., Tomassini, A., Ferrari, F., Boga, R., & Russo, D. (2023). Admittance to Wildlife Rehabilitation Centres Points to Adverse Effects of Climate Change on Insectivorous Bats. Biology, 12(4), 543. https://doi.org/10.3390/biology12040543