1. Animal Regeneration Derives from the Development of Specific Life Cycles
Once fully developed as an adult, an animal can incur small or large injuries, including the loss of body regions such as an arm, a leg, an eye, a kidney and so forth. The body senses the loss and repairs the damage through scarring in some animals while in others a regenerative process allows to anatomically and functionally restore the damaged organ. In the animal world regeneration abilities are very variable so that we can say that there are broad regenerators, medium–low regenerators and species that cannot regenerate [
1,
2,
3,
4,
5,
6,
7] (
Figure 1).
Why can some animals regenerate while others, in particular humans, cannot? This question should initially consider that a hydra or a planaria, the champions of regeneration, can broadly regenerate since they are simple multicellular organisms, although some species may have large genomes or genomes similar to those of more morphologically complex animals [
8]. In fact, genome size and number of functional genes are not correlated for numerous species and large genomes contain numerous repetitive DNA sequences with few active genes, especially for development. Therefore, genome coding for developing simpler anatomical planes have ample regeneration capacity while more complex bodies derived from large functional genomes are present in animals that cannot regenerate. The developmental, anatomical and physiological complexity of a vertebrate is much higher from those of a hydra, a flatworm, a ribbonworm, a polychete anellid or a seasquirt and justify the poor regeneration of the former versus the high regeneration ability of the latter.
Despite the above consideration, it is very important to learn the mechanisms of regeneration from regenerative species in the hope that this knowledge will allow translating these mechanisms into non-regenerative animals. This question has activated a vast amount of research in the last 150 years and, lately, on cellular, molecular biology and on the genetics of regeneration [
2,
3,
9,
10]. The final goal of these investigations is the discovery of molecular clues from regenerating animals and to apply this knowledge to improve human regeneration [
11,
12]. Although the basic molecular mechanisms will ultimately explain the ample regeneration of competent organisms, most physicians and biologists neglect to see general basic biological reasons for the presence of regeneration only in some animals and its absence in others [
1,
5]. The noted differences among species reflect how complex development occurs in different environments and the anatomical organization present in diverse animals as adults. The latter is an adaptation to the environment during the progressive phases of the biological cycle in each species—cycles that derive from a long evolutionary history (
Figure 2).
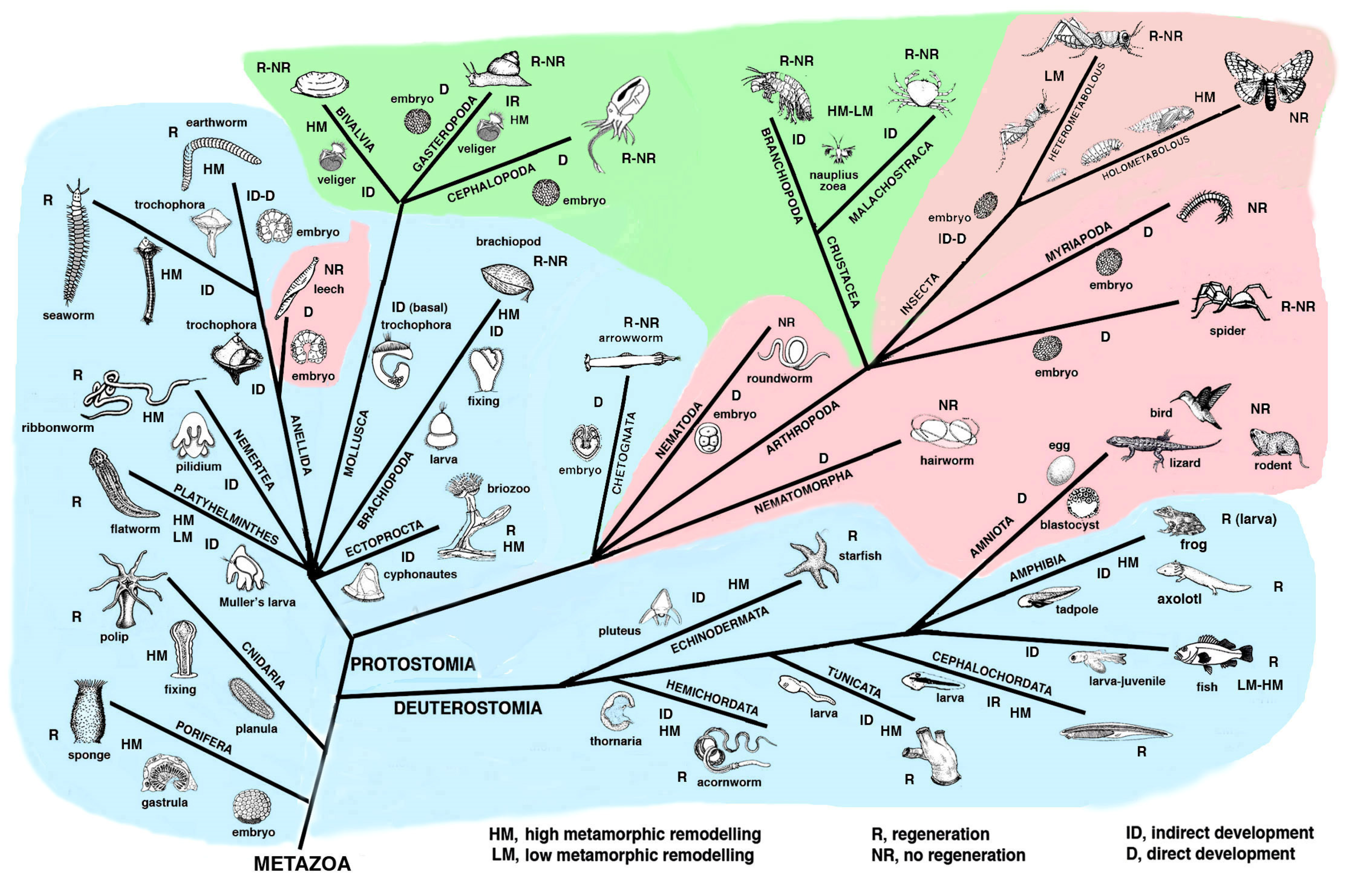
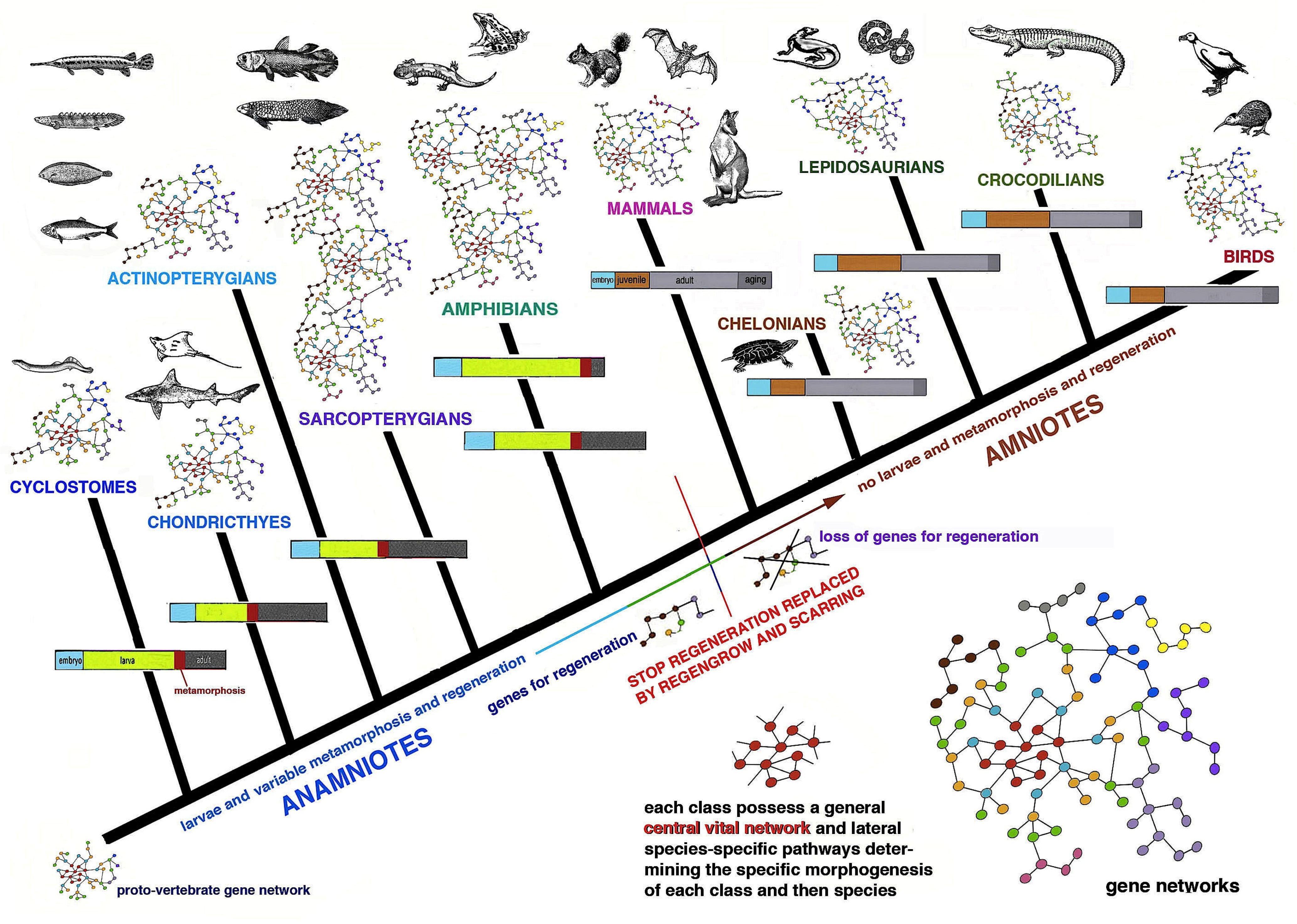
Figure 1 and
Figure 2 summarize the distribution of regeneration among metazoans, noting that most regenerative species are linked to the water environment and are mainly marine animals. Numerous low forms of marine animals such as sponges, flatworms, ribbonworms and ctenophores have less complex bodies in comparison to those of terrestrial animals, particularly those of arthropods and vertebrates. However, a relatively high regenerative ability is also present in more complex animals, such as sea slugs, sea cucumbers, starfish, seasquirt, numerous fishes, caudate amphibians and frog tadpoles—all animals linked to the sea, freshwater or humid environments and that have metamorphosis in their life cycle (
Figure 2). Other animals such as earthworms, octopus, sea snails, numerous crustaceans and many fishes, also living in water or humid environments but missing drastic meta-morphic transitions, can repair or even regenerate few organs. Another group of terrestrial animals such as spiders, some crustaceans and insects with progressive growth through molts, can only regenerate their appendages but little or none of their inner organs (nervous ganglia, heart, intestine, hepatopancreas, gonads etc.). Finally, other terrestrial animals such as roundworms and scorpions, and all terrestrial vertebrates (amniotes) such as reptiles, birds and mammals, cannot regenerate at all, aside from healing their wounds by scarring and occasionally through “
regengrow” [
1,
4,
5,
13,
14,
15,
16,
17,
18]. The term
regengrow indicates a healing or even regenerative process that occurs while the animal is still variably growing, and this concept has been broadly explained in previous studies [
18,
19]. Broad regeneration only occurs in very hydrated conditions and this process is much more common in sea- or freshwater-adapted animals than in terrestrial adapted animals (
Figure 1 and
Figure 2). This depends on the formation of a regenerative blastema, a soft and hydrated environment for cell multiplication, movements and for the special re-organization that would be hampered in denser and drier tissues typically found in post-embryonic terrestrial animals.
The present analysis is hypothetical and is based on general biological considerations derived from the specific evolutionary trajectories taken by different phyla (
Figure 1), but the molecular developmental details of the specific evolution in each group of animals represent a huge effort for next generations of biologists.
2. Distribution of Regeneration among Animals
Lower invertebrates include acelomates (Poriphera, Cnidaria, Ctenophora, Platyhelminthes, Nemerteans), pseudocelomates (Nematods, Nematomorphs, Rotifers, Achanthocephals, Gastrotrics, Ectoprocts, and other minor groups) and the remaining celomates (mainly Anellids, Molluscs, Arthropoda, Echinoderms and Chordates;
Figure 1). Pseudocelomates and celomates have reached an organ level organization based on a general increase of their genomes and gene regulation [
8]. The biological cycles are variable, including those of anellids, molluscs, arthrophods, echinoderms and chordates (
Figure 2). During evolution, most of these phyla of animals initiated their biological cycles with a primitive, indirect development with a trochophora or other natant types of larvae, but some evolutionary lineages later shortened or suppressed these larval stages that were replaced by a direct development for the terrestrial adaptation [
20].
Using simple models of gene networks in the following discussion I only mean to provide a visual and general indication on the thousands of genes present in different animal genomes [
8] capable or not to determine a regenerative process (
Figure 3).
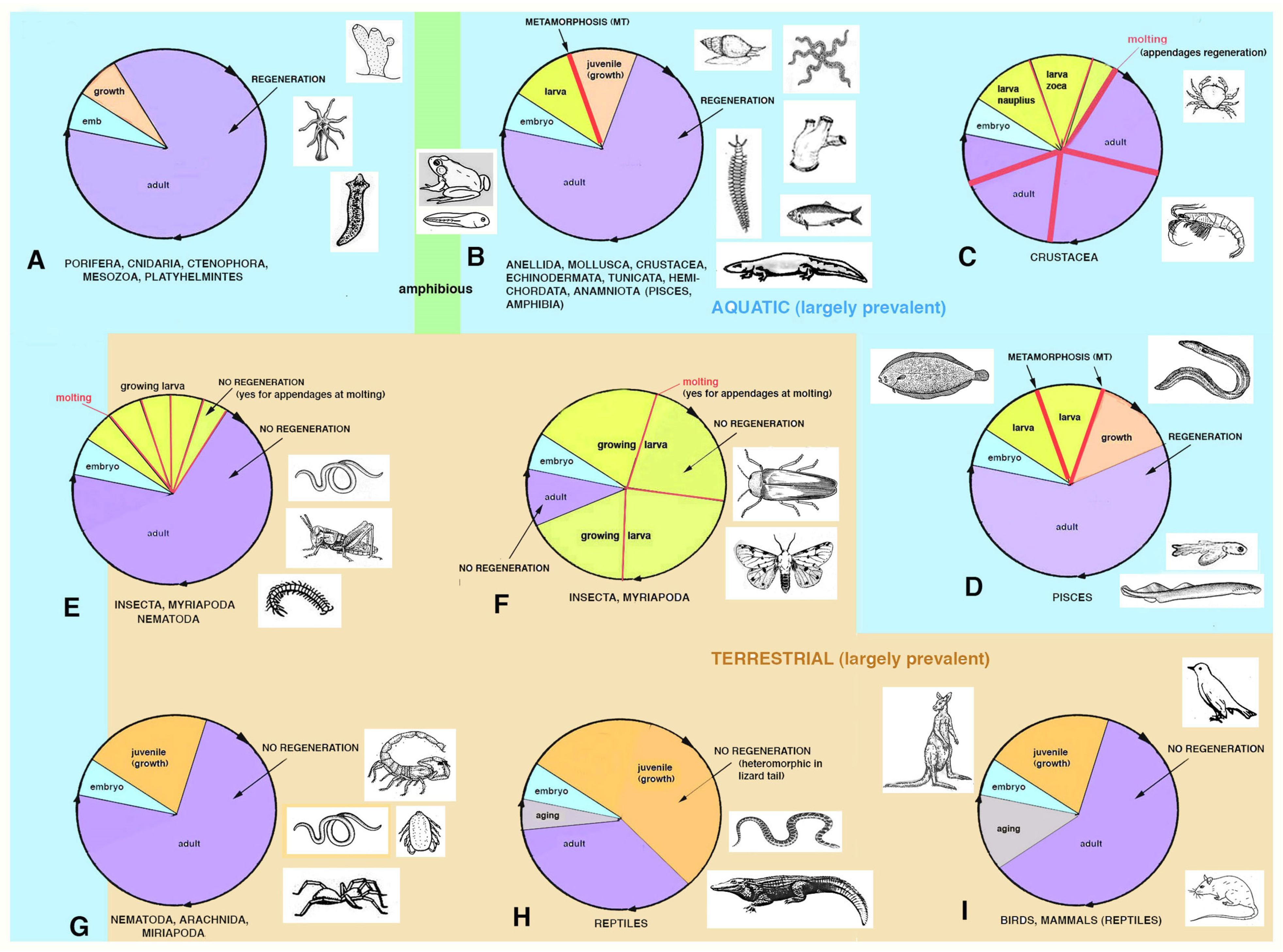
In general, the development and anatomical simplicity of the resulting adult phase favors regeneration because of the lower number of “gene regulative networks” necessary to develop simple organisms that have a tissue organization level of complexity in their bodies such as sponges, jellyfish, planarians, ribbonworms. Also in more complex organisms such as seaworms (anellids) or echinoderms or marine chordates, that are master of regeneration, their more complex anatomical organization can be regenerated after injury as they possess ample restructuring processes operating during their metamorphosis, and that can be largely re-used in case of injury [
2,
4]. The variable regenerative ability present in these animals, from large body regions to minute organs, suggests that these animals re-utilize a number of metamorphic-developmental genes for their partial or broad regeneration when they become adults but are still living in an aquatic environment (
Figure 3). Only metamorphic processes that involve large re-modeling of the larval body to reach the sexually mature adult condition determine a high or discrete ability to regenerate as adults. This derives from genomes involved in large restructuring morphogenetic processes that transform a larva into an adult [
15,
16,
17]. In contrast, the simple increase of body size during larval growth after successive molts, such as in numerous crustaceans, insects or some spiders, is not associated with a broad regeneration in adults, aside from body appendages [
1,
2,
3,
4,
5,
6]. This likely derives from the lack of developmental gene networks implicated in broad morphogenetic and restructuring processes in the genomes of these animals operating during their life-cycle.
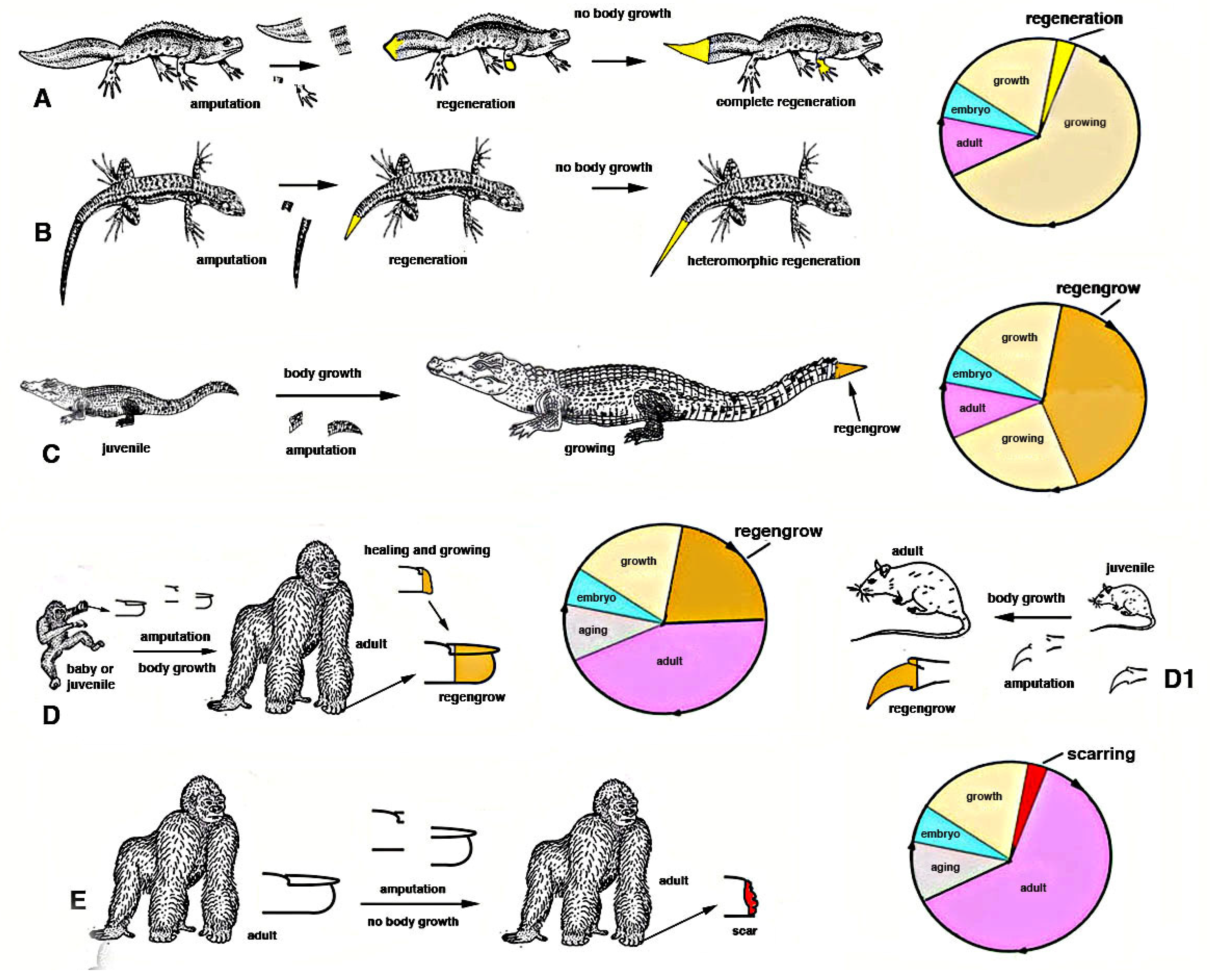
After injury in some species of juvenile arthropods and vertebrate amniotes, wound healing takes place together with their ongoing somatic growth and the injured or lost organ can apparently regenerate. This occurs for some appendages such as legs, digits or tails [
1,
11]. However, what derives from true tissue regeneration or from the process of somatic growth of the same tissue is undefined and this mixed healing process is indicated as
regengrow [
18,
19,
21] (
Figure 4).
In
growing invertebrates or vertebrates various tissues increase their mass due to a more or less continuous production of new cells, from local staminal/reserve cells. For instance a growing squid, a spider or an insect, a fish or a snake, a rat or a child possess muscles that add new (satellite) cells inside the myofiber, contributing to their volumetric increment before reaching the specific full growth of the species. Other processes of addition of new cells also occurs in the new cuticle that is formed from imaginal disks in insects as long as molt occurs, or in growth plates of vertebrate long bones, or in the liver and intestine during somatic growth etc. In case of injury or traumatic loss of muscles, skin, liver etc. in still growing animals, the activation of a wound healing process simply increases the rate of cell proliferation and tissue growth that was already present in these animals. Consequently, what derives from the physiological growth process superimposed to the stimulated healing or broader regeneration becomes undefined, and represents a mixed process indicated as
regengrow. Regengrow is progressive but requires time to occur, over 4-5% of its life-span [
18,
19,
21], like the growth of that specific animal, and stops when these animals complete their somatic growth. Numerous examples of
regengrow are reported for vertebrates [
18,
19], but also numerous terrestrial invertebrates incurs in a similar process. At the end of growth in each species, and during aging in species that manifest this process, the addition of new cells is reduced, halted and only a physiological regeneration (skin, blood, intestine etc.) remains. An injury at this late stage of the life cycle in different species (e.g., after the end of the last molt in a crab or in a grasshopper or after reaching adulthood in a mammal etc.) cannot anymore combine growth (ceased) with wound healing/regeneration, and only scarring can occur with the permanent loss of the injured organ.
In contrast, a true process of regeneration takes a relatively short time (2–3% of the lifetime of a species, at least in vertebrates) such that the process of somatic growth is not influencing or overlapping with the healing (regenerative) process [
21]. While
regengrow takes a long time, even years in some vertebrates to apparently reform the missing organ, regeneration rapidly occurs through proliferation and differentiation of a very large number of new cells that determines the reconstitution of the injured organ. Regeneration can also occur during active somatic growth and also in invertebrate and vertebrate larvae, but requires a short time where the growth contribution is minimum or irrelevant. Regeneration also rapidly occurs in any other phases of the life cycle of regenerative competent species, including adult and even during aging, while
regengrow is no longer possible at the end of growth in adults [
19]. For example, when full somatic growth is reached in an animal, the amputation of a mammalian digit, a crocodilian tail, the leg of a crab or a lobster or a cockroach or a grasshopper or a spider after the last molt, determines scarring or the formation of short outgrowths but not their regeneration (
Figure 4).
Chordate regeneration is very likely a basal capability for the phylum like asexual propagation (reproduction), and the fact that extant chordate larvae can broadly regenerate after injury supports this notion [
22] (
Figure 1). The only vertebrates capable of a variable but generally broad inner organ and appendages regeneration are water-related, fish and amphibians, while amniotes can only heal their wounds or form scars [
3,
13,
15,
17]. The only exception among amniotes is the broad regeneration of the tail in numerous lizards, often a short process that takes 2–4% of the entire life-span of a lizard to restore a voluminous organ (1/7th–1/4th of the body mass), but that gives rise to a heteromorphic tail, missing of axial anatomical structures. Therefore, a general conclusion is that after organ loss the more larval-metamorphosis genes recruited in adults of invertebrates or vertebrates, the better the regeneration that takes places (
Figure 3).
The above discussion indicates that gene networks present in any genome can be largely or partially re-activated for regeneration in simple or more complex aquatic animals with larval stages and metamorphosis while terrestrial animals, without metamorphosis, cannot re-activate functional gene networks for regenerate their lost organs. The presence of metamorphic events in the life cycle of animals, determined by genes, for organ destruction of larvae and for re-construction of new organs in adults, allows for the re-use of these genes in part for regeneration in adult animals living in water but not on the land [
16] (
Figure 3). After injury, it is difficult to activate alternative developmental pathways from those utilized during development, especially in complex terrestrial arthropods or vertebrates. Additionally, during leg regeneration in arthropods or tail regeneration in lizards, a process that allows a broad but heteromorphic regeneration, only few developmental genes appear to be re-activated [
15,
16,
17,
18,
23,
24,
25]. This suggests that regeneration, like development, requires the function of basic developmental pathways, which have altered during the evolution of different forms of metazoans, resulting in an inability to regenerate [
5].
3. Regenerative Species Evolved Genomes including Programs for Larvae Development and Metamorphosis
The genomes of different animals have been acquired during the evolution in each lineage of metazoans and allow their specific phases of development and post-embryonic development during the progression in their specific life cycles (
Figure 2 and
Figure 5). We should interrogate ourselves and reflect on the biological reasons that motivate regeneration only in some species. Can what we learn from regenerative species really be applied to non-regenerating species? A response should not only look to the genetic and informational molecules but also elaborate on the specific evolutive process (trajectories) that generated different animal species capable or incapable of regeneration (
Figure 1 and
Figure 5).
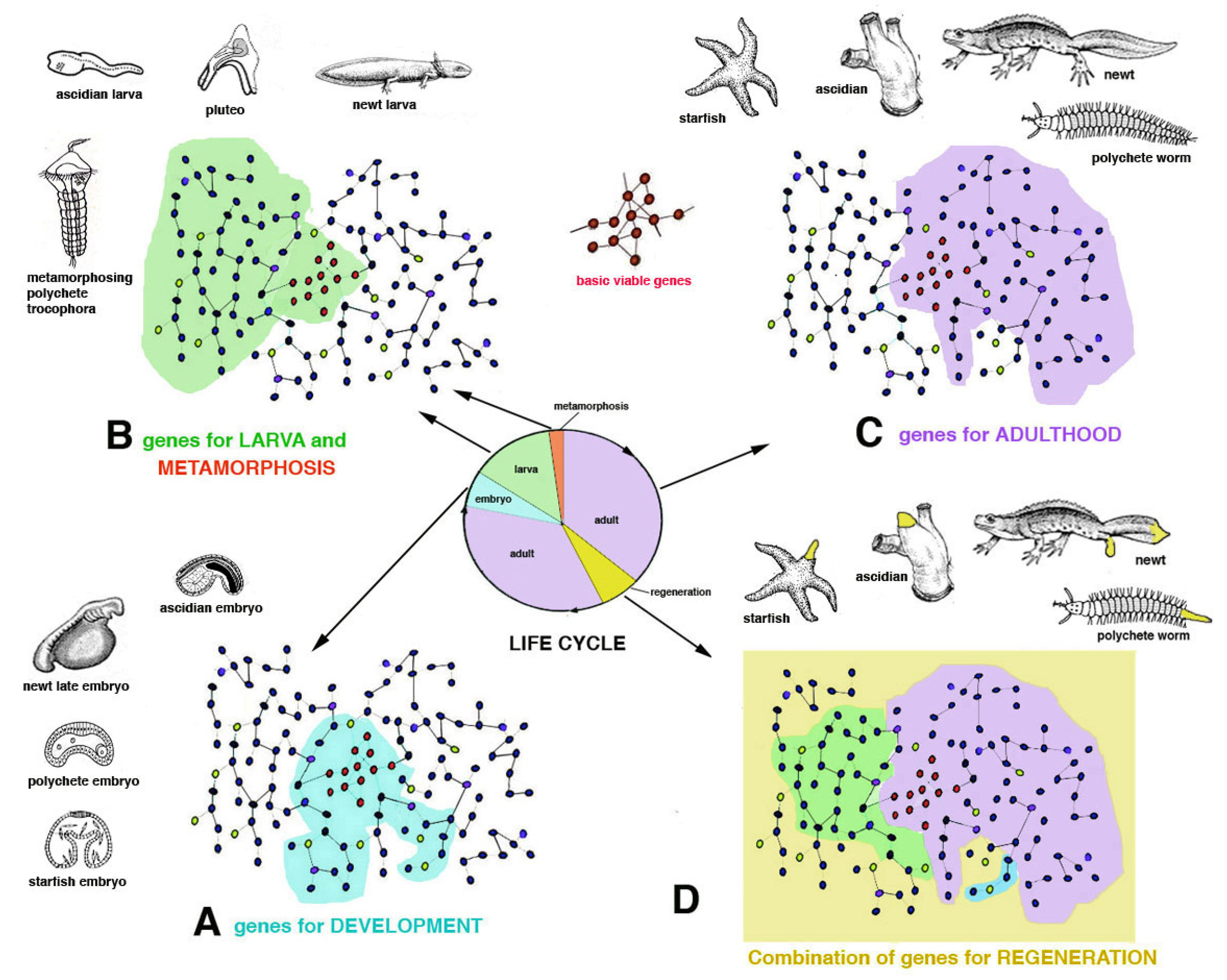
The evolution in each lineage gave rise to species with different genomes but that possess a common region for basic metabolic and genetic functions (basic vital genes, ideally shown in
Figure 3C). These are roughly common among most organisms for metabolism, cell division, cell communication and interactions. Other genes work in different networks (schematically indicated as peripheral areas in
Figure 3 and
Figure 6) for specific functions (development, endocrine, nervous, sensing, movement, reproduction, waste elimination, etc.). Among specific functions I include healing from injuries through a broad or limited regeneration or through scarring.
In the case of vertebrates, broad organ regeneration was lost during the transition from water to land in the Upper Carboniferous, a transition that determined the partial or total loss of the larval stage and of intense metamorphic transformation [
14,
15,
16,
17,
26]. I hypothesize that some or most of the gene networks that are utilized for larva and metamorphic transformations also functioned for organ regeneration in adults of fish and amphibians, but they were lost or modified for the terrestrial adaptation in amniotes [
16] (
Figure 5). Even the insertion of only one single, new gene present in amniotes into the genome of a fish can inhibit its ability to regenerate the fin [
27], demonstrating that the developmental regenerative program can be easily altered.
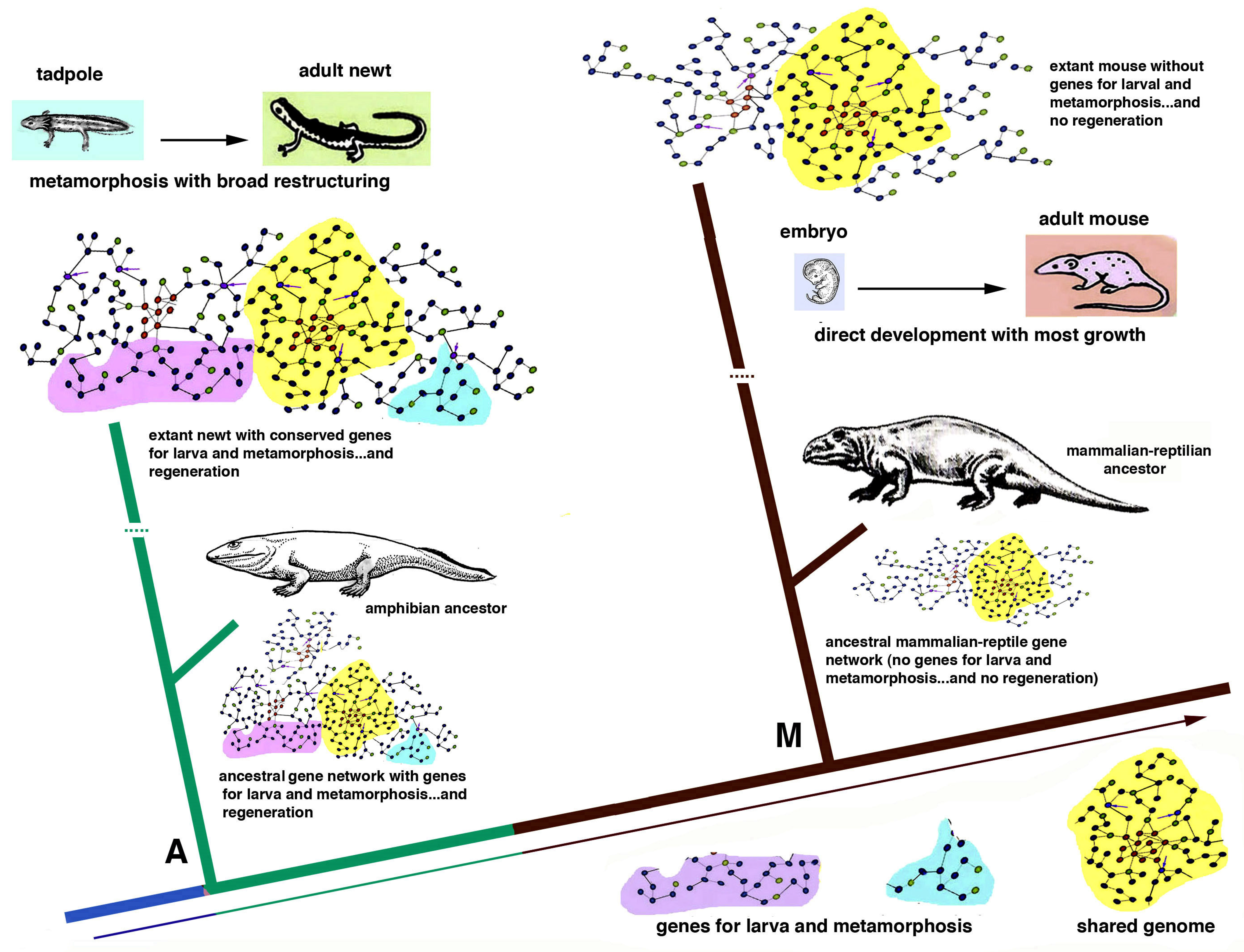
As a theoretical example of gene insertion in different species, we here consider two very different vertebrates, a newt and a mouse, that derived from a common ancestor, about 280–300 million years ago [
20] (
Figure 6). Like the amphibian ancestor of the Carboniferous Period, the extant newt possesses a genome that includes several gene networks in common with those of a mammalian-reptilian ancestor and in the extant mouse (idealized yellow areas in
Figure 6). This “central region of the genome” governs mainly basic viable processes needed for both these species. Other regions of the genome of the newt, but not in the mouse, represented in
Figure 6 as located peripherally to the gene network (in pink and light blue colors in
Figure 6), act on specific anatomical-physiological characteristics of the newt such as development of a larval stage and metamorphosis. The way morphogenesis occurs in the newt and in the mouse is, however, different in many other gene networks that evolved in amphibians in comparison to others that evolved in mammals. Like its amphibian ancestors, the newt possesses a larval stage with broad metamorphosis that determines a large body-restructuring and a derived broad ability to regenerate appendages and inner organs (
Figure 3 and
Figure 5). In contrast, the mouse, like its reptilian-mammalian ancestor, has a direct development and the delivered baby mainly grows into a juvenile mouse with no metamorphosis. In case of general injury, an adult mouse can only rapidly heal by scarring [
13] or producing new digit tips by
regengrow if the loss occurred in juveniles (
Figure 4D1). The different evolution of the “gene networks” undergone in the two species cannot be reversed by the transplant of genes only involved in regeneration (
Figure 6 and
Figure 7, see later discussion).
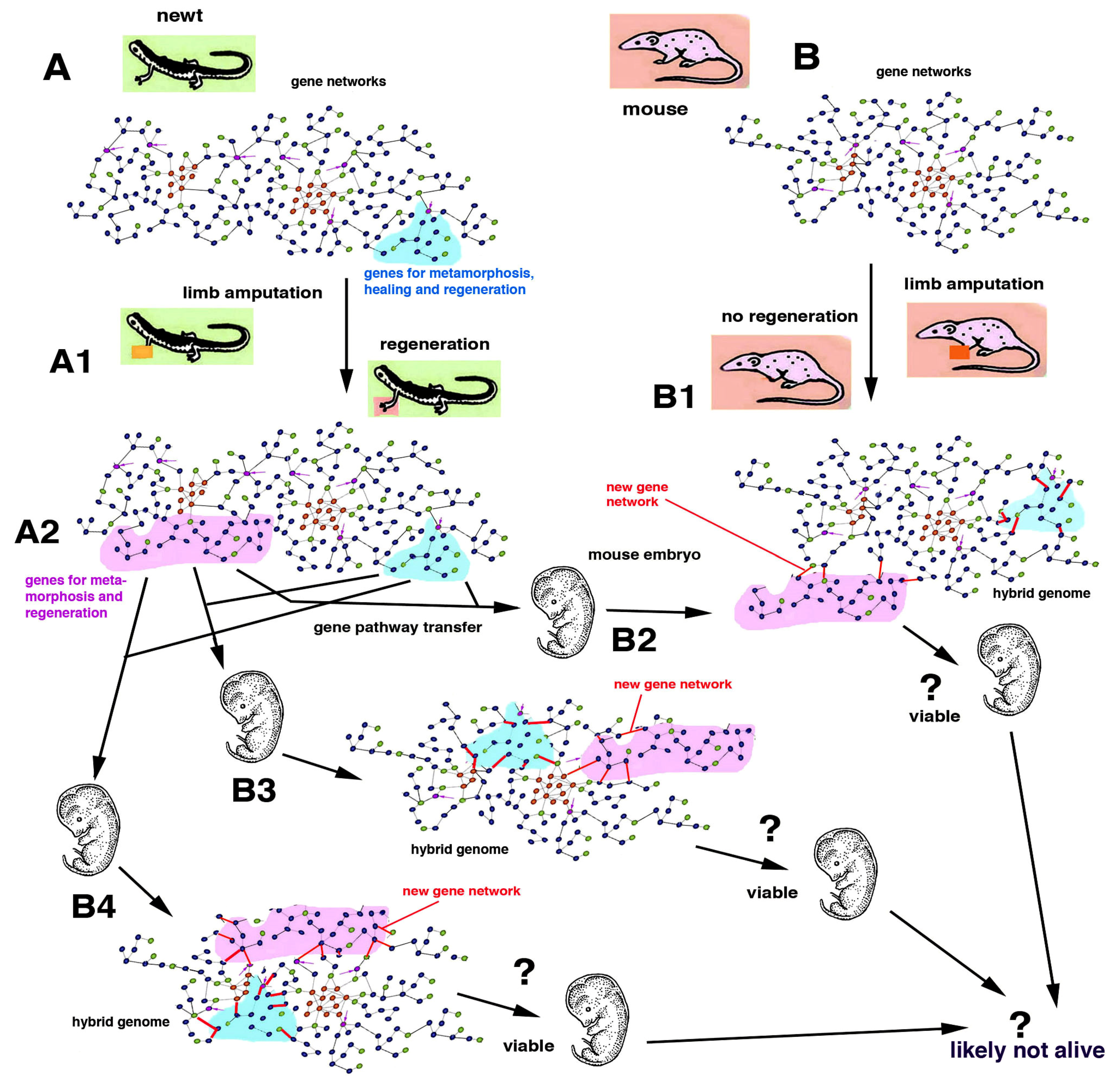
General gene pathways governing regenerative cellular processes in polyps, flatworms, earthworms, starfish and chordates including urodele amphibians will hopefully be deciphered in the future with the goal of applying this knowledge to animals with no regeneration [
7,
12,
24]. This future knowledge will have to face the biological specificity of the developmental processes of non-regenerating animals and the biological limits and perils derived from the introduction of “regenerative gene pathways” within the genomes of embryos or adults in species that have evolved abolishing the gene regulatory mesh-works of regeneration initially present in their ancestors [
8] (
Figure 5 and
Figure 6). The above consideration indicates that the information on the molecular mechanisms of regeneration gained from simple models (hydra and planaria) but also from urodele amphibians and fish, cannot be directly applied to different metazoans, especially to vertebrates, because their evolution followed so many different developmental trajectories that they cannot be modified anymore without introducing unpredictable effects (?) including cancer, teratomas or even monstrosity or, more likely, death of the embryo (
Figure 7).
4. Problems to Induce Regeneration in Non-Regenerating Animals
Gene therapy functions with genes operating by the terminal phase of a gene pathway, e.g., the origin of an eye pigment (cure for albinism) or of hemoglobin production (cure for thalassemia or sickle cell anemia). In contrast, in the case of developmental pathways and their interactions with other pathways within a gene network, it is difficult to predict the outcome of a gene therapy. To further complicate the entire matter of the genetic material and its manipulations, recent studies have indicated the instability of genomes with the loss of segments of DNA in somatic cells [
28,
29]. Whether the loss of DNA during evolution may also be connected to the loss of regeneration in some phyla, classes or families of metazoans remains an open question for future investigations. Whatever the case, it is likely that with present and future technologies, the molecular details of the developmental pathways leading to various levels of organ regeneration in urodeles will become known. At that point, some gene therapies might be planned, transferring some key genes or one or more “gene modules” identified in the newt (or axolotl) into (a) the genome of a fertilized egg or the early embryos of a mouse, or (b) into some mouse cells that are implanted into a damaged but not completely lost organ [
30,
31,
32]. In the first case (a), this “early regenerative gene therapy” for curing an organ or limb loss that may occur in adults of mice would be unpredictable since the numerous interactions and consequences on the entire function of a mouse genome would be likely teratogenic and lethal (
Figure 7). In fact, gene pathways that have, since at least 280–300 million years ago, evolved to form numerous differences between those of a newt and a mouse cannot be re-assembled anymore. Few experiments have already indicated that to stop regeneration, only the alteration of some (key) genes that lead to change of basic signaling pathways such as Wnt and BMP or other genes is required [
2,
6,
32]. This type of genetic manipulation would very likely produce teratomas and death of the embryo and is consequently not appliable (
Figure 7).
In the second case (b), some attempts of gene therapies for tissue regeneration are currently in trial but these therapies insert few specific genes directly into specific injured organs or insert specific genes in cell types that are later transplanted in precise locations of the damaged organs [
30,
32,
33]. The engineered cells proliferate and give rise to a regenerated tissue but do not integrate into the genome of the receiving animal. However, numerous problems must be solved, such as the type of viral or non-viral (e.g., liposomes) vectors and care should be exerted to control that the regenerating tissue does not turn into a tumor. These methods are an extension of those that use stem cells for stimulating tissue regeneration [
31,
32]. New technologies that utilize innovative genetic manipulations such as “tissue regeneration enhancer elements” and are delivered to localized cells of specific organs are currently under active study [
33].
Some improvements in healing can be obtained in non-regenerating animals, including humans, using these methods when the technology will be perfected. Regenerative gene therapies transferring genes such as
Wnt,
BMP,
VEGF,
PDGF,
IGF,
TGGb1 etc. are in trial for bone, teeth and cartilage regeneration, as are
microdystrophin and
Cyclin A2 genes for muscle regeneration,
CDGF,
BDGF,
GDNF genes for neural regeneration,
KGF,
IGF1 genes for skin regeneration and so forth [
32]. For inducing digit and limb regeneration in mice, other genes transfer (
Lin28) has been attempted [
31]. Whether this gene therapy will last in the injected cells in order to be able to induce the regeneration of large organs such as limbs, heart, intestine, kidneys, etc. remains uncertain. This results from the lack of information on the fate of the engineered cells among adult tissues and under immunological check-up from the receiving body. Furthermore, the formation of a possible blastema should keep the tissue wet since regeneration only occurs in a liquid environment, as previously indicated [
21]. Other technological avenues (c) should be taken for alleviating inner organ, leg and arm loss based on tissue engineering or combining bio-engineered prostheses [
31,
34] and not acting on invariant animal genomes derived from long evolutive differences (
Figure 7). The functionality of organs after injury or pathological damage could be replaced efficiently only through improvements in the technology of prostheses and organ implants which are not impinging on genetic problems but mainly have immunological problems related to the acceptance of these organs.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






