


Red Ginseng Attenuates the Hepatic Cellular Senescence in Aged Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Animals
2.3. Glucose and Insulin Tolerance Test
2.4. Immunoblotting Assay
2.5. Isolation of Primary Hepatocytes and Mouse Embryonic Fibroblasts (MEFs)
2.6. Cellular Senescence Induction in MEFs by Ionizing Radiation (IR)
2.7. Senescence-Associated β-Galactosidase (SA-β-gal) Staining
2.8. Statistical Analyses
3. Results
3.1. Red Ginseng Supplementation Is Involved in Glucose Homeostasis in Aged Mice
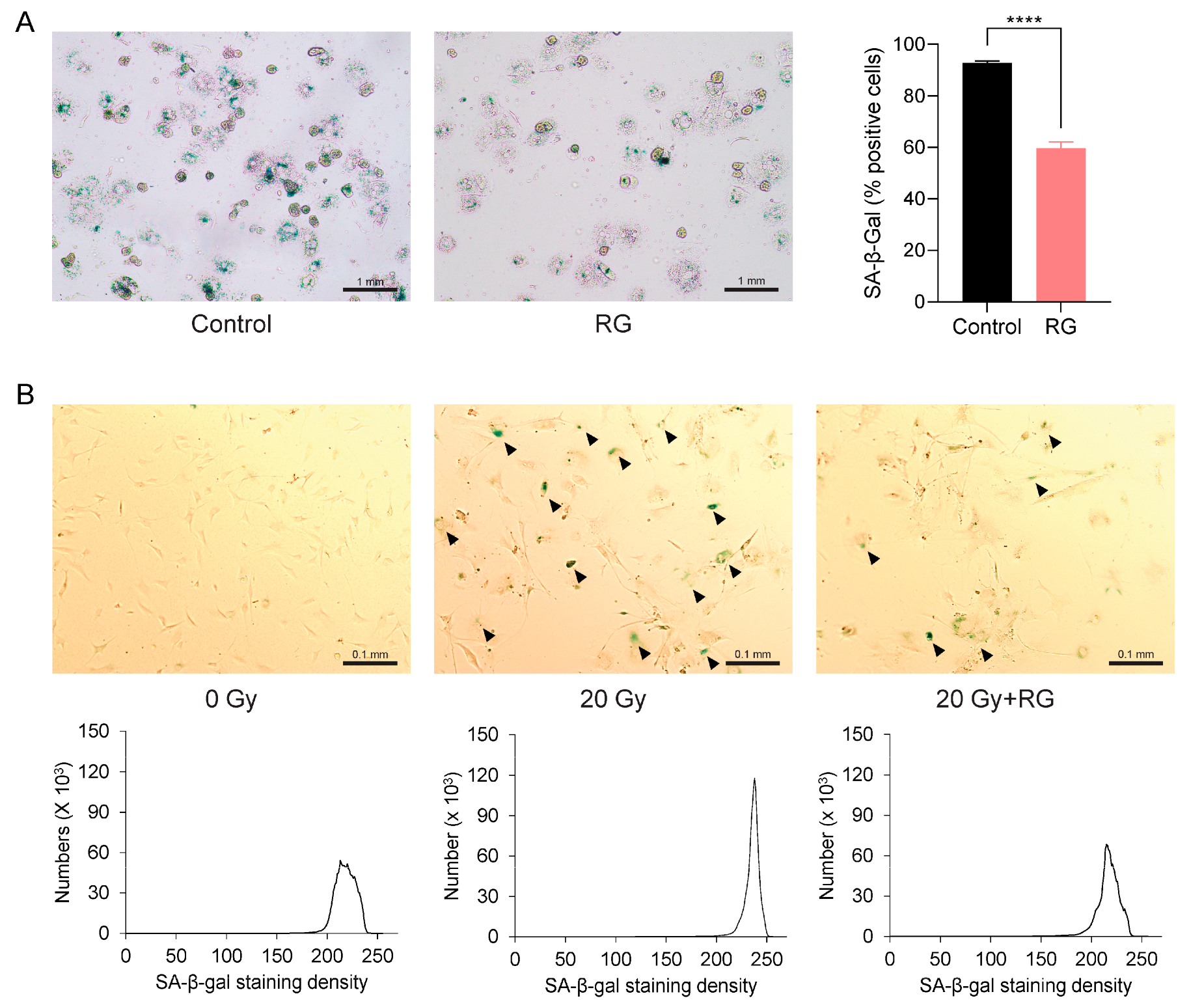
3.2. Red Ginseng Attenuates Hepatic Cellular Senescence Pathways in Aged Mice
3.3. Red Ginseng Administration Improves Insulin Homeostasis in Aged Mice
3.4. Red Ginseng Suppresses the JNK/NF-κB Signaling Pathway in Aged Mice
3.5. Red Ginseng Alleviates Cellular Senescence in Primary Hepatocytes and MEFs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di Micco, R.; Krizhanovsky, V.; Baker, D.; d’Adda di Fagagna, F. Cellular senescence in ageing: From mechanisms to therapeutic opportunities. Nat. Rev. Mol. Cell Biol. 2021, 22, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.G.; Durik, M.; Baker, D.J.; Van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Jat, P. Mechanisms of cellular senescence: Cell cycle arrest and senescence associated secretory phenotype. Front. Cell Dev. Biol. 2021, 9, 485. [Google Scholar] [CrossRef] [PubMed]
- Mijit, M.; Caracciolo, V.; Melillo, A.; Amicarelli, F.; Giordano, A. Role of p53 in the Regulation of Cellular Senescence. Biomolecules 2020, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-J.; Jiang, S.-S.; Zhang, J.; Luo, D.; Yu, B.; Yang, L.-Y.; Zhong, H.-H.; Yang, M.-W.; Liu, L.-Y.; Hong, F.-F.; et al. Effects of apoptosis on liver aging. World J. Clin. Cases 2019, 7, 691–704. [Google Scholar] [CrossRef]
- Amaya-Montoya, M.; Pérez-Londoño, A.; Guatibonza-García, V.; Vargas-Villanueva, A.; Mendivil, C.O. Cellular senescence as a therapeutic target for age-related diseases: A review. Adv. Ther. 2020, 37, 1407–1424. [Google Scholar] [CrossRef] [PubMed]
- Lagoumtzi, S.M.; Chondrogianni, N. Senolytics and senomorphics: Natural and synthetic therapeutics in the treatment of aging and chronic diseases. Free Radic. Biol. Med. 2021, 171, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Qin, L.; Feng, R.; Hu, G.; Sun, H.; He, Y.; Zhang, R. Emerging senolytic agents derived from natural products. Mech. Ageing Dev. 2019, 181, 1–6. [Google Scholar] [CrossRef]
- Li, X.-G.; Yan, Y.Z.; Jin, X.-J.; Kim, Y.K.; Uddin, M.R.; Kim, Y.B.; Bae, H.; Kim, Y.C.; Lee, S.W.; Park, S.U. Ginsenoside content in the leaves and roots of Panax ginseng at different ages. Life Sci. J. 2012, 9, 670–683. [Google Scholar]
- Lee, M.R.; Yun, B.S.; In, O.H.; Sung, C.K. Comparative study of Korean white, red, and black ginseng extract on cholinesterase inhibitory activity and cholinergic function. J. Ginseng Res. 2011, 35, 421. [Google Scholar] [CrossRef]
- Lee, S.M.; Bae, B.-S.; Park, H.-W.; Ahn, N.-G.; Cho, B.-G.; Cho, Y.-L.; Kwak, Y.-S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, T.; Kim, S.-W.; Hwang, S.-Y.; Sohn, S.-H.; Yoo, S.-K.; Kim, S.-K. Panax ginseng reduces oxidative stress and restores antioxidant capacity in aged rats. Nutr. Res. 2012, 32, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.S.; Yoon, M. Korean red ginseng (Panax ginseng) inhibits obesity and improves lipid metabolism in high fat diet-fed castrated mice. J. Ethnopharmacol. 2018, 210, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-H.; Kim, K.-J.; Chei, S.; Seo, Y.-J.; Lee, K.; Lee, B.-Y. Korean red ginseng and Korean black ginseng extracts, JP5 and BG1, prevent hepatic oxidative stress and inflammation induced by environmental heat stress. J. Ginseng Res. 2020, 44, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Koo, S.; Ha, K.-T.; Kim, S. Neuroprotective effect of Korea Red Ginseng extract on 1-methyl-4-phenylpyridinium-induced apoptosis in PC12 Cells. Anim. Cells Syst. 2016, 20, 363–368. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, H.; Ko, E.-J.; Lee, Y.-N.; Kwon, Y.-M.; Kim, M.-C.; Kang, S.-M. Immunomodulatory activity of red ginseng against influenza A virus infection. Nutrients 2014, 6, 517–529. [Google Scholar] [CrossRef]
- Kim, E.-H.; Kim, S.-W.; Park, S.-J.; Kim, S.; Yu, K.-M.; Kim, S.G.; Lee, S.H.; Seo, Y.-K.; Cho, N.-H.; Kang, K.; et al. Greater efficacy of black ginseng (CJ EnerG) over red ginseng against lethal influenza A virus infection. Nutrients 2019, 11, 1879. [Google Scholar] [CrossRef]
- Kim, H.J.; Oh, T.K.; Kim, Y.H.; Lee, J.; Moon, J.M.; Park, Y.S.; Sung, C.M. Pharmacokinetics of ginsenoside Rb1, Rg3, Rk1, Rg5, F2, and compound K from red ginseng extract in healthy Korean volunteers. Evid. Based Complement. Altern. Med. 2022, 2022, 8427519. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Goldblatt, P.J.; Hinton, D.E.; Lipsky, M.M.; Chacko, J.; Trump, B.F. Mouse liver cell culture. In Vitro 1981, 17, 913–925. [Google Scholar] [CrossRef]
- Park, J.Y.; Lee, D.-S.; Kim, C.-E.; Shin, M.-S.; Seo, C.-S.; Shin, H.-K.; Hwang, G.S.; An, J.M.; Kim, S.-N.; Kang, K.S. Effects of fermented black ginseng on wound healing mediated by angiogenesis through the mitogen-activated protein kinase pathway in human umbilical vein endothelial cells. J. Ginseng Res. 2018, 42, 524–531. [Google Scholar] [CrossRef]
- Rufini, A.; Tucci, P.; Celardo, I.; Melino, G. Senescence and aging: The critical roles of p53. Oncogene 2013, 32, 5129–5143. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Sharpless, N.E. The regulation of INK4/ARF in cancer and aging. Cell 2006, 127, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.V.; Vaughn, E.; Sunohara-Neilson, J.; Ovari, J.; Leri, F. Oral gavage in rats: Animal welfare evaluation. J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 25–30. [Google Scholar] [PubMed]
- Anerillas, C.; Abdelmohsen, K.; Gorospe, M. Regulation of senescence traits by MAPKs. GeroScience 2020, 42, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Huuskonen, J.; Ojala, J.; Kauppinen, A.; Kaarniranta, K.; Suuronen, T. Activation of innate immunity system during aging: NF-kB signaling is the molecular culprit of inflamm-aging. Ageing Res. Rev. 2008, 7, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Manning, J.A.; Kumar, S. A potential role for NEDD1 and the centrosome in senescence of mouse embryonic fibroblasts. Cell Death Dis. 2020, 1, e35. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Kim, T.-H.; Kim, J.-Y.; Bae, J.; Kim, Y.-M.; Won, M.-H.; Ha, K.-S.; Kwon, Y.-G.; Kim, Y.-M. Korean Red ginseng prevents endothelial senescence by downregulating the HO-1/NF-κB/miRNA-155-5p/eNOS pathway. J. Ginseng Res. 2021, 45, 344–353. [Google Scholar] [CrossRef]
- Cho, D.-E.; Choi, G.-M.; Lee, Y.-S.; Hong, J.-P.; Yeom, M.; Lee, B.; Hahm, D.-H. Long-term administration of red ginseng non-saponin fraction rescues the loss of skeletal muscle mass and strength associated with aging in mice. J. Ginseng Res. 2022, 46, 657–665. [Google Scholar] [CrossRef]
- Kang, K.A.; Piao, M.J.; Fernando, P.D.S.M.; Herath, H.M.U.L.; Yi, J.M.; Hyun, J.W. Korean red ginseng attenuates particulate matter-induced senescence of skin keratinocytes. Antioxidants 2023, 12, 1516. [Google Scholar] [CrossRef]
- Ben-Porath, I.; Weinberg, R.A. The signals and pathways activating cellular senescence. Int. J. Biochem. Cell Biol. 2005, 37, 961–976. [Google Scholar] [CrossRef] [PubMed]
- Van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Menthena, A.; Koehler, C.I.; Sandhu, J.S.; Yovchev, M.I.; Hurston, E.; Shafritz, D.A.; Oertel, M. Activin A, p15INK4b signaling, and cell competition promote stem/progenitor cell repopulation of livers in aging rats. Gastroenterology 2011, 140, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Farr, J.N. Cellular senescence in age-related disorders. Transl. Res. 2020, 226, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Xia, H.; Guo, Y.; Qian, X.; Zou, X.; Yang, H.; Yin, M.; Liu, H. Ginsenoside Rb1 retards aging process by regulating cell cycle, apoptotic pathway and metabolism of aging mice. J. Ethnopharmacol. 2020, 255, 112746. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Adam, V.; Nepovimova, E.; Heger, Z.; Valko, M.; Wu, Q.; Wei, W.; Kuca, K. c-Jun N-terminal kinase signaling in cellular senescence. Arch Toxicol. 2023, 97, 2089–2109. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, R.; Wang, H.; Liang, F. Mechanisms linking inflammation to insulin resistance. Int. J. Endocrinol. 2015, 2015, 508409. [Google Scholar] [CrossRef]
- Wang, M.C.; Bohmann, D.; Jasper, H. JNK extends life span and limits growth by antagonizing cellular and organism-wide responses to insulin signaling. Cell 2005, 121, 115–125. [Google Scholar] [CrossRef]
- Karpac, J.; Jasper, H. Insulin and JNK: Optimizing metabolic homeostasis and lifespan. Trends Endocrinol. Metab. 2009, 20, 100–106. [Google Scholar] [CrossRef]
- Kim, J.K.; Shin, K.K.; Kim, H.; Hong, Y.H.; Choi, W.; Kwak, Y.-S.; Han, C.-K.; Hyun, S.H.; Cho, J.Y. Korean Red Ginseng exerts anti-inflammatory and autophagy-promoting activities in aged mice. J. Ginseng Res. 2021, 45, 717–725. [Google Scholar] [CrossRef]
- Xu, K.; Hu, B.; Ding, X.; Zhan, Z. Alleviation of D-gal-induced senile liver injury by Rg3, a signature component of red ginseng. Aging 2023, 15, 6749. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.-Q.; Zhou, Y.-D.; Hou, J.-G.; Liu, Y.; Wang, Y.-P.; Gong, X.-J.; Lin, X.-H.; Jiang, S.; Wang, Z. Rare ginsenoside 20 (R)-Rg3 inhibits D-galactose-induced liver and kidney injury by regulating oxidative stress-induced apoptosis. Am. J. Chin. Med. 2020, 48, 1141–1157. [Google Scholar] [CrossRef]
- Park, S.K.; Kim, S.-W.; Seo, H.W.; Hyun, S.H.; Kyung, J.-S.; Youn, S.-H.; So, S.-H.; In, G.; Park, C.-K.; Yi, E.C.; et al. Long-term evaluation of safety and biological effects of Korean Red Ginseng (Panax Ginseng): A long-term in vivo study. BMC Complement. Med. Ther. 2022, 22, 284. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-Y.; Arndt, J.; O’Connell, J.F.; Egan, J.M.; Kim, Y. Red Ginseng Attenuates the Hepatic Cellular Senescence in Aged Mice. Biology 2024, 13, 36. https://doi.org/10.3390/biology13010036
Lee D-Y, Arndt J, O’Connell JF, Egan JM, Kim Y. Red Ginseng Attenuates the Hepatic Cellular Senescence in Aged Mice. Biology. 2024; 13(1):36. https://doi.org/10.3390/biology13010036
Chicago/Turabian StyleLee, Da-Yeon, Juliana Arndt, Jennifer F. O’Connell, Josephine M. Egan, and Yoo Kim. 2024. "Red Ginseng Attenuates the Hepatic Cellular Senescence in Aged Mice" Biology 13, no. 1: 36. https://doi.org/10.3390/biology13010036
APA StyleLee, D. -Y., Arndt, J., O’Connell, J. F., Egan, J. M., & Kim, Y. (2024). Red Ginseng Attenuates the Hepatic Cellular Senescence in Aged Mice. Biology, 13(1), 36. https://doi.org/10.3390/biology13010036