
Cellulolytic Aerobic Bacteria Isolated from Agricultural and Forest Soils: An Overview

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Soil Cellulolytic Bacteria and Methods for Their Identification
3. Methods for Qualitative and Quantitative Determination of Cellulase Activity
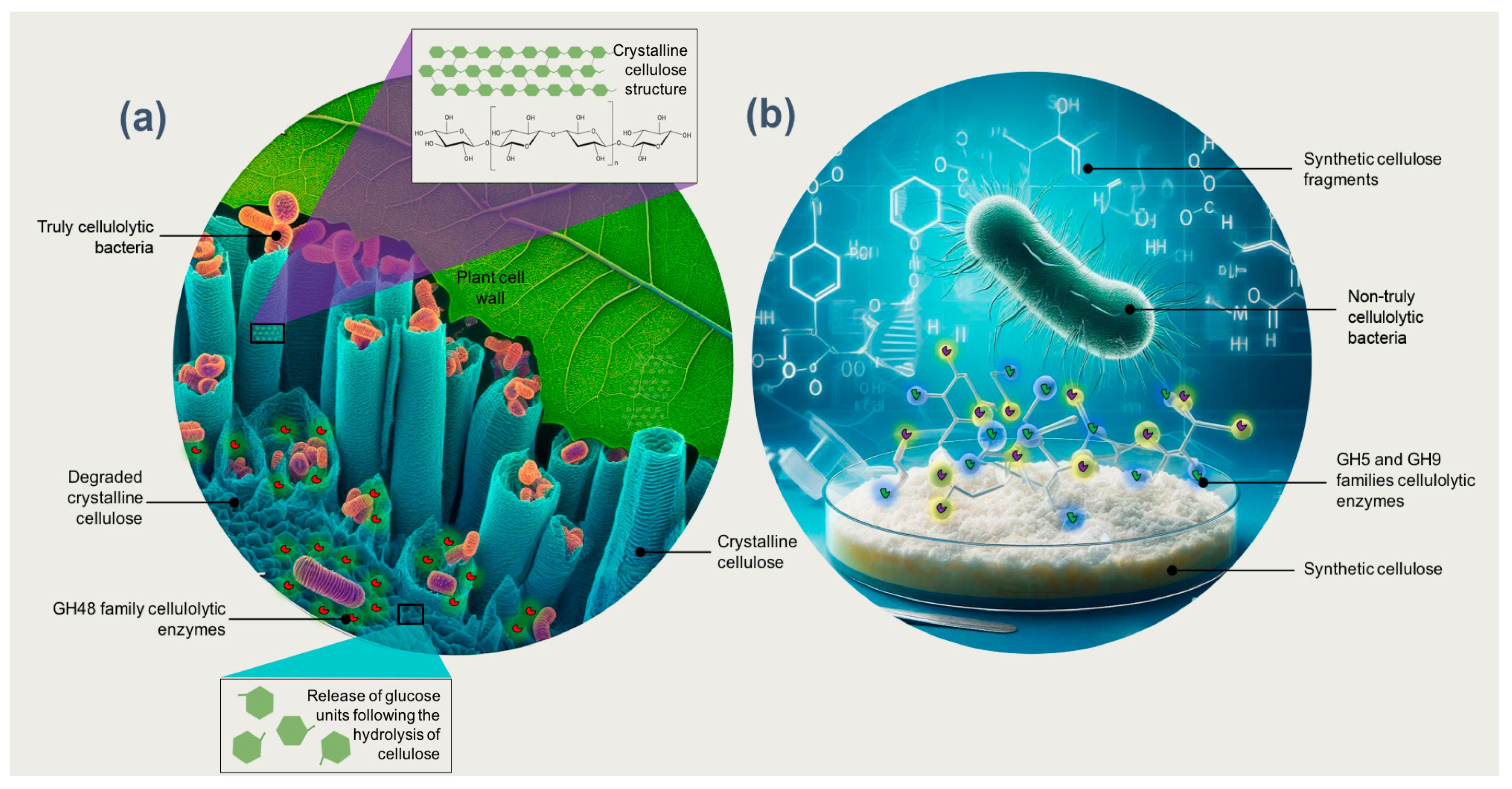
4. Truly Cellulolytic Bacteria
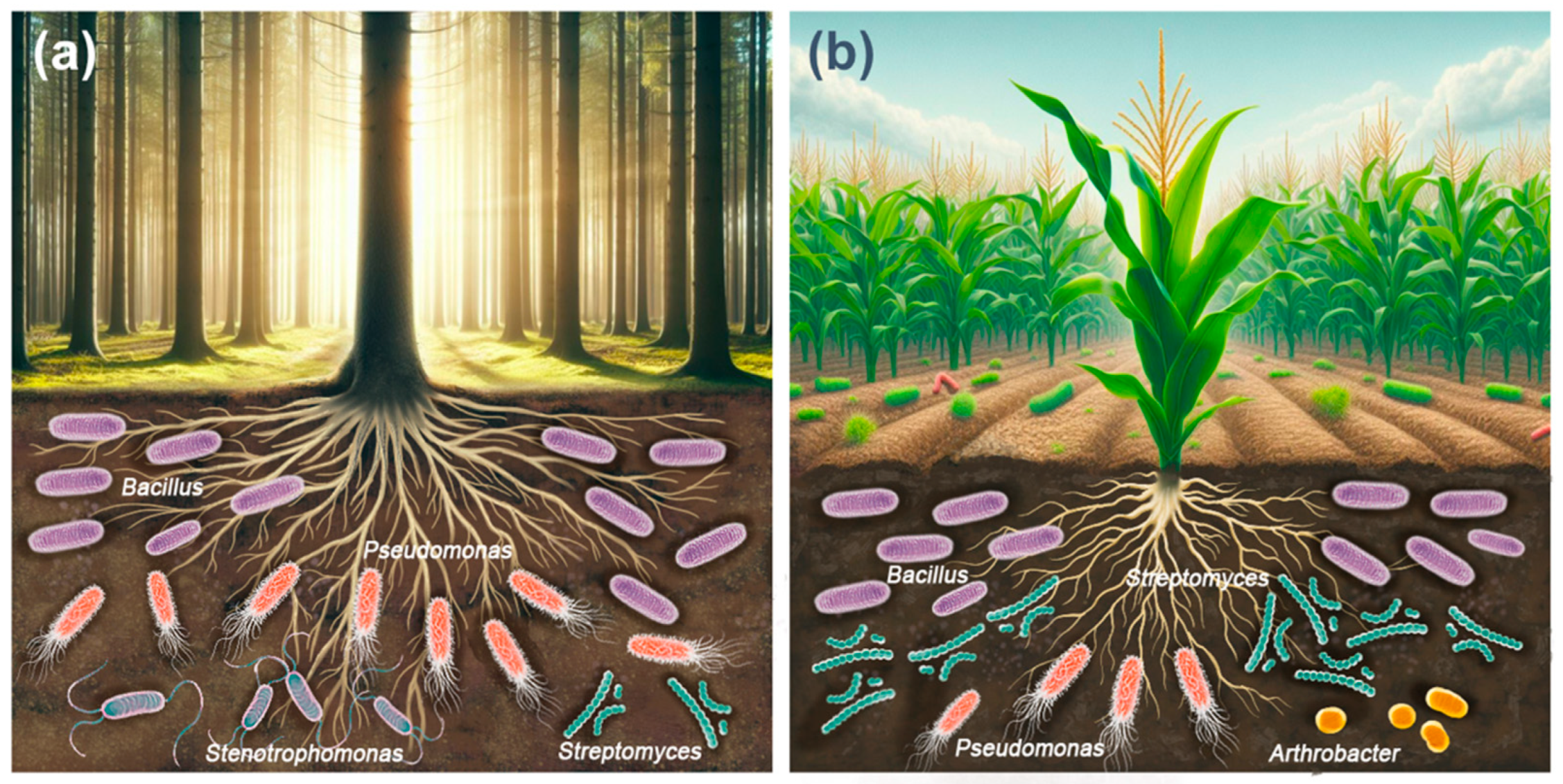
5. Cellulolytic Aerobic Bacteria in Forest Soils
6. Cellulolytic Aerobic Bacteria in Agricultural Soils
7. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Himanshu; Saini, J.K. Diversity and Biotechnological Importance of Cellulolytic Microorganisms from Biodiversity Hotspots. In Microbial Diversity in Hotspots; Academic Press: Cambridge, MA, USA, 2022; pp. 207–230. [Google Scholar] [CrossRef]
- Rettenmaier, R.; Lo, Y.K.; Schmidt, L.; Munk, B.; Lagkouvardos, I.; Neuhaus, K.; Schwarz, W.; Liebl, W.; Zverlov, V. A Novel Primer Mixture for Gh48 Genes: Quantification and Identification of Truly Cellulolytic Bacteria in Biogas Fermenters. Microorganisms 2020, 8, 1297. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Yasir, S.; Khan, M.S.; Manan, S.; Ullah, M.W.; Ul-Islam, M. Plant Extract-Loaded Bacterial Cellulose Composite Membrane for Potential Biomedical Applications. J. Bioresour. Bioprod. 2021, 6, 26–32. [Google Scholar] [CrossRef]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of Global Forest Area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Lebeis, S.L. Greater than the Sum of Their Parts: Characterizing Plant Microbiomes at the Community-Level. Curr. Opin. Plant Biol. 2015, 24, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Berg, B.; Laskowski, R. Litter Decomposition: A Guide to Carbon and Nutrient Turnover. In Advances in Ecological Research; Caswell, H., Ed.; Academic Press: Cambridge, MA, USA, 2006; Volume 38, pp. 18–71. [Google Scholar] [CrossRef]
- Ja’afar, J.N.; Shitu, A. Utilization of Lignocellulosic Agro-Waste as an Alternative Carbon Source for Industrial Enzyme Production. In Waste Management, Processing and Valorisation; Yaser, A.Z., Tajarudin, H.A., Embrandiri, A., Eds.; Springer: Singapore, 2022; pp. 221–233. [Google Scholar] [CrossRef]
- Periyasamy, S.; Karthik, V.; Senthil Kumar, P.; Isabel, J.B.; Temesgen, T.; Hunegnaw, B.M.; Melese, B.B.; Mohamed, B.A.; Vo, D.-V.N. Chemical, Physical and Biological Methods to Convert Lignocellulosic Waste into Value-Added Products. A Review. Environ. Chem. Lett. 2022, 20, 1129–1152. [Google Scholar] [CrossRef]
- Schnitzer, M.; Monreal, C.M.; Powell, E.E. Wheat Straw Biomass: A Resource for High-Value Chemicals. J. Environ. Sci. Health Part B 2014, 49, 51–67. [Google Scholar] [CrossRef]
- Haghighi Mood, S.; Hossein Golfeshan, A.; Tabatabaei, M.; Salehi Jouzani, G.; Najafi, G.H.; Gholami, M.; Ardjmand, M. Lignocellulosic Biomass to Bioethanol, a Comprehensive Review with a Focus on Pretreatment. Renew. Sustain. Energy Rev. 2013, 27, 77–93. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest Soil Bacteria: Diversity, Involvement in Ecosystem Processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef]
- Habib, M.; Khatun, F.; Yasmin, S.; Rahman, A. Screening, Isolation and Characterization of Cellulolytic Bacteria from Soil Samples of Different Agro Industrial Area of Bangladesh and Their Optimization for Cellulase Enzyme Production. J. Bangladesh Agric. Univ. 2021, 19, 30–37. [Google Scholar] [CrossRef]
- Fuhrmann, J.J.; Zuberer, D.A. Carbon Transformations and Soil Organic Matter Formation. In Principles and Applications of Soil Microbiology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 327–361. [Google Scholar] [CrossRef]
- Khokhar, I.; Haider, M.S.; Mushtaq, S.; Mukhtar, I. Isolation and Screening of Highly Cellulolytic Filamentous Fungi. Sch. J. Agric. Sci. 2012, 7, 126–129. [Google Scholar]
- Gupta, P.; Samant, K.; Sahu, A. Isolation of Cellulose-Degrading Bacteria and Determination of Their Cellulolytic Potential. Int. J. Microbiol. 2012, 2012, 578925. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.L.; Zhang, Z.; Wu, M.; Wu, Y.; Feng, J.X. Isolation, Screening, and Identification of Cellulolytic Bacteria from Natural Reserves in the Subtropical Region of China and Optimization of Cellulase Production by Paenibacillus Terrae ME27-1. BioMed Res. Int. 2014, 2014, 512497. [Google Scholar] [CrossRef] [PubMed]
- Brás, J.L.A.; Cartmell, A.; Carvalho, A.L.M.; Verzé, G.; Bayer, E.A.; Vazana, Y.; Correia, M.A.S.; Prates, J.A.M.; Ratnaparkhe, S.; Boraston, A.B.; et al. Structural Insights into a Unique Cellulase Fold and Mechanism of Cellulose Hydrolysis. Proc. Natl. Acad. Sci. USA 2011, 108, 5237–5242. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.B. Cellulases. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 252–258. [Google Scholar] [CrossRef]
- Lo, Y.C.; Saratale, G.D.; Chen, W.M.; Bai, M.D.; Chang, J.S. Isolation of Cellulose-Hydrolytic Bacteria and Applications of the Cellulolytic Enzymes for Cellulosic Biohydrogen Production. Enzyme Microb. Technol. 2009, 44, 417–425. [Google Scholar] [CrossRef]
- Maki, M.; Leung, K.T.; Qin, W. The Prospects of Cellulase-Producing Bacteria for the Bioconversion of Lignocellulosic biomass. Int. J. Biol. Sci. 2009, 5, 500–516. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.B. Microbial Diversity of Cellulose Hydrolysis. Curr. Opin. Microbiol. 2011, 14, 259–263. [Google Scholar] [CrossRef]
- Evdokimova, E.V.; Gladkov, G.V.; Kuzina, N.I.; Ivanova, E.A.; Kimeklis, A.K.; Zverev, A.O.; Kichko, A.A.; Aksenova, T.S.; Pinaev, A.G.; Andronov, E.E. The Difference between Cellulolytic ‘Culturomes’ and Microbiomes Inhabiting Two Contrasting Soil Types. PLoS ONE 2020, 15, e0242060. [Google Scholar] [CrossRef]
- Singh, A.; Bajar, S.; Devi, A.; Pant, D. An Overview on the Recent Developments in Fungal Cellulase Production and Their Industrial Applications. Bioresour. Technol. Rep. 2021, 14, 100652. [Google Scholar] [CrossRef]
- Singh, S.; Jaiswal, D.K.; Sivakumar, N.; Verma, J.P. Developing Efficient Thermophilic Cellulose Degrading Consortium for Glucose Production From Different Agro-Residues. Front. Energy Res. 2019, 7, 61. [Google Scholar] [CrossRef]
- Saratale, G.D.; Saratale, R.G.; Oh, S.E. Production and Characterization of Multiple Cellulolytic Enzymes by Isolated Streptomyces sp. MDS. Biomass Bioenergy 2012, 47, 302–315. [Google Scholar] [CrossRef]
- Balla, A.; Silini, A.; Cherif-Silini, H.; Bouket, A.C.; Boudechicha, A.; Luptakova, L.; Alenezi, F.N.; Belbahri, L. Screening of Cellulolytic Bacteria from Various Ecosystems and Their Cellulases Production under Multi-Stress Conditions. Catalysts 2022, 12, 769. [Google Scholar] [CrossRef]
- Sadhu, S.; Maiti, T.K. Cellulase Production by Bacteria: A Review. Microbiol. Res. J. Int. 2013, 3, 235–258. [Google Scholar] [CrossRef]
- Malik, W.A.; Javed, S. Biochemical Characterization of Cellulase From Bacillus Subtilis Strain and Its Effect on Digestibility and Structural Modifications of Lignocellulose Rich Biomass. Front. Bioeng. Biotechnol. 2021, 9, 800265. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, S.A.; Kokitkar, S.S. Isolation and Identification of Bacteria with Cellulose-Degrading Potential from Soil and Optimization of Cellulase Production. J. Appl. Biol. Biotechnol. 2021, 9, 154–161. [Google Scholar] [CrossRef]
- Grevitara, Y.; Rahma, B.; Septirangga, H.; Dahlia, I.; Suarsini, E. Isolation and Identification of Cellulose Degrading Bacteria from Banana Peel Compost. El-Hayah 2019, 7, 6–11. [Google Scholar] [CrossRef]
- Koeck, D.E.; Pechtl, A.; Zverlov, V.V.; Schwarz, W.H. Genomics of Cellulolytic Bacteria. Curr. Opin. Biotechnol. 2014, 29, 171–183. [Google Scholar] [CrossRef]
- Setiawati, M.R.; Ulfah, N.; Suryatmana, P.; Hindersah, R. Peran Mikroba Dekomposer Selulolitik Dari Sarang Rayap Dalam Menurunkan Kandungan Selulosa Limbah Pertanian Berselulosa Tinggi. Soilrens 2019, 17, 1–8. [Google Scholar] [CrossRef]
- Shamshitov, A.; Decorosi, F.; Viti, C.; Fornasier, F.; Kadžienė, G.; Supronienė, S. Characterisation of Cellulolytic Bacteria Isolated from Agricultural Soil in Central Lithuania. Sustainability 2023, 15, 598. [Google Scholar] [CrossRef]
- Ulrich, A.; Klimke, G.; Wirth, S. Diversity and Activity of Cellulose-Decomposing Bacteria, Isolated from a Sandy and a Loamy Soil after Long-Term Manure Application. Microb. Ecol. 2008, 55, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Cabrera, Y.; Ponce-Mendoza, A.; Vásquez-Murrieta, M.S.; Rivera-Orduña, F.N.; Wang, E.T. Diverse Cellulolytic Bacteria Isolated from the High Humus, Alkaline-Saline Chinampa Soils. Ann. Microbiol. 2013, 63, 779–792. [Google Scholar] [CrossRef]
- Berlemont, R.; Allison, S.D.; Weihe, C.; Lu, Y.; Brodie, E.L.; Martiny, J.B.H.; Martiny, A.C. Cellulolytic Potential under Environmental Changes in Microbial Communities from Grassland Litter. Front. Microbiol. 2014, 5, 639. [Google Scholar] [CrossRef]
- Jiménez, D.J.; Dini-Andreote, F.; van Elsas, J.D. Metataxonomic Profiling and Prediction of Functional Behaviour of Wheat Straw Degrading Microbial Consortia. Biotechnol. Biofuels 2014, 7, 92. [Google Scholar] [CrossRef]
- Ward, D.M.; Weller, R.; Bateson, M.M. 16S RRNA Sequences Reveal Numerous Uncultured Microorganisms in a Natural Community. Nature 1990, 345, 63–65. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of Culture-Independent Studies on the Emerging Phylogenetic View of Bacterial Diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [CrossRef]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S RRNA Gene Sequencing for Species and Strain-Level Microbiome Analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed]
- Lebuhn, M.; Hanreich, A.; Klocke, M.; Schlüter, A.; Bauer, C.; Pérez, C.M. Towards Molecular Biomarkers for Biogas Production from Lignocellulose-Rich Substrates. Anaerobe 2014, 29, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, G. Plant Cell Wall Degradation in Insects: Recent Progress on Endogenous Enzymes Revealed by Multi-Omics Technologies. In Advances in Insect Physiology; Jurenka, R., Ed.; Academic Press: Johnston, IA, USA, 2019; Volume 57, pp. 97–136. [Google Scholar] [CrossRef]
- Sardar, P.; Šustr, V.; Chroňáková, A.; Lorenc, F. Metatranscriptomic Holobiont Analysis of Carbohydrate-Active Enzymes in the Millipede Telodeinopus Aoutii (Diplopoda, Spirostreptida). Front. Ecol. Evol. 2022, 10, 931986. [Google Scholar] [CrossRef]
- Kaster, A.K.; Sobol, M.S. Microbial Single-Cell Omics: The Crux of the Matter. Appl. Microbiol. Biotechnol. 2020, 104, 8209–8220. [Google Scholar] [CrossRef] [PubMed]
- Avellaneda-Torres, L.M.; Pulido, C.P.G.; Rojas, E.T. Assessment of Cellulolytic Microorganisms in Soils of Nevados Park, Colombia. Braz. J. Microbiol. 2014, 45, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, C.W.; Doyle, J.D.; Hugley, B. A New Solid Medium for Enumerating Cellulose-Utilizing Bacteria in Soil. Appl. Environ. Microbiol. 1995, 61, 2016–2019. [Google Scholar] [CrossRef] [PubMed]
- Teather, R.M.; Wood, P.J. Use of Congo Red-Polysaccharide Interactions in Enumeration and Characterization of Cellulolytic Bacteria from the Bovine Rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar] [CrossRef]
- Wood, P.J.; Erfle, J.D.; Teather, R.M. Use of Complex Formation between Congo Red and Polysaccharides in Detection and Assay of Polysaccharide Hydrolases. Methods Enzymol. 1988, 160, 59–74. [Google Scholar] [CrossRef]
- Grata, K. Determining Cellulolytic Activity of Microorganisms. Chem. Didact. Ecol. Metrol. 2020, 25, 133–143. [Google Scholar] [CrossRef]
- Mandels, M.; Andreotti, R.; Roche, C. Measurement of Saccharifying Cellulase. Biotechnol. Bioeng. Symp. 1976, 6, 21–33. [Google Scholar]
- Ghose, T.K. Measurement of Cellulase Activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Coward-Kelly, G.; Aiello-Mazzari, C.; Kim, S.; Granda, C.; Holtzapple, M. Suggested Improvements to the Standard Filter Paper Assay Used to Measure Cellulase Activity. Biotechnol. Bioeng. 2003, 82, 745–749. [Google Scholar] [CrossRef]
- Becker, J.M.; Caldwell, G.A.; Zachgo, E.A. Protein Assays. In Biotechnology: A Laboratory Course, 2nd ed.; Becker, J.M., Caldwell, G.A., Zachgo, E.A., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 119–124. [Google Scholar] [CrossRef]
- Khadka, S.; Khadka, D.; Poudel, R.C.; Bhandari, M.; Baidya, P.; Sijapati, J.; Maharjan, J. Production Optimization and Biochemical Characterization of Cellulase from Geobacillus sp. KP43 Isolated from Hot Spring Water of Nepal. BioMed Res. Int. 2022, 2022, 6840409. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Du, W.; Zhong, L. Probing the Interaction Between Cellulose and Cellulase with a Nanomechanical Sensor. In Cellulose–Medical, Pharmaceutical and Electronic Applications; van de Ven, T., Godbout, L., Eds.; Intech Open: London, UK, 2013; pp. 125–140. [Google Scholar] [CrossRef]
- Yunus, G.; Kuddus, M. Cold-Active Microbial Cellulase: Novel Approach To Understand Mechanism and Its Applications in Food and Beverages Industry. J. Microbiol. Biotechnol. Food Sci. 2021, 10, 524–530. [Google Scholar] [CrossRef]
- Fujita, Y.; Takahashi, S.; Ueda, M.; Tanaka, A.; Okada, H.; Morikawa, Y.; Kawaguchi, T.; Arai, M.; Fukuda, H.; Kondo, A. Direct and Efficient Production of Ethanol from Cellulosic Material with a Yeast Strain Displaying Cellulolytic Enzymes. Appl. Environ. Microbiol. 2002, 68, 5136–5141. [Google Scholar] [CrossRef] [PubMed]
- Shuangqi, T.; Zhenyu, W.; Ziluan, F.; Lili, Z.; Jichang, W. Determination Methods of Cellulase Activity. Afr. J. Biotechnol. 2011, 10, 7122–7125. [Google Scholar]
- Hernández-López, A.; Sánchez Félix, D.A.; Zuñiga Sierra, Z.; García Bravo, I.; Dinkova, T.D.; Avila-Alejandre, A.X. Quantification of Reducing Sugars Based on the Qualitative Technique of Benedict. ACS Omega 2020, 5, 32403–32410. [Google Scholar] [CrossRef] [PubMed]
- Lone, M.; Wani, M.; Bhat, N.; Sheikh, S.; Reshi, M. Evaluation of Cellulase Enzyme Secreted by Some Common and Stirring Rhizosphere Fundi of Juglans Regia L. by DNS Method. J. Enzym. Res. 2012, 3, 18–22. [Google Scholar]
- Hendel, B.; Marxsen, J. Viscosimetric Determination of Endocellulase Activity. In Methods to Study Litter Decomposition, a Practical Guide; Bärlocher, F., Gessner, M., Graça, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 405–410. [Google Scholar] [CrossRef]
- Pomeranz, Y. Functional Properties of Food Components, 2nd ed.; Academic Press: San Diego, CA, USA, 1991. [Google Scholar] [CrossRef]
- Nero, G.; Kivirand, K.; Ben Othman, S.; Rinken, T. Amperometric Method for the Determination of Cellulase Activity and Its Optimization Using Response Surface Method. J. Anal. Sci. Technol. 2022, 13, 21. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- dos Santos, T.C.; Filho, G.A.; De Brito, A.R.; Pires, A.J.V.; Bonomo, R.C.F.; Franco, M. Production and Characterization of Cellulolytic Enzymes by Aspergillus niger and Rhizopus sp. by Solid State Fermentation of Prickly Pear. Rev. Caatinga 2016, 29, 222–233. [Google Scholar] [CrossRef]
- Manfredi, A.P.; Pisa, J.H.; Valdeón, D.H.; Perotti, N.I.; Martínez, M.A. Synergistic Effect of Simple Sugars and Carboxymethyl Cellulose on the Production of a Cellulolytic Cocktail from Bacillus sp. AR03 and Enzyme Activity Characterization. Appl. Biochem. Biotechnol. 2016, 179, 16–32. [Google Scholar] [CrossRef]
- Oliveira, L.R.C.; Barbosa, J.B.; Martins, M.L.L.; Martins, M.A. Extracellular Production of Avicelase by the Thermophilic Soil Bacterium Bacillus sp. SMIA-2. Acta Sci. Biol. Sci. 2014, 36, 215–222. [Google Scholar] [CrossRef]
- Chantarasiri, A. Aquatic Bacillus Cereus JD0404 Isolated from the Muddy Sediments of Mangrove Swamps in Thailand and Characterization of Its Cellulolytic Activity. Egypt. J. Aquat. Res. 2015, 41, 257–264. [Google Scholar] [CrossRef]
- Chernoglazov, V.M.; Jafarova, A.N.; Klyosov, A.A. Continuous Photometric Determination of Endo-1,4-Beta-D-Glucanase (Cellulase) Activity Using 4-Methylumbelliferyl-Beta-D-Cellobioside as a Substrate. Anal. Biochem. 1989, 179, 186–189. [Google Scholar] [CrossRef]
- Koubová, A.; Lorenc, F.; Horváthová, T.; Chroňáková, A.; Šustr, V. Millipede Gut-Derived Microbes as a Potential Source of Cellulolytic Enzymes. World J. Microbiol. Biotechnol. 2023, 39, 169. [Google Scholar] [CrossRef]
- Bakare, M.K.; Adewale, I.O.; Ajayi, A.; Shonukan, O.O. Purification and Characterization of Cellulase from the Wild-Type and Two Improved Mutants of Pseudomonas Fluorescens. Afr. J. Biotechnol. 2005, 4, 898–904. [Google Scholar]
- Raju, V.N.E.; Divakar, G. Production of Pectinase by Using Bacillus Circulans Isolated from Dump Yards of Vegetable Wastes. Int. J. Pharm. Sci. Res. 2013, 4, 2615–2622. [Google Scholar] [CrossRef]
- Goyari, S.; Devi, S.S.; Kalita, M.C.; Talukdar, N.C. Population, Diversity and Characteristics of Cellulolytic Microorganisms from the Indo-Burma Biodiversity Hotspot. Springerplus 2014, 3, 700. [Google Scholar] [CrossRef]
- Gutiérrez-Rojas, I.; Moreno-Sarmiento, N.; Montoya, D. Mecanismos y Regulación de La Hidrólisis Enzimática de Celulosa En Hongos Filamentosos: Casos Clásicos y Nuevos Modelos. Rev. Iberoam. Micol. 2015, 32, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, L.; Liu, D. Biomass Recalcitrance. Part I: The Chemical Compositions and Physical Structures Affecting the Enzymatic Hydrolysis of Lignocellulose. Biofuels Bioprod. Biorefin. 2012, 6, 465–482. [Google Scholar] [CrossRef]
- Bichot, A.; Delgenès, J.P.; Méchin, V.; Carrère, H.; Bernet, N.; García-Bernet, D. Understanding Biomass Recalcitrance in Grasses for Their Efficient Utilization as Biorefinery Feedstock. Rev. Environ. Sci. Biotechnol. 2018, 17, 707–748. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhao, S.; Yang, S.; Ding, S.-Y. Lignin Plays a Negative Role in the Biochemical Process for Producing Lignocellulosic Biofuels. Curr. Opin. Biotechnol. 2014, 27, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A.; Mayer-Laigle, C.; Solhy, A.; Arancon, R.A.D.; De Vries, H.; Luque, R. Mechanical Pretreatments of Lignocellulosic Biomass: Towards Facile and Environmentally Sound Technologies for Biofuels Production. RSC Adv. 2014, 4, 48109–48127. [Google Scholar] [CrossRef]
- Vaidya, A.A.; Donaldson, L.A.; Newman, R.H.; Suckling, I.D.; Campion, S.H.; Lloyd, J.A.; Murton, K.D. Micromorphological Changes and Mechanism Associated with Wet Ball Milling of Pinus Radiata Substrate and Consequences for Saccharification at Low Enzyme Loading. Bioresour. Technol. 2016, 214, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Zheng, M.; Li, X.; Sebastian, J.; Jiang, Y.; Zhao, Y.; Wang, A.; Zhang, T. Unlock the Compact Structure of Lignocellulosic Biomass by Mild Ball Milling for Ethylene Glycol Production. ACS Sustain. Chem. Eng. 2019, 7, 679–687. [Google Scholar] [CrossRef]
- Yu, H.; Xiao, W.; Han, L.; Huang, G. Characterization of Mechanical Pulverization/Phosphoric Acid Pretreatment of Corn Stover for Enzymatic Hydrolysis. Bioresour. Technol. 2019, 282, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Jeoh, T.; Ishizawa, C.I.; Davis, M.F.; Himmel, M.E.; Adney, W.S.; Johnson, D.K. Cellulase Digestibility of Pretreated Biomass Is Limited by Cellulose Accessibility. Biotechnol. Bioeng. 2007, 98, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Grethlein, H.E. The Effect of Pore Size Distribution on the Rate of Enzymatic Hydrolysis of Cellulosic Substrates. Bio/Technology 1985, 3, 155–160. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The Carbohydrate-Active Enzyme Database: Functions and Literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Levasseur, A.; Drula, E.; Lombard, V.; Coutinho, P.M.; Henrissat, B. Expansion of the Enzymatic Repertoire of the CAZy Database to Integrate Auxiliary Redox Enzymes. Biotechnol. Biofuels 2013, 6, 41. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Zühlke, D.; Becher, D.; Riedel, K.; Baldrian, P. Cellulose and Hemicellulose Decomposition by Forest Soil Bacteria Proceeds by the Action of Structurally Variable Enzymatic Systems. Sci. Rep. 2016, 6, 25279. [Google Scholar] [CrossRef] [PubMed]
- Brumm, P.J. Bacterial Genomes: What They Teach Us about Cellulose Degradation. Biofuels 2013, 4, 669–681. [Google Scholar] [CrossRef]
- Gaur, R.; Tiwari, S. Isolation, Production, Purification and Characterization of an Organic-Solvent-Thermostable Alkalophilic Cellulase from Bacillus Vallismortis RG-07. BMC Biotechnol. 2015, 15, 19. [Google Scholar] [CrossRef]
- Cragg, S.M.; Beckham, G.T.; Bruce, N.C.; Bugg, T.D.H.; Distel, D.L.; Dupree, P.; Etxabe, A.G.; Goodell, B.S.; Jellison, J.; McGeehan, J.E.; et al. Lignocellulose Degradation Mechanisms across the Tree of Life. Curr. Opin. Chem. Biol. 2015, 29, 108–119. [Google Scholar] [CrossRef]
- Saini, A.; Aggarwal, N.K.; Sharma, A.; Yadav, A. Actinomycetes: A Source of Lignocellulolytic Enzymes. Enzyme Res. 2015, 2015, 279381. [Google Scholar] [CrossRef]
- Lamed, R.; Setter, E.; Bayer, E.A. Characterization of a Cellulose-Binding, Cellulase-Containing Complex in Clostridium Thermocellum. J. Bacteriol. 1983, 156, 828–836. [Google Scholar] [CrossRef]
- Zverlov, V.V.; Hiegl, W.; Köck, D.E.; Kellermann, J.; Köllmeier, T.; Schwarz, W.H. Hydrolytic Bacteria in Mesophilic and Thermophilic Degradation of Plant Biomass. Eng. Life Sci. 2010, 10, 528–536. [Google Scholar] [CrossRef]
- Becher, D.; Bernhardt, J.; Fuchs, S.; Riedel, K. Metaproteomics to Unravel Major Microbial Players in Leaf Litter and Soil Environments: Challenges and Perspectives. Proteomics 2013, 13, 2895–2909. [Google Scholar] [CrossRef] [PubMed]
- Takasuka, T.E.; Book, A.J.; Lewin, G.R.; Currie, C.R.; Fox, B.G. Aerobic Deconstruction of Cellulosic Biomass by an Insect-Associated Streptomyces. Sci. Rep. 2013, 3, 1030. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, U.; Louis, P.; Goesmann, A.; Henrissat, B.; Duncan, S.H.; Flint, H.J. Complete Genome of a New Firmicutes Species Belonging to the Dominant Human Colonic Microbiota (‘Ruminococcus Bicirculans’) Reveals Two Chromosomes and a Selective Capacity to Utilize Plant Glucans. Environ. Microbiol. 2014, 16, 2879–2890. [Google Scholar] [CrossRef] [PubMed]
- Vodovnik, M.; Duncan, S.H.; Reid, M.D.; Cantlay, L.; Turner, K.; Parkhill, J.; Lamed, R.; Yeoman, C.J.; Miller, M.E.B.; White, B.A.; et al. Expression of Cellulosome Components and Type IV Pili within the Extracellular Proteome of Ruminococcus Flavefaciens 007. PLoS ONE 2013, 8, e65333. [Google Scholar] [CrossRef]
- Berlemont, R.; Martiny, A.C. Genomic Potential for Polysaccharide Deconstruction in Bacteria. Appl. Environ. Microbiol. 2015, 81, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Větrovský, T.; Steffen, K.T.; Baldrian, P. Potential of Cometabolic Transformation of Polysaccharides and Lignin in Lignocellulose by Soil Actinobacteria. PLoS ONE 2014, 9, e89108. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.P.; Paul, E.A. Decomposition and Soil Organic Matter Dynamics. In Methods in Ecosystem Science; Sala, O.E., Jackson, R.B., Mooney, H.A., Howarth, R.W., Eds.; Springer: New York, NY, USA, 2000; pp. 104–116. [Google Scholar] [CrossRef]
- Viteri Florez, P.A.; Castillo Guerra, D.A.; Viteri Rosero, S.E. Capacidad y diversidad de bacterias celulolíticas aisladas de tres hábitats tropicales en Boyacá, Colombia. Acta Agronómica 2016, 65, 362–367. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Nwachukwu, B.C.; Ayangbenro, A.S.; Babalola, O.O. Elucidating the Rhizosphere Associated Bacteria for Environmental Sustainability. Agriculture 2021, 11, 75. [Google Scholar] [CrossRef]
- Khotimah, S.; Suharjono; Ardyati, T.; Nurani, Y. Isolation and Identification of Cellulolytic Bacteria at Fibric, Hemic and Sapric Peat in Teluk Bakung Peatland, Kubu Raya District, Indonesia. Biodiversitas 2020, 21, 2103–2112. [Google Scholar] [CrossRef]
- Tang, A.; Haruna, A.O.; Majid, N.M.A.; Jalloh, M.B. Potential PGPR Properties of Cellulolytic, Nitrogen-Fixing, Phosphate-Solubilizing Bacteria in Rehabilitated Tropical Forest Soil. Microorganisms 2020, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Saber, M.A.; Tripty, I.A.; Karim, M.A.; Islam, M.A.; Hasan, M.S.; Alam, A.S.M.R.U.; Jahid, M.I.K.; Hasan, M.N. Molecular Characterization of Cellulolytic (Endo- and Exoglucanase) Bacteria from the Largest Mangrove Forest (Sundarbans), Bangladesh. Ann. Microbiol. 2020, 70, 68. [Google Scholar] [CrossRef]
- Bontemps, C.; Toussaint, M.; Revol, P.-V.; Hotel, L.; Jeanbille, M.; Uroz, S.; Turpault, M.-P.; Blaudez, D.; Leblond, P. Taxonomic and Functional Diversity of Streptomyces in a Forest Soil. FEMS Microbiol. Lett. 2013, 342, 157–167. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Singh, R.; Eltis, L.D.; Mohn, W.W. Bacterial Contributions to Delignification and Lignocellulose Degradation in Forest Soils with Metagenomic and Quantitative Stable Isotope Probing. ISME J. 2019, 13, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Lu, Y.; Yan, H.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Screening of Cellulolytic Bacteria from Rotten Wood of Qinling (China) for Biomass Degradation and Cloning of Cellulases from Bacillus Methylotrophicus. BMC Biotechnol. 2020, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Ghio, S.; Lorenzo, G.S.D.; Lia, V.; Talia, P.; Cataldi, A.; Grasso, D.; Campos, E. Isolation of Paenibacillus sp. and Variovorax sp. Strains from Decaying Woods and Characterization of Their Potential for Cellulose Deconstruction. Int. J. Biochem. Mol. Biol. 2012, 3, 352–364. [Google Scholar] [PubMed]
- Ghio, S.; Martinez Cáceres, A.I.; Talia, P.; Grasso, D.H.; Campos, E. Draft Genome Sequence of Cellulolytic and Xylanolytic Paenibacillus sp. A59, Isolated from Decaying Forest Soil from Patagonia, Argentina. Genome Announc. 2015, 3, e01233-15. [Google Scholar] [CrossRef]
- Ashwani, K.; Saida, L.; Reddy, K.V. Characterization of Cellulolytic Bacteria from Forest Soil Sample. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 679–685. [Google Scholar]
- Schlösser, A.; Aldekamp, T.; Schrempf, H. Binding Characteristics of CebR, the Regulator of the Ceb Operon Required for Cellobiose/Cellotriose Uptake in Streptomyces Reticuli. FEMS Microbiol. Lett. 2000, 190, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Kolařík, M.; Stursová, M.; Kopecký, J.; Valášková, V.; Větrovský, T.; Zifčáková, L.; Snajdr, J.; Rídl, J.; Vlček, C.; et al. Active and Total Microbial Communities in Forest Soil Are Largely Different and Highly Stratified during Decomposition. ISME J. 2012, 6, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N. Embracing the Unknown: Disentangling the Complexities of the Soil Microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Prescott, C.E.; Vesterdal, L. Decomposition and Transformations along the Continuum from Litter to Soil Organic Matter in Forest Soils. For. Ecol. Manag. 2021, 498, 119522. [Google Scholar] [CrossRef]
- Dobrzyński, J.; Wierzchowski, P.S.; Stępień, W.; Górska, E.B. The Reaction of Cellulolytic and Potentially Cellulolytic Spore-Forming Bacteria to Various Types of Crop Management and Farmyard Manure Fertilization in Bulk Soil. Agronomy 2021, 11, 772. [Google Scholar] [CrossRef]
- Dobrzyński, J.; Wróbel, B.; Górska, E.B. Cellulolytic Properties of a Potentially Lignocellulose-Degrading Bacillus sp. 8E1A Strain Isolated from Bulk Soil. Agronomy 2022, 12, 665. [Google Scholar] [CrossRef]
- Elkhalil, E.A.I.; Gaffar, F.Y.; Siddig, M.A.E.; Osman, H.A.H. Isolation and Molecular Characterization of Cellulolytic Bacillus Isolates from Soil and Compost. Am. J. Microbiol. Res. 2015, 3, 55–58. [Google Scholar]
- Sharma, V.; Vij, H.; Kumar, P.; Sheelendra, S. Potential Cellulase Production, Optimization and Sachharification Study by Novel Thermophilic Microbes. J. Sustain. Biotechnol. 2013, 1, 19–26. [Google Scholar]
- Abdel-Aziz, S.H.; Ibrahim, A.M.; Guirgis, A.; Dawwam, G.E.; Elsababty, Z.E. Isolation and Screening of Cellulase Producing Bacteria Isolated from Soil. Benha J. Appl. Sci. 2021, 6, 207–213. [Google Scholar] [CrossRef]
- Dias, P.V.S.; Ramos, K.O.; Padilha, I.Q.M.; Araújo, D.A.M.; Santos, S.F.M.; Silva, F.L.H. Optimization of Cellulase Production by Bacillus sp. Isolated from Sugarcane Cultivated Soil. Chem. Eng. Trans. 2014, 38, 277–282. [Google Scholar] [CrossRef]
- Guzmán Cedeño, Á.M.; Zambrano Pazmiño, D.E.; Rivera Fernández, R.D.; Rondón, A.J.; Silva, M.L.; Quintana, M.P. Isolation and Selection of Autochthonous Bacteria from Manabí-Ecuador with Cellulolytic Activity. Cultiv. Trop. 2015, 36, 7–16. [Google Scholar]
- Máté, R.; Kutasi, J.; Bata-Vidács, I.; Kosztik, J.; Kukolya, J.; Tóth, E.; Bóka, K.; Táncsics, A.; Kovács, G.; Nagy, I.; et al. Flavobacterium Hungaricum sp. Nov. a Novel Soil Inhabitant, Cellulolytic Bacterium Isolated from Plough Field. Arch. Microbiol. 2022, 204, 301. [Google Scholar] [CrossRef]
- Susilowati, D.; Sudiana, I.M.; Mubarik, N.; Suwanto, A. Species and Functional Diversity of Rhizobacteria of Rice Plant in the Coastal Soils of Indonesia. Indones. J. Agric. Sci. 2015, 16, 39–50. [Google Scholar] [CrossRef]
- Ramos-bello, R.; Cajuste, L.J.; Flores-román, D.; García-calderón, N.E. Metales Pesados, Sales y Sodio En Los Suelos de Chinampa En México. Agrociencia 2001, 35, 385–395. [Google Scholar]
- Luque, A.G.; Pioli, R.; Bonel, B.; Alvarez, D.P. Cellulolytic Fungi Populations in Stubble and Soil as Affected by Agricultural Management Practices. Biol. Agric. Hortic. 2005, 23, 121–142. [Google Scholar] [CrossRef]
- Fan, F.; Li, Z.; Wakelin, S.A.; Yu, W.; Liang, Y. Mineral Fertilizer Alters Cellulolytic Community Structure and Suppresses Soil Cellobiohydrolase Activity in a Long-Term Fertilization Experiment. Soil Biol. Biochem. 2012, 55, 70–77. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Isolation Site | Genus/Species | Intensity of Cellulase Activity | Method for Identifying Cellulolytic Bacteria | Carbon Source for Screening Cellulolytic Bacteria | Methods for Analyzing Cellulase Activity | Ref. |
|---|---|---|---|---|---|---|
| Native forests of El Páramo El Malmo El Páramo El Horizonte Iguaque Natural Reserve Iguaque-Arcabuco stretch and Gomeca river basin in the Department of Boyacá, Colombia | Bacillus sp. Erwinia sp. Pseudomonas sp. | +++ ++ ++ | Biochemical testing | Carboxymethylcellulose | Qualitative: Congo red assay | [100] |
| Superpáramo, Páramo, and High Andean Forest in the Nevados National Natural Park, Colombia | Pseudomonas Streptomyces Rhodococcus Stenotrophomonas Variovorax Serratia Janthinobacterium | ++ +++ + + + + + | 16S rRNA gene sequencing | Carboxymethylcellulose | Qualitative: Congo red assay Quantitative: β-glucosidase activity was measured by its absorbance at 405 nm, while endoglucanase and exoglucanase activities were determined using the 3,5-dinitrosalicylic acid (DNS) method. Total cellulase activity was assessed by adding filter paper and phosphate buffer. In all cases the protein content was determined by Bradford method | [45] |
| Forest floor of a temperate oak forest (Quercus petraea), Czech Republic | Pedobacter Mucilaginibacter Luteibacter | ++ +++ + | 16S rRNA gene sequencing * | Carboxymethylcellulose | Qualitative: Congo red assay Quantitative: Exocellulase and β-glucosidase activities were measured using methylumbelliferol (MUF)-based substrates by mass spectrophotometry proteomics | [86] |
| Swamp/peat forests, Indonesia | Bacillus cereus Bacillus stratosphericus | +++ ++ | 16S rRNA gene sequencing | Carboxymethylcellulose | Qualitative: Congo red assay Quantitative: Endoglucanase activity was analyzed using the DNS (3,5-dinitrosalicylic acid) method, which quantifies glucose levels through spectrophotometry at 540 nm | [103] |
| Tropical rehabilitated forest soils, Malaysia | Serratia nematodiphila strain SP6 Serratia marcescens subsp. sakuensis Stenotrophomonas maltophilia strain KNUC605 Bacillus thuringiensis Stenotrophomonas sp. Ellin162 | +++ ++ ++ ++ ++ | Biochemical testing and 16S rRNA gene sequencing | Cellulose microgranular powder | Quantitative: CMCase (carboxymethyl cellulase) activity was determined by quantifying reducing sugars with the Somogyi-Nelson reagent, measured spectrophotometrically at 520 nm | [104] |
| Mangrove forests of the Bagerhat district, Bangladesh | Bacillus sp. Pseudomonas aeruginosa | +++ ++ | Morphological and biochemical testing, along with 16S rRNA gene sequencing | Carboxymethylcellulose | Qualitative: Congo red assay Quantitative: Endoglucanase and exoglucanase activities were analyzed using the DNS method, which quantifies glucose levels through spectrophotometry at 540 nm | [105] |
| Montiers Forest experimental site, French | Streptomyces sp. strain S2n2 Streptomyces sp. strain S8n36 | +++ +++ | 16S rRNA gene sequencing * | Carboxymethylcellulose | Qualitative: Congo red assay | [106] |
| Mature coniferous forest located in northern Ontario, British Columbia, California and Texas | Cellvibrio Janthinobacterium Cytophaga Salinibacterium | +++ ++ + + | 16S rRNA gene sequencing | Carboxymethylcellulose | Stable isotope probing (SIP) integrated with amplicon and shotgun metagenomic techniques | [107] |
| Rotten wood samples, Qinling Mountain in Shaanxi Province, China | Bacillus subtilis B. licheniformis B. megaterium B. methylotrophicus Pseudomonas aeruginosa | +++ ++ + + + | 16S rRNA gene sequencing * | Carboxymethylcellulose or Avicel | Qualitative: Congo red assay Quantitative: FPA (Filter paper activity), activities of CMCase and Avicelase were analyzed using the DNS method, which quantifies glucose levels through spectrophotometry at 540 nm | [108] |
| Forest soil in the Patagonia region, Argentina | Variovorax paradoxus Paenibacillus alvei Pseudomonas jessenii AMBI2391 Stenotrophomonas maltophilia CQ1 Paenibacillus sp. 61724 Bacillus sp. S3.TSA.017 Bacillus sp. A2022 Bacillus arenosi Brevundimonas sp. SOZ3-5041 Bacillus cereus SH 01 Lysinibacillus sphaericus strain DE4 Xanthomonas sp. X1 Achromobacter xylosoxidans X96 Lysinibacillus sp. KB1 | +++ +++ +++ +++ +++ + + + + + +++ + +++ +++ | 16S rRNA gene sequencing | Carboxymethylcellulose | Qualitative: Congo red assay | [109,110] |
| Nallamala forest Srisailam, Kurnool District of Andhrapradesh, India | Bacillus sp. Pseudomonas sp. | Not quantified | Morphological and Biochemical testing | Ashed, acid-washed cellulose powder | Qualitative: Congo red assay | [111] |
| Dump yards, undisturbed garden soil, and undisturbed forest soil, Raigad district, Maharashtra, India | Bacillus subtilis CP053102.1 Bacillus flexus NR_113800.1 Bacillus licheniformis CP034569.1 Bacillus paralicheniformis KY694465.1 | +++ + ++ + | 16S rRNA gene sequencing | Carboxymethylcellulose | Qualitative: Congo red assay Quantitative: CMCase activity was analyzed using the DNS method, and the quantification of released sugar units was performed following Miller’s protocol. | [29] |
| Crop | Site | Genus/Species | Intensity of Cellulase Activity | Method for Identifying Cellulolytic Bacteria | Carbon Source for Screening of Cellulolytic Bacteria | Agricultural Management | Methods for Analyzing Cellulase Activity | Ref. |
|---|---|---|---|---|---|---|---|---|
| Potato (Solanum tuberosum L.) | Shambat, Khartoum North, Sudan | Bacillus | +++ | 16S rRNA gene sequencing | Carboxymethylcellulose | Not specified | Quantitative: CMCase activity was analyzed using the DNS method, with the quantification of released sugar units carried out following Miller’s protocol | [118] |
| Rice (Oryza sativa L.) | Lovely Professional University Campus, India | Bacillus coagulans Geobacillus stearothermophilus | ++ +++ | 16S rRNA gene sequencing | Carboxymethylcellulose | Not specified | Quantitative: Total cellulase activity was determined using the DNS method | [119] |
| Not specified | Menoufia, Egipto | Bacillus licheniformis, KT693282 Bacillus cereus, KT693283 Klebsiella oxytoca, KT693284 | +++ ++ + | Morphological and biochemical testing, along with 16S rRNA gene sequencing | Carboxymethylcellulose | Not specified | Qualitative: Congo red assay | [120] |
| Pea (Pisum sativum) | Akademija, Central Lithuania | Stenotrophomonas rhizophila Arthrobacter pascens Paenarthrobacter nicotinovorans Oerskovia paurometabola Terrabacter carboxydivorans Agromyces cerinus Streptomyces canus Streptomyces argenteolus Bacillus pumilus Bacillus altitudinis Bacillus mobilis Bacillus butanolivorans | + + + + + + + + +++ + + + | Biochemical testing and 16S rRNA gene sequencing | Cellulose powder Carboxymethylcellulose | Plowing and harrowing No-tillage All tillage treatments, including both with and without cover crop conditions Cereal cropping sequences consisting of five-member crop rotations: winter wheat (Triticum aestivum L.)- winter rape (Brassica napus)- spring wheat (Triticum aestivum L.)- spring barley (Hordeum vulgare)- pea (Pisum sativum) | Qualitative: Testing with Congo red and Gram’s iodine solution Quantitative: Activities of endoglucanases and β-glucosidase were determined in 200 mM MES (morpholineptansulfonic acid) solution. The hydrolytic activities were quantified using 4-methylumbelliferyl and 7-amino-4-methylcoumarin as fluorogenic conjugated substrates | [33] |
| Sugar cane (Saccharum officinarum L.) | Santa Rita, Brazil | Bacillus sp. C1AC55.07 | +++ | 16S rRNA gene sequencing | Carboxymethylcellulose | Not specified | Qualitative: Congo red assay Quantitative: CMCase was determined using the DNS method, which involved the quantification of glucose levels via spectrophotometry at a wavelength of 540 nm | [121] |
| Sugar cane (Saccharum officinarum L.) | Manabi, Ecuador | Bacillus sp. AO-19 | +++ | Morphological testing | Carboxymethylcellulose | A decade-old sugarcane monoculture with substantial fibrous waste accumulation | Qualitative: Congo red assay | [122] |
| Corn (Zea mays L.) | Hungary | Flavobacterium hungaricum sp. nov | Not quantified | 16S rRNA gene sequencing | Carboxymethylcellulose | The authors only mention that the soil pH was moderately alkaline and the soil was fertilized | Qualitative: Congo red assay | [123] |
| Alfalfa (Medicago sativa L.) Chard (Beta vulgaris L.) Grass (Arrhenatherum elatius (Linn.) Pressl) Common sorrel (Rumex crispus L.) | Mexico City, Mexico | Microbacterium oxydans Streptomyces anulatus Cellulomonas cellulans Agrobacterium rubi Sphingobium-bacterium Alcaligenes sp. Pseudomonas pseudoalcaligenes Stenotrophomonas maltophilia Pseudomonas mendocina | +++ +++ +++ ++ - - +++ +++ +++ | 16S rRNA gene sequencing * | Cellulose powder | The soils were collected from agricultural production systems known as chinampas. pH 8.0–8.6, 4.6% to 7.5% organic matter, loamy sandy clayey texture | Qualitative: Congo red assay | [35] |
| Rice (Oryza sativa L.) | Indonesia | Bacillus stratosphericus Bacillus amyloliquefaciens Bacillus cereus Bacillus pumilus Citrobacter freundii Pseudomonas pseudoalcaligenes Rhodobacter aestuarii Bacillus marisflavi Pantoea dispersa Streptomyces coelicoflavus Pseudomonas mosselii Rhodococcus ruber Arthrobacter alpinus Streptomyces albidoflavus | Not quantified | 16S rRNA gene sequencing | Carboxymethylcellulose | Not specified | Qualitative: Congo red assay | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bautista-Cruz, A.; Aquino-Bolaños, T.; Hernández-Canseco, J.; Quiñones-Aguilar, E.E. Cellulolytic Aerobic Bacteria Isolated from Agricultural and Forest Soils: An Overview. Biology 2024, 13, 102. https://doi.org/10.3390/biology13020102
Bautista-Cruz A, Aquino-Bolaños T, Hernández-Canseco J, Quiñones-Aguilar EE. Cellulolytic Aerobic Bacteria Isolated from Agricultural and Forest Soils: An Overview. Biology. 2024; 13(2):102. https://doi.org/10.3390/biology13020102
Chicago/Turabian StyleBautista-Cruz, Angélica, Teodulfo Aquino-Bolaños, Jessie Hernández-Canseco, and Evangelina Esmeralda Quiñones-Aguilar. 2024. "Cellulolytic Aerobic Bacteria Isolated from Agricultural and Forest Soils: An Overview" Biology 13, no. 2: 102. https://doi.org/10.3390/biology13020102
APA StyleBautista-Cruz, A., Aquino-Bolaños, T., Hernández-Canseco, J., & Quiñones-Aguilar, E. E. (2024). Cellulolytic Aerobic Bacteria Isolated from Agricultural and Forest Soils: An Overview. Biology, 13(2), 102. https://doi.org/10.3390/biology13020102