
The Effects of Salinity and Genotype on the Rhizospheric Mycobiomes in Date Palm Seedlings



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Sampling, Seed Sterilization, and Plant Growth
2.2. DNA Extraction and Illumina MiSeq Sequencing of ITS
2.3. Bioinformatic Analysis
3. Results
3.1. Metagenomic Analysis Revealed Minor Variations in Fungal Community Structures across Distinct Cultivars Due to Environmental Changes
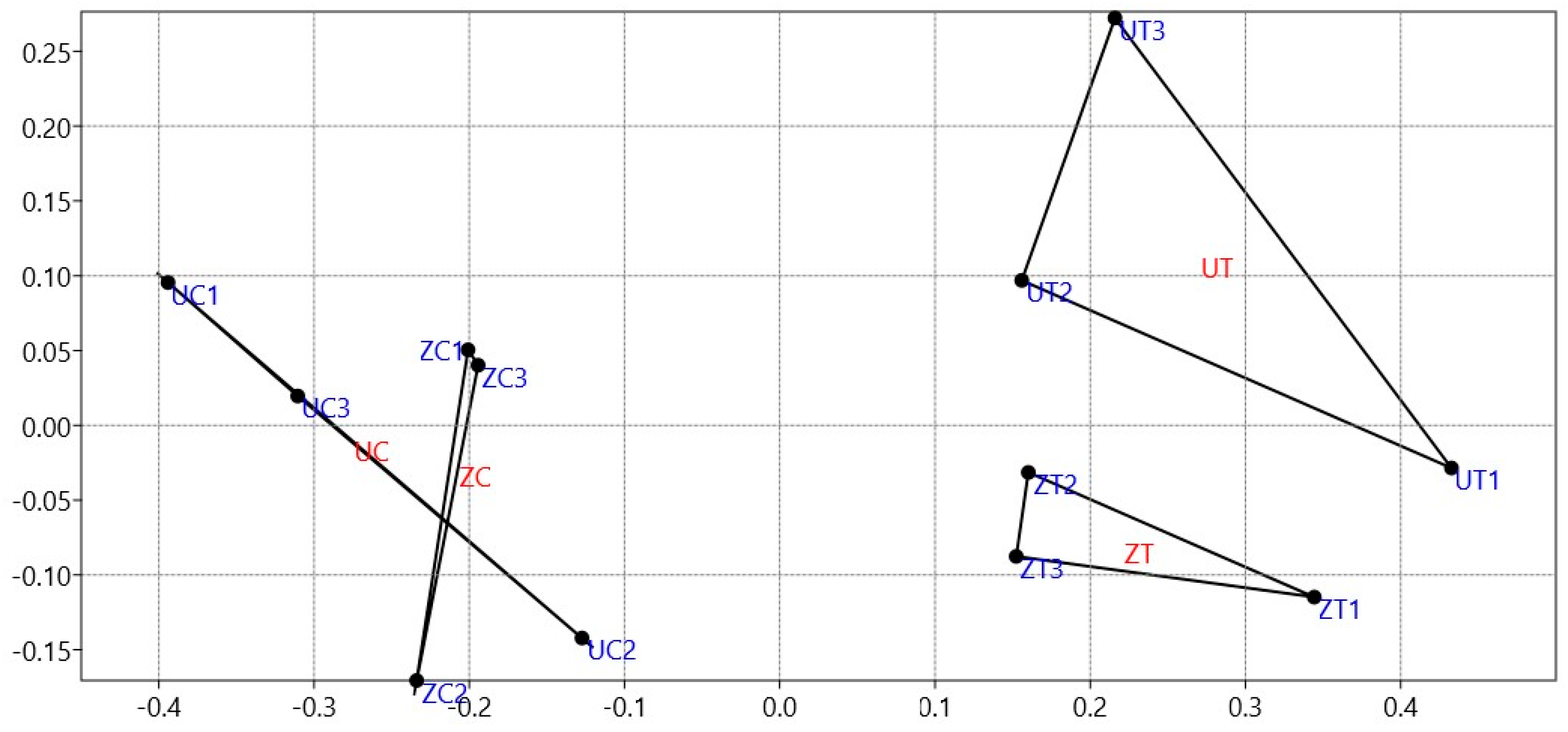
3.2. Multidimensional Analysis Reveals Salinity Impact on Date Palm Rhizofungal Communities
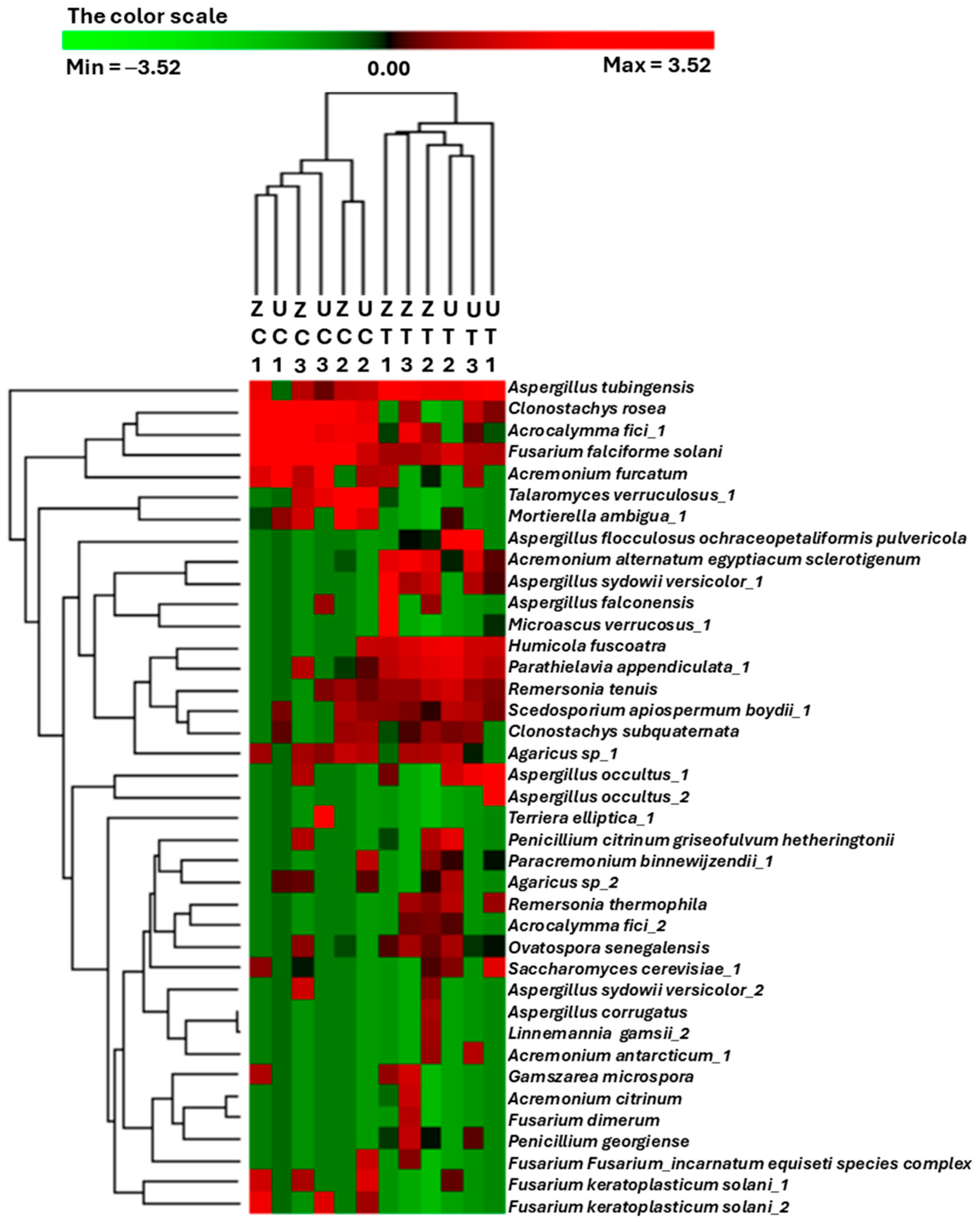
3.3. Comparative Heatmap Analysis Reveals Treatment-Specific Fungal Abundance Patterns
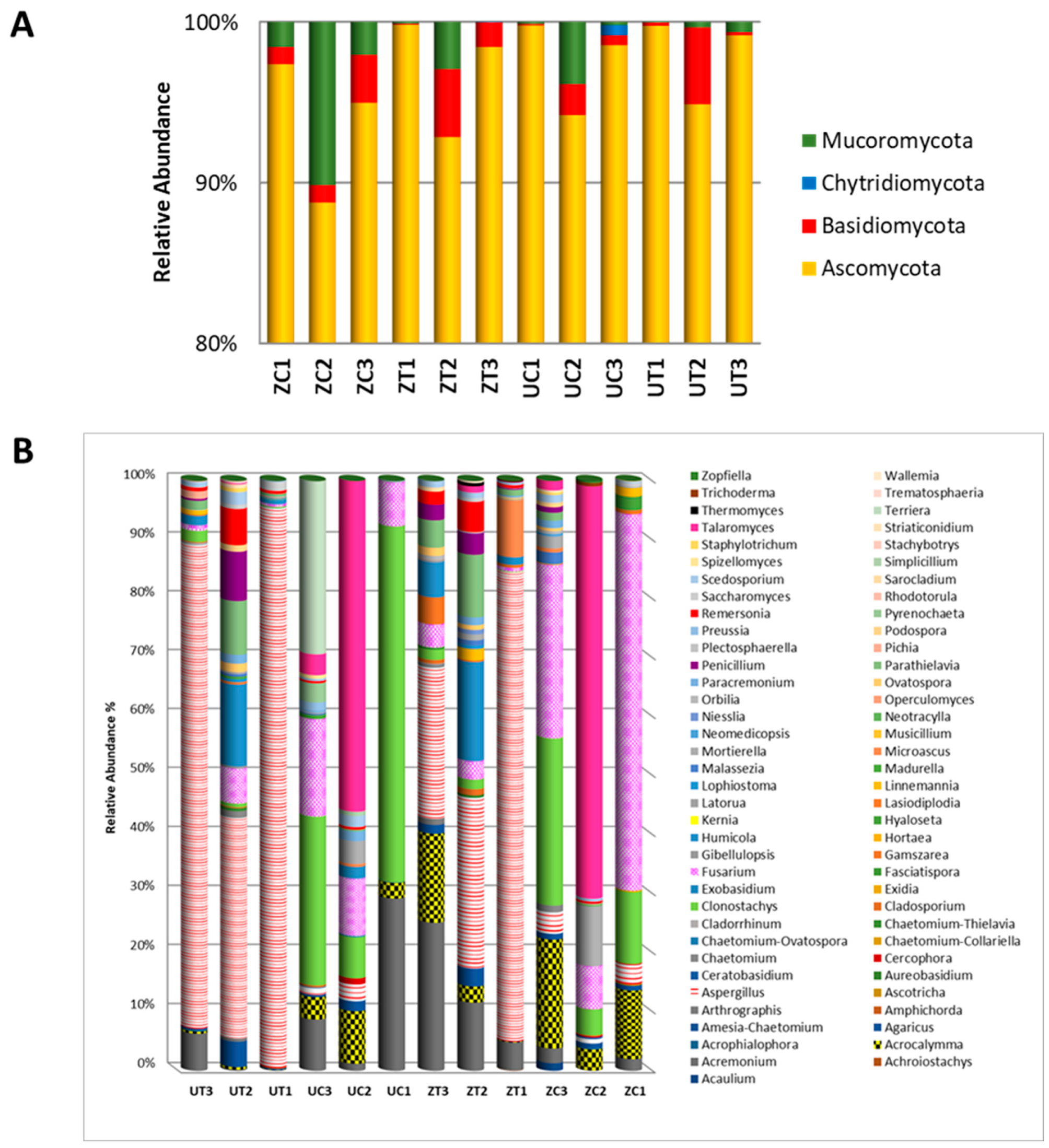
3.4. Epiphytic Fungal Diversity in Response to Plant Genotype and Salinity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Valenzuela, F.J.; Reineke, D.; Leventini, D.; Chen, C.C.L.; Barrett-Lennard, E.G.; Colmer, T.D.; Dodd, I.C.; Shabala, S.; Brown, P.; Bazihizina, N. Plant responses to heterogeneous salinity: Agronomic relevance and research priorities. Ann. Bot. 2022, 129, 499–518. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Ahmad, P. Role of mineral nutrients in abiotic stress tolerance: Revisiting the associated signaling mechanisms. Plant Signal. Mol. 2019, 269–285. [Google Scholar] [CrossRef]
- Al-Hadidi, S.H.; Abumaali, D.A.; Ahmed, T.; Al-khis, A.F.; Al-Malki, S.A.; Yaish, M.; Rahim, H.U.; Khalid, M.F.; Hassan, H.; Alatalo, J.M. The effect of type and combination of fertilizers on eukaryotic microbiome of date palm rhizosphere. Plant Growth Regul. 2024, 1–13. [Google Scholar] [CrossRef]
- Baldrian, P.; López-Mondéjar, R.; Kohout, P. Forest microbiome and global change. Nat. Rev. Microbiol. 2023, 21, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, G.; Dakheel, A.; Brown, G.; Rillig, M. Potential of arbuscular mycorrhizal technology in date palm production. Date Palm Biotechnol. 2011, 449–476. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Baslam, M.; Ben-Laouane, R.; Anli, M.; Boutasknit, A.; Mitsui, T.; Wahbi, S.; Meddich, A. Alleviation of detrimental effects of salt stress on date palm (Phoenix dactylifera L.) by the application of arbuscular mycorrhizal fungi and/or compost. Front. Sustain. Food Syst. 2020, 4, 131. [Google Scholar] [CrossRef]
- Anli, M.; Kaoua, M.E.; Boutasknit, A.; Ben-Laouane, R.; Toubali, S.; Baslam, M.; Lyamlouli, K.; Hafidi, M.; Meddich, A. Seaweed extract application and arbuscular mycorrhizal fungal inoculation: A tool for promoting growth and development of date palm (Phoenix dactylifera L.) cv «Boufgous». S. Afr. J. Bot. 2020, 132, 15–21. [Google Scholar] [CrossRef]
- Adedayo, A.A.; Babalola, O.O. Fungi That Promote Plant Growth in the Rhizosphere Boost Crop Growth. J. Fungi 2023, 9, 239. [Google Scholar] [CrossRef]
- Jaiti, F.; Meddich, A.; El Hadrami, I. Effectiveness of arbuscular mycorrhizal fungi in the protection of date palm (Phoenix dactylifera L.) against bayoud disease. Physiol. Mol. Plant Pathol. 2007, 71, 166–173. [Google Scholar] [CrossRef]
- Abohatem, M.; Chakrafi, F.; Jaiti, F.; Dihazi, A.; Baaziz, M. Arbuscular mycorrhizal fungi limit incidence of Fusarium oxysporum f. sp. albedinis on date palm seedlings by increasing nutrient contents, total phenols and peroxidase activities. Open Hortic. J. 2011, 4, 10–16. [Google Scholar] [CrossRef]
- Al-Karaki, G.N. Application of mycorrhizae in sustainable date palm cultivation. Emir. J. Food Agric. 2013, 25, 854–862. [Google Scholar] [CrossRef]
- Qaddoury, A. Arbuscular mycorrhizal fungi provide complementary characteristics that improve plant tolerance to drought and salinity: Date palm as model. In Mycoremediation and Environmental Sustainability; Springer: Cham, Switzerland, 2017; Volume 1, pp. 189–215. [Google Scholar] [CrossRef]
- Ghadbane, M.; Medjekal, S.; Benderradji, L.; Belhadj, H.; Daoud, H. Assessment of arbuscular mycorrhizal fungi status and rhizobium on date palm (Phoenix dactylifera L.) cultivated in a Pb contaminated soil. In Proceedings of the Recent Advances in Environmental Science from the Euro-Mediterranean and Surrounding Regions (2nd Edition), Proceedings of 2nd Euro-Mediterranean Conference for Environmental Integration (EMCEI-2), Sousse, Tunisia, 10–13 October 2019; Springer: Berlin/Heidelberg, Germany, 2021; pp. 703–707. [Google Scholar] [CrossRef]
- Robinson, C.H.; Wookey, P.A.; Parker, T.C. Root-associated fungi and carbon storage in Arctic ecosystems. N. Phytol. 2020, 226, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Arunrat, N.; Sansupa, C.; Sereenonchai, S.; Hatano, R. Short-term response of soil bacterial and fungal communities to fire in rotational shifting cultivation, northern Thailand. Appl. Soil Ecol. 2024, 196, 105303. [Google Scholar] [CrossRef]
- Szili-Kovács, T.; Takács, T. Advanced Research of Rhizosphere Microbial Activity. Agriculture 2023, 13, 911. [Google Scholar] [CrossRef]
- Yaish, M.W.; Kumar, P. Salt Tolerance Research in Date Palm Tree (Phoenix dactylifera L.), past, present, and future perspectives. Front. Plant Sci. 2015, 6, 348. [Google Scholar] [CrossRef]
- Jana, G.A.; Yaish, M.W. Isolation and functional characterization of a mVOC producing plant-growth-promoting bacterium isolated from the date palm rhizosphere. Rhizosphere 2020, 16, 100267. [Google Scholar] [CrossRef]
- Yaish, M.W.; Al-Harrasi, I.; Alansari, A.S.; Al-Yahyai, R.; Glick, B.R. The use of high throughput DNA sequence analysis to assess the endophytic microbiome of date palm roots grown under different levels of salt stress. Int. Microbiol. 2016, 19, 143–155. [Google Scholar]
- Al-Busaidi, A.; Glick, B.R.; Yaish, M.W. The Effect of Date Palm Genotypes on Rhizobacterial Community Structures under Saline Environments. Biology 2022, 11, 1666. [Google Scholar] [CrossRef]
- Gołębiewski, M.; Tretyn, A. Generating amplicon reads for microbial community assessment with next-generation sequencing. J. Appl. Microbiol. 2020, 128, 330–354. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Caraux, G.; Pinloche, S. PermutMatrix: A graphical environment to arrange gene expression profiles in optimal linear order. Bioinformatics 2005, 21, 1280–1281. [Google Scholar] [CrossRef] [PubMed]
- IBM. IBM SPSS Statistics for Windows; IBM: Armonk, NY, USA, 2012. [Google Scholar]
- Jana, G.A.; Glick, B.R.; Yaish, M.W. Salt tolerance in plants: Using OMICS to assess the impact of plant growth-promoting bacteria (PGPB). In Mitigation of Plant Abiotic Stress by Microorganisms; Academic Press: Cambridge, MA, USA, 2022; pp. 299–320. [Google Scholar]
- Al Kharusi, L.; Jana, G.A.; Patankar, H.V.; Yaish, M.W. Comparative Metabolic Profiling of Two Contrasting Date Palm Genotypes Under Salinity. Plant Mol. Biol. Report. 2021, 39, 351–363. [Google Scholar] [CrossRef]
- Vega-Gutiérrez, T.; Tirado-Ramírez, M.; López-Urquídez, G.; Angulo-Castro, A.; Martínez-Gallardo, J.; López-Orona, C. Fusarium falciforme (FSSC 3 + 4) causing root and stem rot in papaya (Carica papaya) in Mexico. Plant Dis. 2019, 103, 2681. [Google Scholar] [CrossRef]
- Sousa, E.; Melo, M.; Mota, J.; Sousa, E.; Beserra, J., Jr.; Matos, K. First report of Fusarium falciforme (FSSC 3 + 4) causing root rot in lima bean (Phaseolus lunatus L.) in Brazil. Plant Dis. 2017, 101, 1954. [Google Scholar] [CrossRef]
- Wang, R.-Y.; Gao, B.; Li, X.-H.; Ma, J.; Chen, S.-L. First report of Fusarium solani causing Fusarium root rot and stem canker on storage roots of sweet potato in China. Plant Dis. 2014, 98, 160. [Google Scholar] [CrossRef]
- Kyekyeku, J.O.; Kusari, S.; Adosraku, R.K.; Bullach, A.; Golz, C.; Strohmann, C.; Spiteller, M. Antibacterial secondary metabolites from an endophytic fungus, Fusarium solani JK10. Fitoterapia 2017, 119, 108–114. [Google Scholar] [CrossRef]
- Kavroulakis, N.; Doupis, G.; Papadakis, I.E.; Ehaliotis, C.; Papadopoulou, K.K. Tolerance of tomato plants to water stress is improved by the root endophyte Fusarium solani FsK. Rhizosphere 2018, 6, 77–85. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, W.; Perera, R.H.; Lim, Y.W. Long-Term Investigation of Marine-Derived Aspergillus Diversity in the Republic of Korea. Mycobiology 2023, 51, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Gómez, I.; Valdés-Muñoz, G.; Moreno-Ulloa, A.; Pérez-Llano, Y.; Moreno-Perlín, T.; Silva-Jiménez, H.; Barreto-Curiel, F.; Sánchez-Carbente, M.d.R.; Folch-Mallol, J.L.; Gunde-Cimerman, N. Surviving in the brine: A multi-omics approach for understanding the physiology of the halophile fungus Aspergillus sydowii at saturated NaCl concentration. Front. Microbiol. 2022, 13, 840408. [Google Scholar] [CrossRef] [PubMed]
- Baron, N.C.; Costa, N.T.A.; Mochi, D.A.; Rigobelo, E.C. First report of Aspergillus sydowii and Aspergillus brasiliensis as phosphorus solubilizers in maize. Ann. Microbiol. 2018, 68, 863–870. [Google Scholar] [CrossRef]
- Escobar Diaz, P.A.; Gil, O.J.A.; Barbosa, C.H.; Desoignies, N.; Rigobelo, E.C. Aspergillus spp. and Bacillus spp. as growth promoters in cotton plants under greenhouse conditions. Front. Sustain. Food Syst. 2021, 5, 709267. [Google Scholar] [CrossRef]
- Vala, A.K.; Dave, B.P.; Dube, H.C. Chemical characterization and quantification of siderophores produced by marine and terrestrial aspergilli. Can. J. Microbiol. 2006, 52, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llano, Y.; Rodríguez-Pupo, E.C.; Druzhinina, I.S.; Chenthamara, K.; Cai, F.; Gunde-Cimerman, N.; Zalar, P.; Gostinčar, C.; Kostanjšek, R.; Folch-Mallol, J.L. Stress reshapes the physiological response of halophile fungi to salinity. Cells 2020, 9, 525. [Google Scholar] [CrossRef] [PubMed]
- Pauvert, C.; Buée, M.; Laval, V.; Edel-Hermann, V.; Fauchery, L.; Gautier, A.; Lesur, I.; Vallance, J.; Vacher, C. Bioinformatics matters: The accuracy of plant and soil fungal community data is highly dependent on the metabarcoding pipeline. Fungal Ecol. 2019, 41, 23–33. [Google Scholar] [CrossRef]
- Mefteh, F.B.; Daoud, A.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Rateb, M.E.; Kadri, A.; Gharsallah, N.; Belbahri, L. Fungal root microbiome from healthy and brittle leaf diseased date palm trees (Phoenix dactylifera L.) reveals a hidden untapped arsenal of antibacterial and broad spectrum antifungal secondary metabolites. Front. Microbiol. 2017, 8, 307. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Aoki, T.; Ariyawansa, H.A.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Fungal taxonomy and sequence-based nomenclature. Nat. Microbiol. 2021, 6, 540–548. [Google Scholar] [CrossRef]
- Ya, T.; Du, S.; Li, Z.; Liu, S.; Zhu, M.; Liu, X.; Jing, Z.; Hai, R.; Wang, X. Successional dynamics of molecular ecological network of anammox microbial communities under elevated salinity. Water Res. 2021, 188, 116540. [Google Scholar] [CrossRef]
- Chandran, S.; Sanka Loganathachetti, D.; Masmoudi, K.; Iratni, R.; Mundra, S. Irrigation water source matters: Saline groundwater irrigation lowers date palm root-associated fungal richness and alters their community structural patterns. Phytobiomes J. 2023, 8, 59–69. [Google Scholar] [CrossRef]
- Ge, Y.; Yu, F.; Tan, Y.; Zhang, X.; Liu, Z. Comparative transcriptome sequence analysis of sporulation-related genes of Aspergillus cristatus in response to low and high osmolarity. Curr. Microbiol. 2017, 74, 806–814. [Google Scholar] [CrossRef]
- Chebaane, A.; Symanczik, S.; Oehl, F.; Azri, R.; Gargouri, M.; Mäder, P.; Mliki, A.; Fki, L. Arbuscular mycorrhizal fungi associated with Phoenix dactylifera L. grown in Tunisian Sahara oases of different salinity levels. Symbiosis 2020, 81, 173–186. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Number of Reads | ||||||
|---|---|---|---|---|---|---|
| R1 | R2 | R3 | Average | Comparison | p-Value | |
| ZC | 11,297 | 18,693 | 17,903 | 15,964 | ZC vs. ZT | 0.1338 |
| ZT | 25,098 | 28,667 | 36,091 | 29,952 | UC vs. UT | 1.0 |
| UC | 27,727 | 23,607 | 13,800 | 21,711 | ZT vs. UT | 0.5187 |
| UT | 31,704 | 21,278 | 12,964 | 21,982 | ZC vs. UC | 0.7385 |
| OTU Enrichment | ||||||
| R1 | R2 | R3 | Average | Pairwise Comparison | p-Value | |
| ZC | 20 | 21 | 34 | 25 | ZC vs. ZT | 0.37 |
| ZT | 33 | 42 | 31 | 35 | UC vs. UT | 0.30 |
| UC | 9 | 25 | 24 | 19 | ZT vs. UT | 0.86 |
| UT | 27 | 38 | 27 | 31 | ZC vs. UC | 0.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaish, M.W.; Al-Busaidi, A.; Glick, B.R.; Ahmed, T.; Alatalo, J.M. The Effects of Salinity and Genotype on the Rhizospheric Mycobiomes in Date Palm Seedlings. Biology 2024, 13, 190. https://doi.org/10.3390/biology13030190
Yaish MW, Al-Busaidi A, Glick BR, Ahmed T, Alatalo JM. The Effects of Salinity and Genotype on the Rhizospheric Mycobiomes in Date Palm Seedlings. Biology. 2024; 13(3):190. https://doi.org/10.3390/biology13030190
Chicago/Turabian StyleYaish, Mahmoud W., Aya Al-Busaidi, Bernard R. Glick, Talaat Ahmed, and Juha M. Alatalo. 2024. "The Effects of Salinity and Genotype on the Rhizospheric Mycobiomes in Date Palm Seedlings" Biology 13, no. 3: 190. https://doi.org/10.3390/biology13030190
APA StyleYaish, M. W., Al-Busaidi, A., Glick, B. R., Ahmed, T., & Alatalo, J. M. (2024). The Effects of Salinity and Genotype on the Rhizospheric Mycobiomes in Date Palm Seedlings. Biology, 13(3), 190. https://doi.org/10.3390/biology13030190