
Exploring the Climatic Niche Evolution of the Genus Falco (Aves: Falconidae) in Europe

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Occurrence Points, Climatic Data, and Species Distribution Modeling
2.2. Niche Overlap and Niche Breadth Estimation
2.3. Phylogeny and Diversification
2.4. Niche Evolution Analyses
2.5. Age Range Correlation
3. Results
3.1. Environmental Niche Modeling: Niche Analysis
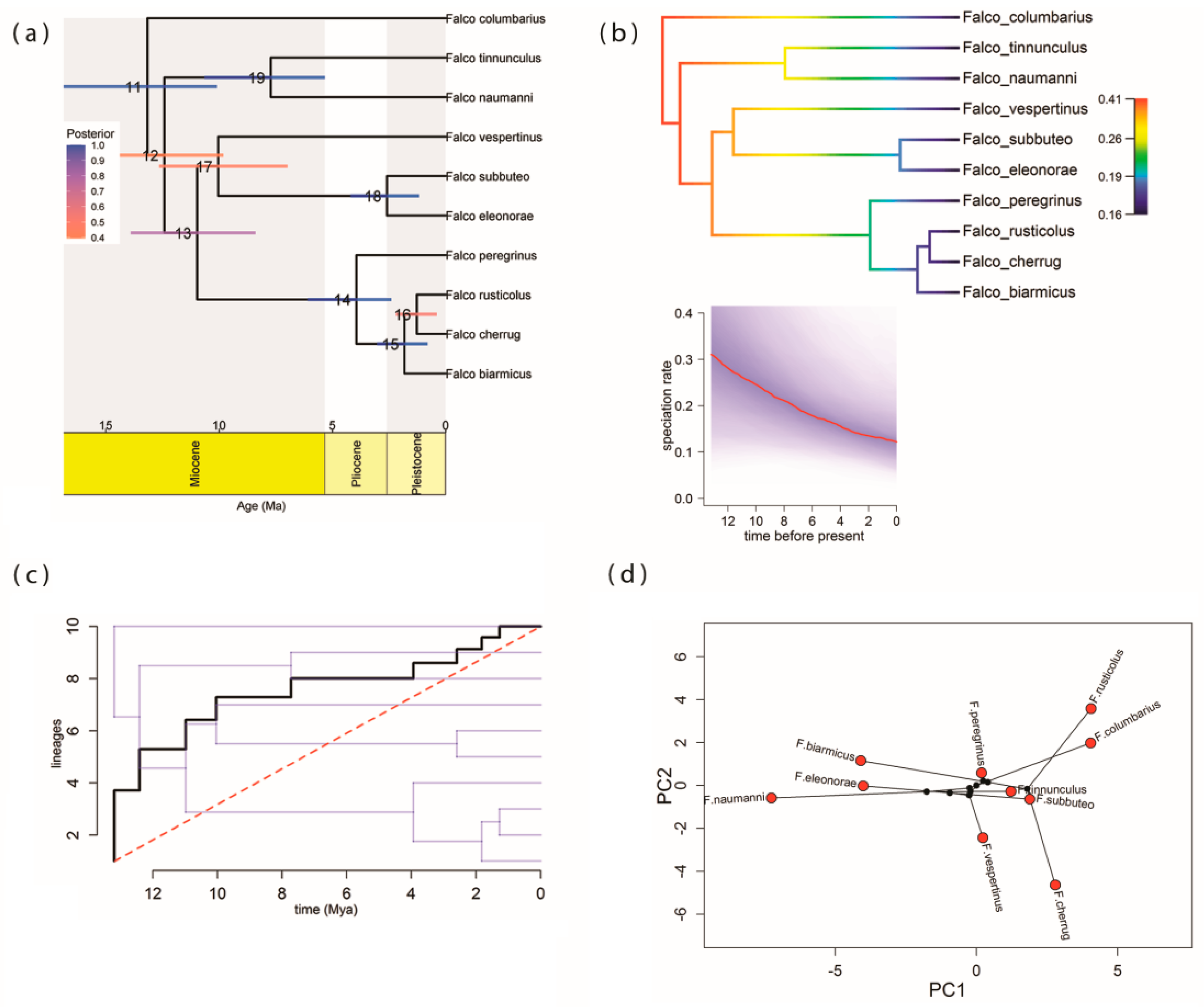
3.2. Phylogenetic Relationships and Divergence Time Estimates
3.3. Pattern of Lineage Diversification
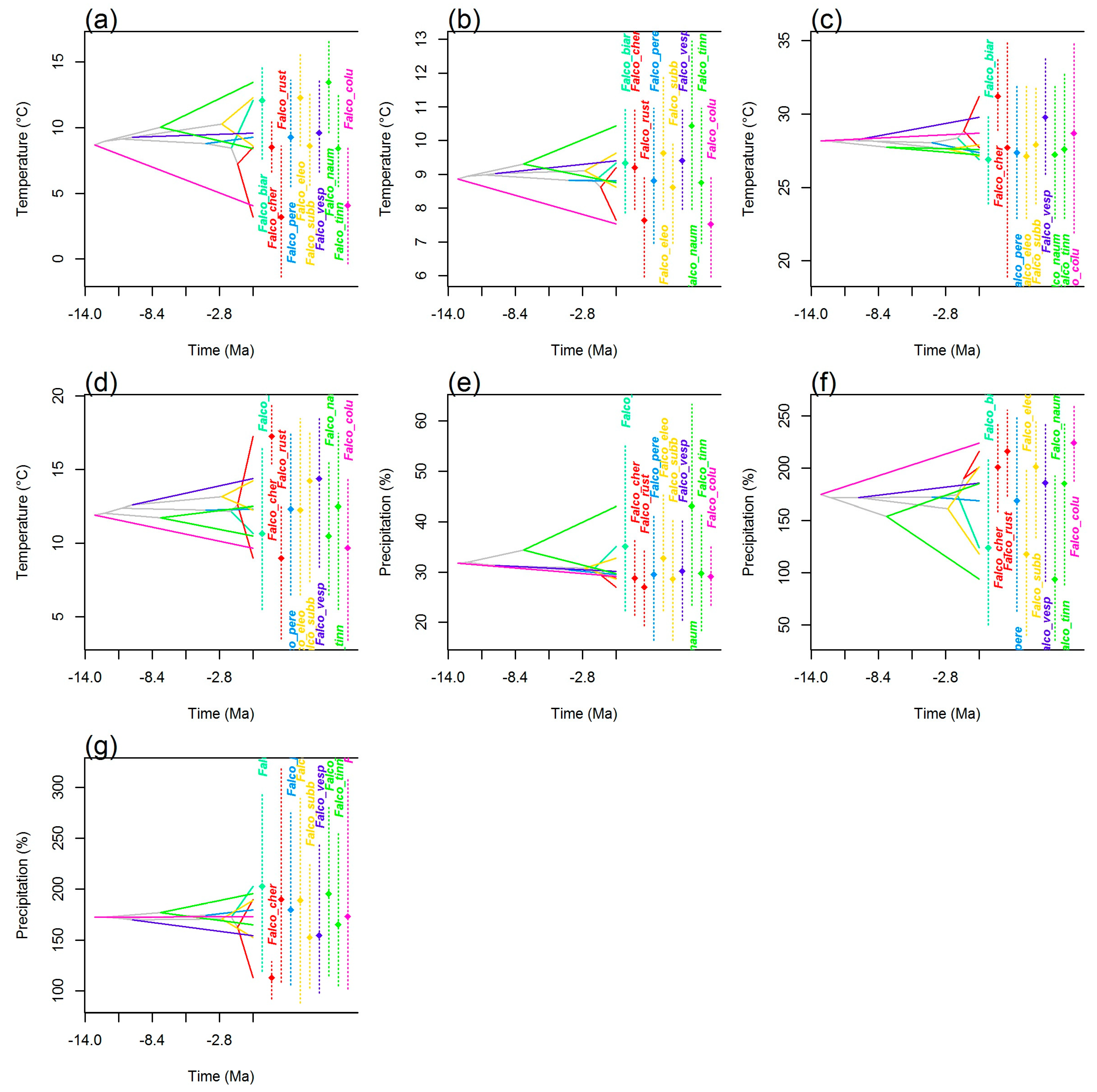
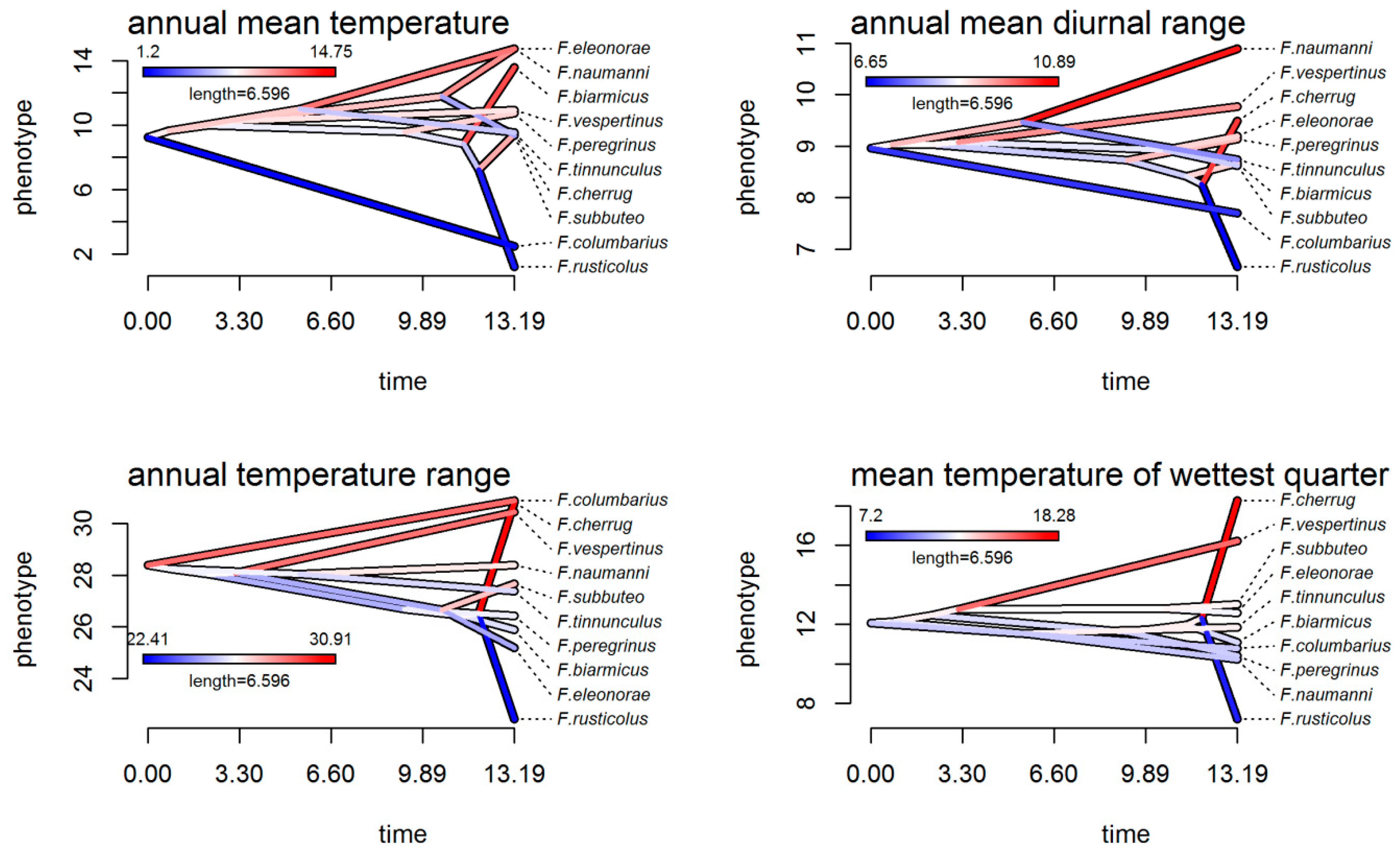
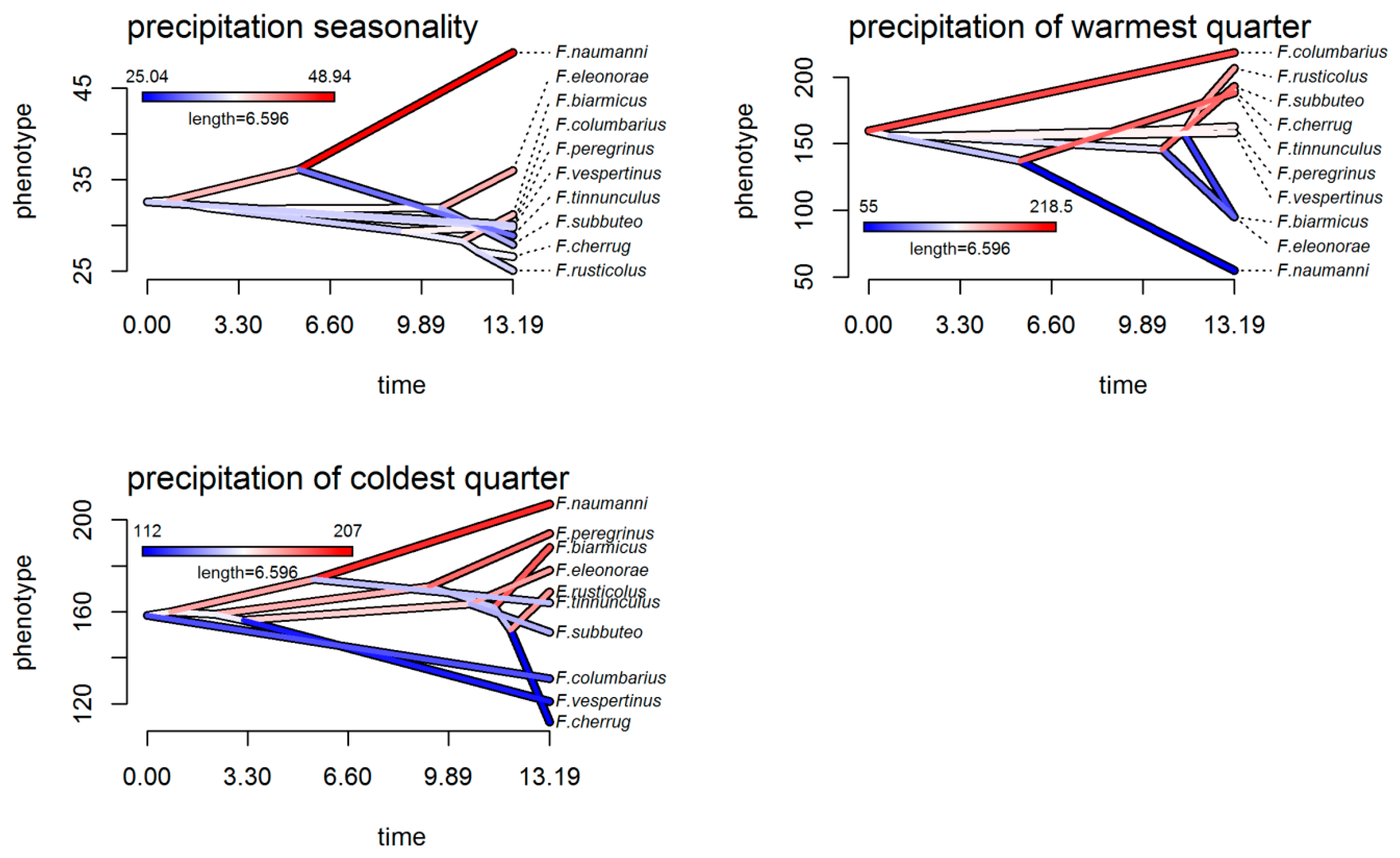
3.4. Climatic Niche Evolution
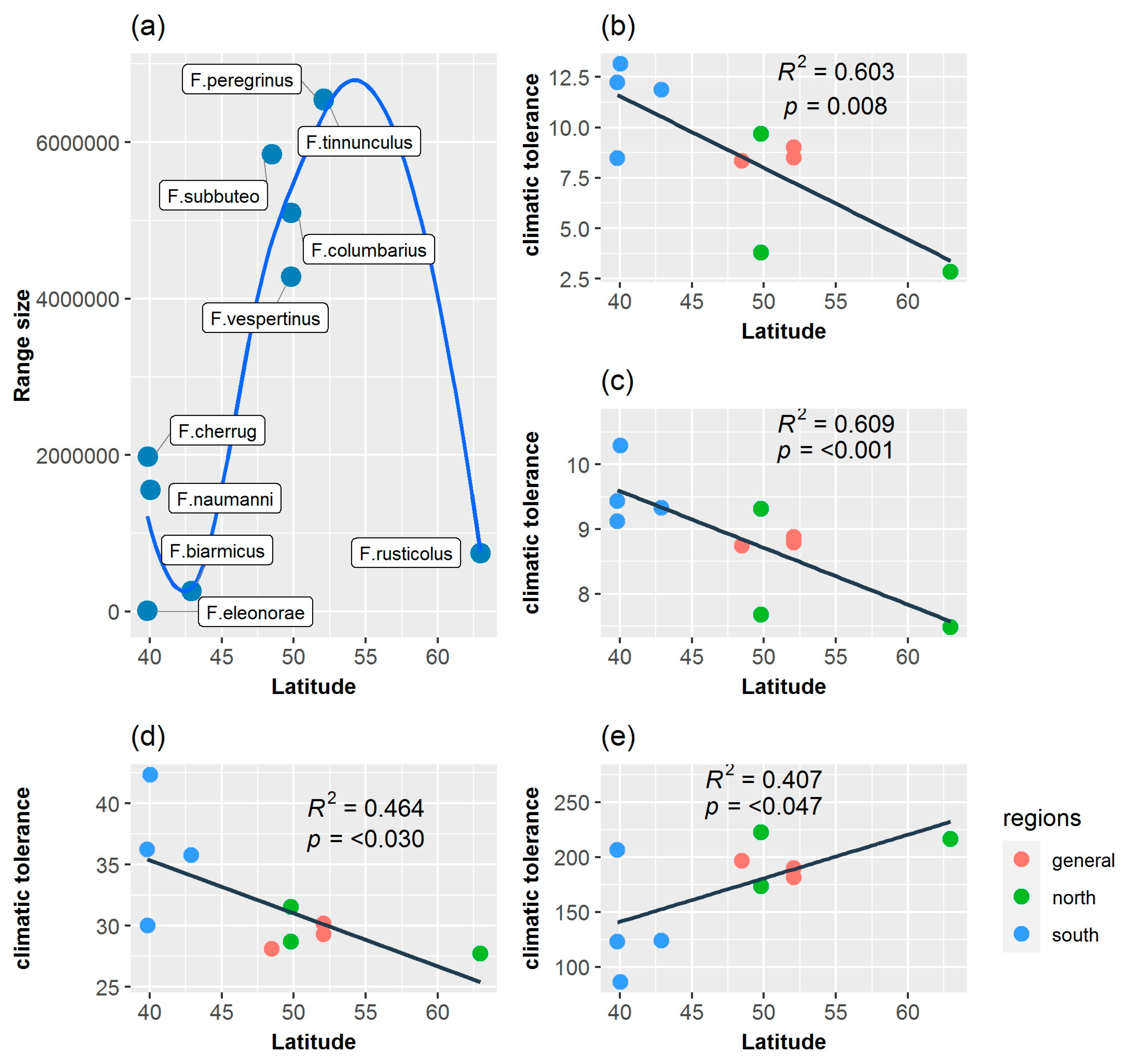
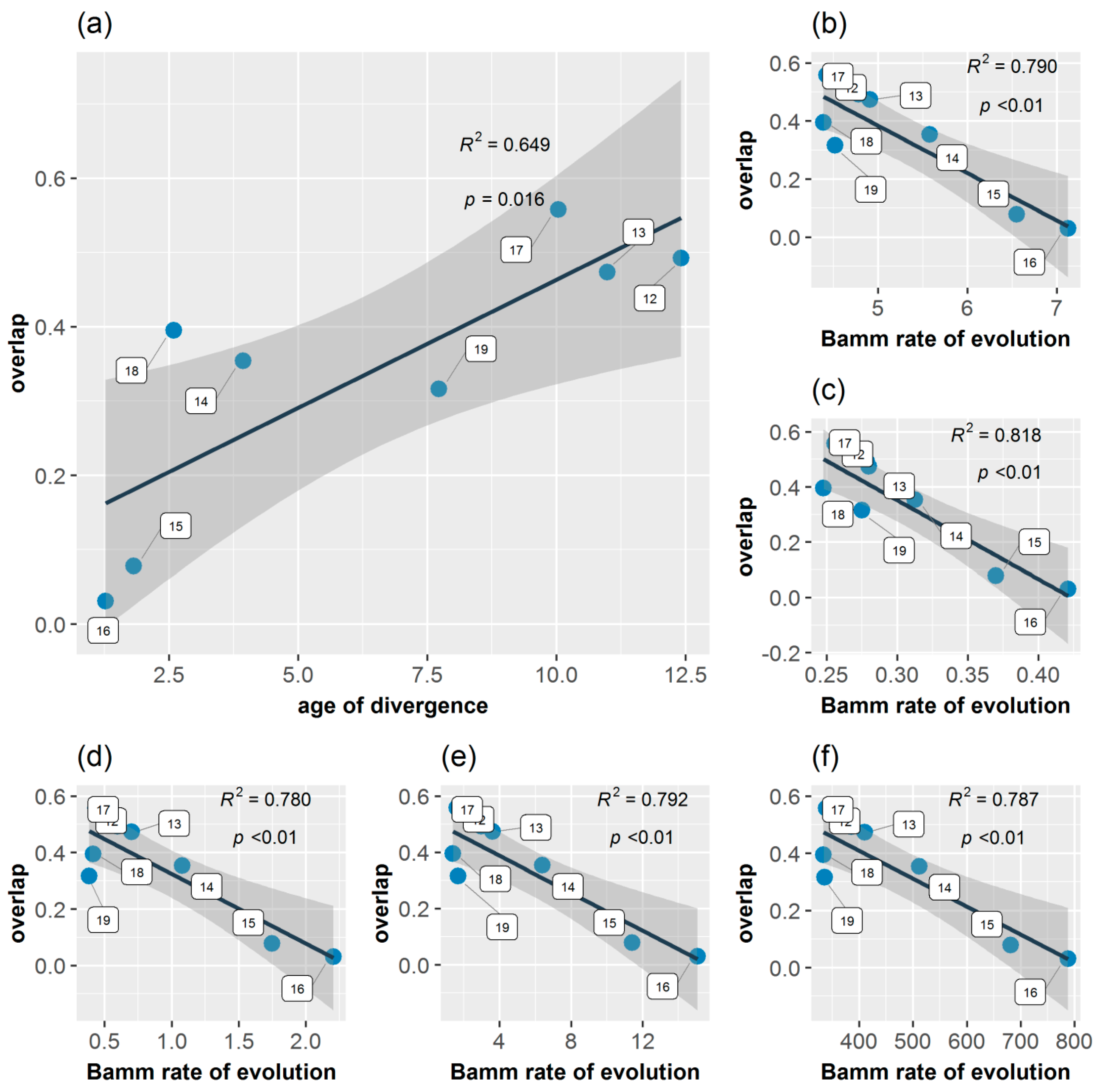
3.5. Age Range Correlation
4. Discussion
4.1. Diversification and Phylogeny
4.2. Niche Evolution
4.2.1. Niche Conservatism
4.2.2. Convergent Evolution
4.2.3. Niche Divergence
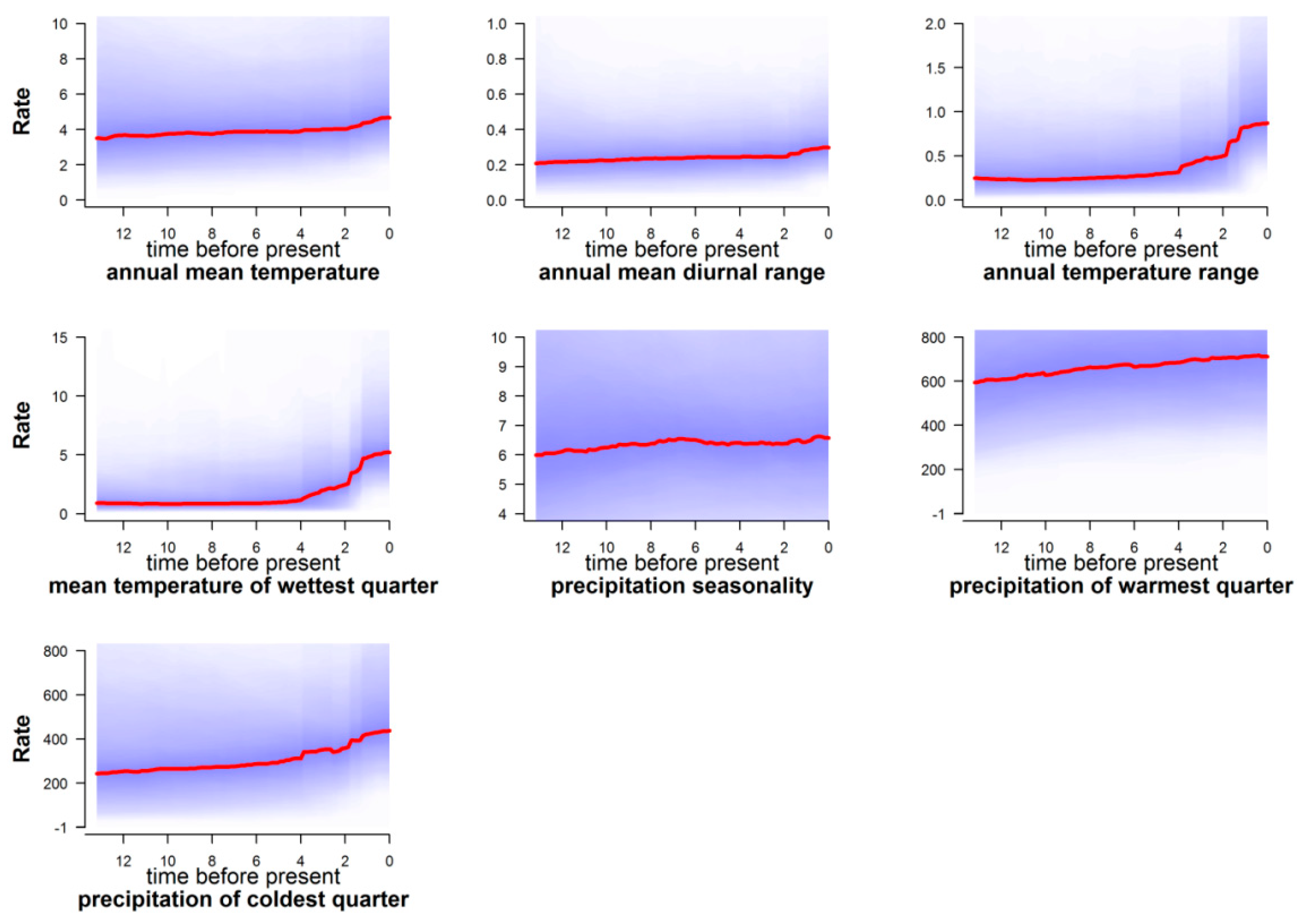
4.2.4. Rate of Climatic Niche Evolution and Climatic Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fisher-Reid, M.C.; Kozak, K.H.; Wiens, J.J. How is the rate of climatic-niche evolution related to climatic-niche breadth? Evolution 2012, 66, 3836–3851. [Google Scholar] [CrossRef]
- Quintero, I.; Suchard, M.A.; Walter, J. Macroevolutionary dynamics of climatic niche space. Proc. R. Soc. B 2022, 289, 20220091. [Google Scholar] [CrossRef]
- Wiens, J.J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Jonathan Davies, T.; Grytnes, J.-A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef]
- Duran, A.; Pie, M.R. Tempo and mode of climatic niche evolution in Primates. Evolution 2015, 69, 2496–2506. [Google Scholar] [CrossRef]
- Portillo, J.T.M.; Barbo, F.E.; Sawaya, R.J. Climatic niche breadths of the Atlantic Forest snakes do not increase with increasing latitude. Curr. Zool. 2022, 68, 535–540. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.G. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Huntley, B.; Gree, R.E.; Collingham, Y.C.; Willis, S.G. A Climatic Atlas of European Breeding Birds; The RSPB and Lynx Edicions; Durham University: Barcelona, Spain, 2007. [Google Scholar]
- Tingley, M.W.; Monahan, W.B.; Beissinger, S.R.; Moritz, C. Birds track their Grinnellian niche through a century of climate change. Proc. Natl. Acad. Sci. USA 2009, 106 (Suppl. S2), 19637–19643. [Google Scholar] [CrossRef]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating Evolution, Ecology, and Conservation Biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Wiens, J.J.; Donoghue, M.J. Historical biogeography, ecology and species richness. Trends Ecol. Evol. 2004, 19, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, S.; Evans, M.E.K.; Burfield, I.J.; Jiguet, F.; Thuiller, W. Are species’ responses to global change predicted by past niche evolution? Philos. Trans. R. Soc. B 2013, 368, 20120091. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; Hemami, M.R.; Kaboli, M.; Nazarizadeh, M.; Malekian, M.; Behrooz, R.; Geniez, P.; Alroy, J.; Zimmermann, N.E. The legacy of Eastern Mediterranean mountain uplifts: Rapid disparity of phylogenetic niche conservatism and divergence in mountain vipers. BMC Ecol. Evol. 2021, 21, 130. [Google Scholar] [CrossRef] [PubMed]
- Jezkova, T.; Wiens, J.J. Rates of change in climatic niches in plant and animal populations are much slower than projected climate change. Proc. R. Soc. B 2016, 283, 20162104. [Google Scholar] [CrossRef]
- Meyer, A.L.S.; Pie, M.R. Climate Change Estimates Surpass Rates of Climatic Niche Evolution in Primates. Int. J. Primatol. 2022, 43, 40–56. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Leavitt, S.D.; Heller, K.; Muggia, L.; Lumbsch, H.T. Contrasting patterns of climatic niche divergence in Trebouxia-A clade of lichen-forming algae. Front. Microbiol. 2022, 13, 791546. [Google Scholar] [CrossRef]
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Nosil, P. Ecological Speciation; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Cruz, C.; Santulli-Sanzo, G.; Ceballos, G. Global patterns of raptor distribution and protected areas optimal selection to reduce the extinction crises. Proc. Natl. Acad. Sci. USA 2021, 118, e2018203118. [Google Scholar] [CrossRef]
- Fuchs, J.; Johnson, J.A.; Mindell, D.P. Rapid diversification of falcons (Aves: Falconidae) due to expansion of open habitats in the Late Miocene. Mol. Phylogenetics Evol. 2015, 82, 166–182. [Google Scholar] [CrossRef]
- Wilcox, J.J.S.; Boissinot, S.; Idaghdour, Y. Falcon genomics in the context of conservation, speciation, and human culture. Ecol. Evol. 2019, 9, 14523–14537. [Google Scholar] [CrossRef]
- del Hoyo, J. All the Birds of the World; Lynx Edicions: Barcelona, Spain, 2020. [Google Scholar]
- del Hoyo, J.; Elliott, A.; Sargatal, J. (Eds.) Handbook of the Birds of the World. Volume 2. New World Vultures to Guineafowl; Lynx Edicions: Barcelona, Spain, 1994. [Google Scholar]
- Keller, V.; Herrando, S.; Voříšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Martí, D.; Anton, M.; Klvaňová, A.; Kalyakin, M.V.; et al. European Breeding Bird Atlas 2: Distribution, Abundance and Change; European Bird Census Council and Lynx Edicions: Barcelona, Spain, 2020. [Google Scholar]
- Nittinger, F.; Gamauf, A.; Pinsker, W.; Wink, M.; Haring, E. Phylogeography and population structure of the saker falcon (Falco cherrug) and the influence of hybridization: Mitochondrial and microsatellite data. Mol. Ecol. 2007, 16, 1497–1517. [Google Scholar] [CrossRef]
- Wink, M. Phylogeny of falconidae and phylogeography of peregrine falcons. Ornis Hung. 2018, 26, 27–37. [Google Scholar] [CrossRef]
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R.; et al. CoordinateCleaner: Standardized cleaning of occurrence records from biological collection databases. Methods Ecol. Evol. 2019, 10, 744–751. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R (3.6.2); Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 30 October 2021).
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. dismo: Species Distribution Modeling; R Package Version 0.8-17. 2013. Available online: https://rspatial.org/raster/sdm/ (accessed on 8 November 2021).
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Change Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Brown, J.L.; Carnaval, A.C. A tale of two niches: Methods, concepts, and evolution. Front. Biogeogr. 2019, 11, e44158. [Google Scholar] [CrossRef]
- Engler, J.O.; Lawrie, Y.; Cabral, J.S.; Lens, L.; Martínez-Meyer, E. Niche evolution reveals disparate signatures of speciation in the ‘great speciator’ (white-eyes, Aves: Zosterops). J. Biogeogr. 2021, 48, 1981–1993. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.-J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Guisan, A. ecospat: An R package to support spatial analyses and modelling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Glor, R.E.; Huron, N.A.; Simões, M.; Iglesias, T.L.; et al. ENMTools 1.0: An R package for comparative ecological biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- Levins, R. Evolution in Changing Environments: Some Theoretical Explorations; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2009, 33, 607–611. [Google Scholar] [CrossRef]
- Rödder, D.; Engler, J.O. Quantitative metrics of overlaps in Grinnellian niches: Advances and possible drawbacks. Glob. Ecol. Biogeogr. 2011, 20, 915–927. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Pybus, O.G.; Harvey, P.H. Testing macro-evolutionary models using incomplete molecular phylogenies. Proc. R. Soc. B 2000, 267, 2267–2272. [Google Scholar] [CrossRef]
- Condamine, F.L.; Antonelli, A.; Lagomarsino, L.P.; Hoorn, C.; Liow, L.H.; Hsiang, L. Teasing apart mountain uplift, climate change and biotic drivers of species diversification. In Mountains, Climate and Biodiversity; Hoorn, C., Perrigo, A., Antonelli, A., Eds.; Wiley: Hoboken, NJ, USA, 2018; pp. 257–272. [Google Scholar]
- Revell, L.J.; Harmon, L.J. Phylogenetic Comparative Methods in R; Princeton University Press: Princeton, NJ, USA, 2022; 440p. [Google Scholar]
- Etienne, R.S.; Haegeman, B.; Stadler, T.; Aze, T.; Pearson, P.N.; Purvis, A.; Phillimore, A.B. Diversity-dependence brings molecular phylogenies closer to agreement with the fossil record. Proc. Biol. Sci. 2012, 279, 1300–1309. [Google Scholar] [CrossRef] [PubMed]
- FitzJohn, R.G. Diversitree: Comparative Phylogenetic Analyses of Diversification in R. Methods Ecol. Evol. 2012; in press. [Google Scholar] [CrossRef]
- Swenson, N.G. Phylogenetic Ecology: A History, Critique, and Remodeling; The University of Chicago Press: Chicago, IL, USA, 2019. [Google Scholar]
- Cadotte, M.W.; Davies, T.J. Phylogenies in Ecology: A Guide to Concepts and Methods; Princeton University Press: Princeton, NJ, USA, 2016. [Google Scholar]
- Keck, F.; Rimet, F.; Bouchez, A.; Franc, A. phylosignal: An R package to measure, test, and explore the phylogenetic signal. Ecol. Evol. 2016, 6, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Heibl, C.; Calenge, C. phyloclim: Integrating Phylogenetics and Climatic Niche Modeling; R Package Version 0.9-4. 2013. Available online: http://CRAN.Rproject.org/package=phyloclim (accessed on 10 February 2022).
- Evans, M.E.; Smith, S.A.; Flynn, R.S.; Donoghue, M.J. Climate, niche evolution, and diversification of the “bird-cage” evening primroses (Oenothera, sections Anogra and Kleinia). Am. Nat. 2009, 173, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Clavel, J.; Escarguel, G.; Merceron, G. mvMORPH: An R package for fitting multivariate evolutionary models to morphometric data. Methods Ecol. Evol. 2015, 6, 1311–1319. [Google Scholar] [CrossRef]
- Finlayson, C. Avian Survivors. The History and Biogeography of Palearctic Birds; Tand A D Poyser: London, UK, 2011. [Google Scholar]
- Felsenstein, J. Phylogenies and the comparative method. Am. Nat. 1985, 125, 1. [Google Scholar] [CrossRef]
- O’Meara, B.C.; Ane, C.; Sanderson, M.J.; Wainwright, P.C. Testing for different rates of continuous trait evolution. Evolution 2006, 60, 922–933. [Google Scholar] [PubMed]
- Revell, L.J.; Collar, D.C. Phylogenetic analysis of the evolutionary correlation using likelihood. Evolution 2009, 63, 1090–1100. [Google Scholar] [CrossRef]
- Hansen, T.F. Stabilizing selection and the comparative analysis of adaptation. Evolution 1997, 51, 1341–1351. [Google Scholar] [CrossRef]
- Butler, M.A.; King, A.A. Phylogenetic comparative analysis: Amodeling approach for adaptive evolution. Am. Nat. 2004, 164, 683–695. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Jhwueng, D.C.; Boettiger, C.; O’Meara, B.C. Modeling stabilizing selection: Expanding the Ornstein–Uhlenbeck model of adaptive evolution. Evolution 2012, 66, 2369–2383. [Google Scholar] [CrossRef]
- Harmon, L.J.; Losos, J.B.; Jonathan Davies, T.; Gillespie, R.G.; Gittleman, J.L.; Bryan Jennings, W.; Kozak, K.H.; McPeek, M.A.; Moreno-Roark, F.; Near, T.J.; et al. Early bursts of body size and shape evolution are rare in comparative data. Evolution 2010, 64, 2385–2396. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Nielsen, R.; Bollback, J.P. Stochastic mapping of morphological characters. Syst. Biol. 2003, 52, 131–158. [Google Scholar] [CrossRef]
- Furze, M.E.; Wainwright, D.K.; Huggett, B.A.; Knipfer, T.; McElrone, A.J.; Brodersen, C.R. Ecologically driven selection of nonstructural carbohydrate storage in oak trees. New Phytol. 2021, 232, 567–578. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Martínez-Méndez, N.; Mejía, O.; Ortega, J.; Méndez-de la Cruz, F. Climatic niche evolution in the viviparous Sceloporus torquatus group (Squamata: Phrynosomatidae). PeerJ 2019, 6, e6192. [Google Scholar] [CrossRef]
- Stevens, G.C. The Latitudinal Gradient in Geographical Range: How so Many Species Coexist in the Tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 20 November 2021).
- Vilela, B.; Villalobos, F. letsR: A new R package for data handling and analysis in macroecology. Methods Ecol. Evol. 2015, 6, 1229–1234. [Google Scholar] [CrossRef]
- Arbuckle, K.; Minter, A. windex: Analyzing convergent evolution using the Wheatsheaf index in R. Evol. Bioinform. 2015, 11, 11–14. [Google Scholar] [CrossRef]
- Rabosky, D.; Santini, F.; Eastman, J.; Smith, S.A.; Sidlauskas, B.; Chang, J.; Alfaro, M.E. Rates of speciation and morphological evolution are correlated across the largest vertebrate radiation. Nat. Commun. 2013, 4, 1958. [Google Scholar] [CrossRef]
- Rabosky, D.; Grundler, M.; Anderson, C.; Title, P.; Shi, J.; Brown, J.; Huang, H.; Larson, J. BAMMtools: An R package for the analysis of evolutionary dynamics on phylogenetic trees. Methods Ecol. Evol. 2014, 5, 701–707. [Google Scholar] [CrossRef]
- Fitzpatrick, B.M.; Turelli, M. The geography of mammalian speciation: Mixed signals from phylogenies and range maps. Evolution 2006, 60, 601–615. [Google Scholar]
- García-Navas, V.; Westerman, M. Niche conservatism and phylogenetic clustering in a tribe of arid-adapted marsupial mice, the Sminthopsini. J. Evol. Biol. 2018, 31, 1204–1215. [Google Scholar] [CrossRef]
- García-Navas, V.; Rodríguez-Rey, M.; Christidis, L. Ecological opportunity and ecomorphological convergence in Australasian robins (Petroicidae). J. Avian. Biol. 2017, 49, jav-01552. [Google Scholar] [CrossRef]
- Molina-Venegas, R.; Lima, H. Should we be concerned about incomplete taxon sampling when assessing the evolutionary history of regional biotas? J. Biogeogr. 2021, 48, 2387–2390. [Google Scholar] [CrossRef]
- Rosenberg, M.S.; Kumar, S. Incomplete taxon sampling is not a problem for phylogenetic inference. Proc. Natl. Acad. Sci. USA 2001, 98, 10751–10756. [Google Scholar] [CrossRef]
- Rosindell, J.; Harmon, L.J. OneZoom: A fractal explorer for the tree of life. PLoS Biol. 2012, 10, e1001406. [Google Scholar] [CrossRef]
- Blondel, J.; Mourer-Chauviré, C. Evolution and history of the western Palaearctic avifauna. Trends Ecol. Evol. 1998, 13, 488–492. [Google Scholar] [CrossRef]
- Avise, J.C.; Walker, D. Pleistocene phylogeographic effects on avian populations and the speciation process. Proc. R. Soc. Lond. B 1998, 265, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecol. Lett. 2008, 11, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Ferri-Yáñez, F.; Bozinovic, F.; Marquet, P.A.; Valladares, F.; Chown, S.L. Heat freezes niche evolution. Ecol. Lett. 2013, 16, 1206–1219. [Google Scholar] [CrossRef]
- Wiens, J.J. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 2004, 58, 193–197. [Google Scholar] [CrossRef]
- Cooper, N.; Jetz, W.; Freckleton, R.P. Phylogenetic comparative approaches for studying niche conservatism. J. Evol. Biol. 2010, 23, 2529–2539. [Google Scholar] [CrossRef]
- Münkemüller, T.; Boucher, F.C.; Thuiller, W.; Lavergne, S. Phylogenetic niche conservatism—Common pitfalls and ways forward. Funct. Ecol. 2015, 29, 627–639. [Google Scholar] [CrossRef]
- Khaliq, I.; Fritz, S.A.; Prinzinger, R.; Pfenninger, M.; Böhning-Gaese, K.; Hof, C. Global variation in thermal physiology of birds and mammals: Evidence for phylogenetic niche conservatism only in the tropics. J. Biogeogr. 2015, 42, 2187–2196. [Google Scholar] [CrossRef]
- Fjeldså, J.; Christidis, L.; Ericson, P.G.P. (Eds.) The Largest Avian Radiation. In The Evolution of Perching Birds, or the Order Passeriformes; Lynx Edicions: Barcelona, Spain, 2020. [Google Scholar]
- Ingram, T.; Mahler, D. SURFACE: Detecting convergent evolution from comparative data by fitting Ornstein-Uhlenbeck models with stepwise Akaike Information Criterion. Methods Ecol. Evol. 2013, 4, 416–425. [Google Scholar] [CrossRef]
- Cooper, N.; Thomas, G.H.; Venditti, C.; Meade, A.; Freckleton, R.P. A cautionary note on the use of Ornstein Uhlenbeck models in macroevolutionary studies. Biol. J. Linn. Soc. 2016, 118, 64–77. [Google Scholar] [CrossRef]
- Gutiérrez-Pesquera, L.M.; Tejedo, M.; Olalla-Tárraga, M.Á.; Duarte, H.; Nicieza, A.; Solé, M. Testing the climate variability hypothesis in thermal tolerance limits of tropical and temperate tadpoles. J. Biogeogr. 2016, 43, 1166–1178. [Google Scholar] [CrossRef]
- Phillimore, A. Geography. Range Evolution, and Speciation. In The Princeton Guide to Evolution; Losos, J.B., Baum, D.A., Futuyma, D.J., Hoeckstra, H.E., Lenski, R.E., Moore, A.J., Peichel, C.L., Schluter, D., Whitlock, M.C., Eds.; Princeton University Press: Princeton, NJ, USA, 2017. [Google Scholar]
- Revell, L.J.; Harmon, L.J.; Collar, D.C. Phylogenetic Signal, Evolutionary Process, and Rate. Syst. Biol. 2008, 57, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.; Shepard, D.B.; Kozak, K.H. Developmental life history is associated with variation in rates of climatic niche evolution in a salamander adaptive radiation. Evolution 2020, 74, 1804–1814. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | logL | λ | μ | AICc | ΔAICc |
|---|---|---|---|---|---|
| Yule | −13.329 | 0.103 | 0 | 28.658 | 0 |
| crBD | −13.330 | 0.105 | <0.001 | 30.660 | 2.002 |
| vS | −12.575 | −0.167 (0.108) | 0.339 | 31.151 | 2.493 |
| vE | −13.329 | 0.103 | 0.081 (<0.001) | 32.658 | 4.000 |
| vSE | −12.549 | −0.209 (0.100) | −0.075 (0.295) | 33.098 | 4.440 |
| DDL + E | −25.114 | 0.262 | 0 | 54.228 | 25.570 |
| DDX + E | −24.365 | 10.248 | 0.070 | 54.731 | 26.073 |
| Trait | Model | Loglik | AICc | ΔAICc | AICc Weight |
|---|---|---|---|---|---|
| Bio1 | BM1 e | −28.402 ± 1.407 | 62.520 ± 2.814 | 1 ± 2.681 | 0.352 |
| BMM ** | −27.355 ± 3.495 | 70.711 ± 6.990 | 9.191 ± 6.857 | 0.004 | |
| OU1 | −25.760 ± 0.067 | 61.520 ± 0.133 | 0 ± 0 | 0.617 | |
| OUM ** | −22.549 ± 2.670 | 70.098 ± 5.341 | 8.578 ± 5.208 | 0.011 | |
| EB ** | −29.265 ± 1.978 | 68.531 ± 3.957 | 7.011 ± 3.824 | 0.016 | |
| Bio2 | BM1 e | −14.232 ± 1.761 | 34.178 ± 0.112 | 0.744 ± 0.183 | 0.445 |
| BMM ** | −12.990 ± 3.532 | 41.980 ± 7.065 | 8.546 ± 6.76 | 0.008 | |
| OU1 | −11. 717 ± 0.152 | 33.434 ± 0.305 | 0 ± 0 | 0.519 | |
| OUM ** | −8.554 ± 2.573 | 42.109 ± 5.146 | 8.675 ± 4.841 | 0.007 | |
| EB * | −15.123 ± 1.841 | 40.246 ± 3.683 | 6.812 ± 3.378 | 0.021 | |
| Bio7 | BM1 * | −18.660 ± 2.247 | 43.035 ± 4.495 | 3.286 ± 4.237 | 0.159 |
| BMM *** | −16.958 ± 4.152 | 49.917 ± 8.305 | 10.168 ± 8.047 | 0.006 | |
| OU1 | −14.874 ± 0.129 | 39.749 ± 0.258 | 0 ± 0 | 0.825 | |
| OUM *** | −13.198 ± 1.781 | 51.396 ± 3.562 | 11.647 ± 3.304 | 0.002 | |
| EB ** | −19.634 ± 2.572 | 49.269 ± 5.144 | 9.52 ± 4.886 | 0.008 | |
| Bio8 | BM1 * | −27.566 ± 2.431 | 60.846 ± 4.862 | 4.43 ± 4.734 | 0.098 |
| BMM *** | −25.993 ± 3.755 | 67.987 ± 7.511 | 11.571 ± 7.383 | 0.002 | |
| OU1 | −23.208 ± 0.064 | 56.416 ± 0.128 | 0 ± 0 | 0.893 | |
| OUM *** | −21.406 ± 1.672 | 67.813 ± 3.344 | 11.397 ± 3.216 | 0.003 | |
| EB *** | −28.670 ± 2.624 | 67.340 ± 5.248 | 10.924 ± 5.12 | 0.004 | |
| Bio15 | BM1 e | −32.143 ± 1.767 | 70.001 ± 3.543 | 1.796 ± 3.324 | 0.277 |
| BMM ** | −30.654 ± 3.086 | 77.308 ± 6.172 | 9.103 ± 5.953 | 0.007 | |
| OU1 | −29.102 ± 0.109 | 68.205 ± 0.219 | 0 ± 0 | 0.679 | |
| OUM ** | −26.022 ± 2.424 | 77.044 ± 4.848 | 8.839 ± 4.629 | 0.008 | |
| EB * | −32.221 ± 1.305 | 74.443 ± 2.611 | 6.238 ± 2.392 | 0.029 | |
| Bio18 | BM1 * | −55.786 ± 1.498 | 116.062 ± 2.99 | 2.345 ± 2.778 | 0.227 |
| BMM ** | −53.178 ± 4.174 | 122.357 ± 8.349 | 8.64 ± 8.137 | 0.009 | |
| OU1 | −51.858 ± 0.106 | 113.717 ± 0.212 | 0 ± 0 | 0.731 | |
| OUM ** | −48.924 ± 2.577 | 122.848 ± 5.155 | 9.131 ± 4.943 | 0.007 | |
| EB * | −55.173 ± 1.498 | 120.347 ± 2.997 | 6.63 ± 2.785 | 0.026 | |
| Bio19 | BM1 * | −51.445 ± 1.990 | 108.605 ± 3.981 | 5.604 ± 3.895 | 0.058 |
| BMM *** | −48.910 ± 4.396 | 113.821 ± 8.793 | 10.82 ± 8.707 | 0.004 | |
| OU1 | −46.500 ± 0.043 | 103.001 ± 0.086 | 0 ± 0 | 0.929 | |
| OUM *** | −44.660 ± 1.846 | 114.320 ± 3.692 | 11.319 ± 3.606 | 0.003 | |
| EB ** | −51.445 ± 1.990 | 112.891 ± 3.981 | 9.89 ± 3.895 | 0.006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popescu, S.M.; Tigae, C.; Dobrițescu, A.; Ștefănescu, D.M. Exploring the Climatic Niche Evolution of the Genus Falco (Aves: Falconidae) in Europe. Biology 2024, 13, 113. https://doi.org/10.3390/biology13020113
Popescu SM, Tigae C, Dobrițescu A, Ștefănescu DM. Exploring the Climatic Niche Evolution of the Genus Falco (Aves: Falconidae) in Europe. Biology. 2024; 13(2):113. https://doi.org/10.3390/biology13020113
Chicago/Turabian StylePopescu, Simona Mariana, Cristian Tigae, Aurelian Dobrițescu, and Dragoș Mihail Ștefănescu. 2024. "Exploring the Climatic Niche Evolution of the Genus Falco (Aves: Falconidae) in Europe" Biology 13, no. 2: 113. https://doi.org/10.3390/biology13020113
APA StylePopescu, S. M., Tigae, C., Dobrițescu, A., & Ștefănescu, D. M. (2024). Exploring the Climatic Niche Evolution of the Genus Falco (Aves: Falconidae) in Europe. Biology, 13(2), 113. https://doi.org/10.3390/biology13020113