Intra-Specific Difference in the Effect of Salinity on Physiological Performance in European Perch (Perca fluviatilis) and Its Ecological Importance for Fish in Estuaries


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Holding
2.2. Experiments
2.3. Data Analysis and Statistics
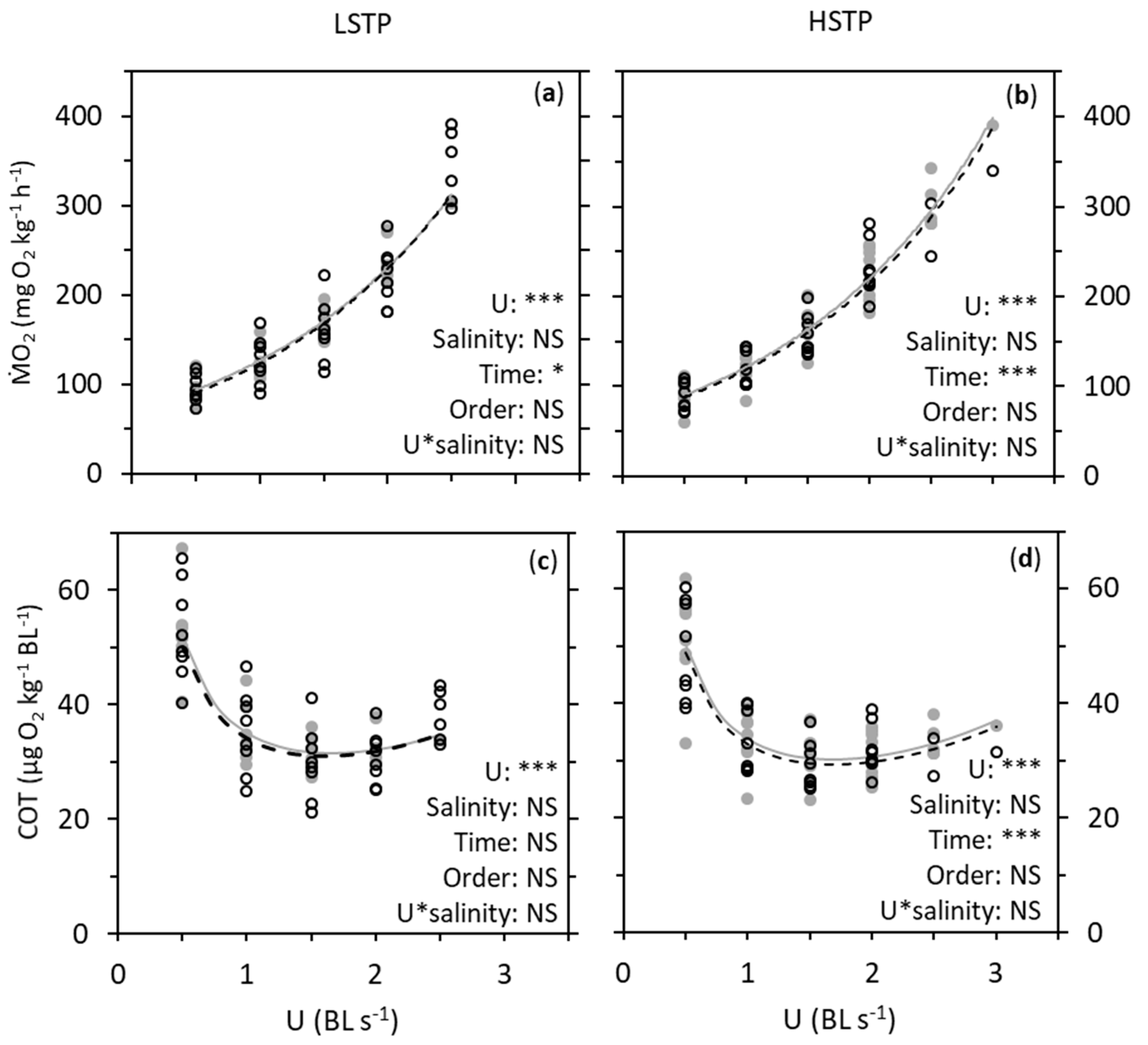
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Remane, A. Die Brackwasserfauna. Verh. Zool. Ges. 1934, 36, 34–74. [Google Scholar]
- Whitfield, A.K.; Elliott, M.; Basset, A.; Blaber, S.J.M.; West, R.J. Paradigms in estuarine ecology—A review of the Remane diagram with a suggested revised model for estuaries. Estuar. Coast. Shelf Sci. 2012, 97, 78–90. [Google Scholar] [CrossRef]
- Kennish, M.J. Ecology of Estuaries; CRC Press: Boca Raton, FL, USA, 1986; ISBN 978-0-8493-5892-0. [Google Scholar]
- Cloern, J.E. Turbidity as a control on phytoplankton biomass and productivity in estuaries. Cont. Shelf Res. 1987, 7, 1367–1381. [Google Scholar] [CrossRef]
- Brennan, R.S.; Hwang, R.; Tse, M.; Fangue, N.A.; Whitehead, A. Local adaptation to osmotic environment in killifish, Fundulus heteroclitus, is supported by divergence in swimming performance but not by differences in excess post-exercise oxygen consumption or aerobic scope. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 196, 11–19. [Google Scholar] [CrossRef]
- Gibbons, T.C.; Metzger, D.C.H.; Healy, T.M.; Schulte, P.M. Gene expression plasticity in response to salinity acclimation in threespine stickleback ecotypes from different salinity habitats. Mol. Ecol. 2017, 26, 2711–2725. [Google Scholar] [CrossRef]
- DeWitt, T.J.; Scheiner, S.M. Phenotypic plasticity—Funtional and Conceptional Approaches; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Nonaka, E.; Svanbäck, R.; Thibert-Plante, X.; Englund, G.; Brännström, Å. Mechanisms by which phenotypic plasticity affects adaptive divergence and ecological speciation. Am. Nat. 2015, 186, E126–E143. [Google Scholar] [CrossRef]
- Pfennig, D.W.; Wund, M.A.; Snell-Rood, E.C.; Cruickshank, T.; Schlichting, C.D.; Moczek, A.P. Phenotypic plasticity’s impacts on diversification and speciation. Trends Ecol. Evol. 2010, 25, 459–467. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef]
- Thorpe, J. Synopsis of Biological Data on the Perch Perca fluviatilis Linnaeus, 1758 and Perca flavescens Mitchill, 1814; FAO Fisheries Synopsis; Food and Agriculture Organization: Rome, Italy, 1977; p. 147. [Google Scholar]
- Craig, J.F. Percid Fishes: Systematics, Ecology, and Exploitation; Fish and Aquatic Resources Series; Blackwell Science: Oxford, UK; Malden, MA, USA, 2000; ISBN 978-0-632-05616-3. [Google Scholar]
- Couture, P.; Pyle, G. The Biology of Perch; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Jeppesen, E.; Peder Jensen, J.; SØndergaard, M.; Lauridsen, T.; Landkildehus, F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient: A detailed study of Danish lakes along a phosphorus gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Ljunggren, L.; Sandström, A.; Bergström, U.; Mattila, J.; Lappalainen, A.; Johansson, G.; Sundblad, G.; Casini, M.; Kaljuste, O.; Eriksson, B.K. Recruitment failure of coastal predatory fish in the Baltic Sea coincident with an offshore ecosystem regime shift. ICES J. Mar. Sci. 2010, 67, 1587–1595. [Google Scholar] [CrossRef]
- Morgan, D.L.; Hambleton, S.J.; Gill, H.S.; Beatty, S.J. Distribution, biology and likely impacts of the introduced redfin perch (Perca fluviatilis) (Percidae) in Western Australia. Mar. Freshw. Res. 2002, 53, 1211. [Google Scholar] [CrossRef]
- Skovrind, M.; Pacheco, G.; Christensen, E.A.F.; Feitz, F.; Carl, H.; Andersen, M.K.; Gilbert, M.T.P.; Møller, P.R. Population structure and natural selection of European perch in the Baltic Sea. 2019; in Preparation. [Google Scholar]
- Nilsson, J.; Andersson, J.; Karås, P.; Sandström, O. Recruitment failure and decreasing catches of perch (Perca fluviatilis L.) and pike (Esox lucius L.) in the coastal waters of southeast Sweden. Boreal Environ. Res. 2004, 9, 295–306. [Google Scholar]
- Ådjers, K.; Appelberg, M.; Eschbaum, R.; Lappalainen, A.; Minde, A.; Repe, R. Trends in coastal fish stocks of the Baltic Sea. Boreal Environ. Res. 2006, 11, 13–25. [Google Scholar]
- Christensen, E.A.F.; Grosell, M.; Steffensen, J.F. Maximum salinity tolerance and osmoregulatory capabilities of European perch Perca fluviatilis populations originating from different salinity habitats. Conserv. Physiol. 2019, 7, coz004. [Google Scholar] [CrossRef]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Lutz, P. Ionic and body compartment responses to increasing salinity in the perch Perca fluviatilis. Comp. Biochem. Physiol. A Physiol. 1972, 42, 711–717. [Google Scholar] [CrossRef]
- Brauner, C.J.; Shrimpton, J.M.; Randall, D.J. Effect of short-duration seawater exposure on plasma ion concentration and swimming performance in coho salmon (Oncorhynchus kisutch) parr. Can. J. Fisher Aquat. Sci. 1992, 49, 2399–2405. [Google Scholar] [CrossRef]
- Kolok, A.S.; Sharkey, D. Effect of freshwater acclimation on the swimming performance and plasma osmolarity of the euryhaline gulf killifish. Trans. Am. Fisher Soc. 1997, 126, 866–870. [Google Scholar] [CrossRef]
- Wolter, C.; Arlinghaus, R. Navigation impacts on freshwater fish assemblages: The ecological relevance of swimming performance. Rev. Fish Biol. Fish. 2003, 13, 63–89. [Google Scholar] [CrossRef]
- Kapoor, B.G.; Domenici, P. Escape Responses in Fish: Kinematics, Performance and Behavior. In Fish Locomotion; Science Publishers: Enfield, UK, 2010; pp. 123–170. ISBN 978-1-57808-448-7. [Google Scholar]
- Glova, G.J.; McInerney, J.E. Critical swimming speeds of coho salmon (Oncorhynchus kisutch) fry to smolt stages in relation to salinity and temperature. J. Fish. Res. Board Can. 1977, 34, 151–154. [Google Scholar] [CrossRef]
- Nelson, J.A.; Tang, Y.; Boutilier, R.G. The effects of salinity change on the exercise performance of two Atlantic cod (Gadus morhua) populations inhabiting different environments. J. Exp. Biol. 1996, 199, 1295–1309. [Google Scholar] [PubMed]
- Crespel, A.; Dupont-Prinet, A.; Bernatchez, L.; Claireaux, G.; Tremblay, R.; Audet, C. Divergence in physiological factors affecting swimming performance between anadromous and resident populations of brook charr Salvelinus fontinalis: Swimming performance in S. fontinalis. J. Fish. Biol. 2017, 90, 2170–2193. [Google Scholar] [CrossRef] [PubMed]
- Yetsko, K.; Sancho, G. The effects of salinity on swimming performance of two estuarine fishes, Fundulus heteroclitus and Fundulus majalis: Swimming performance of two estuarine fishes. J. Fish. Biol. 2015, 86, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.A.F.; Illing, B.; Iversen, N.S.; Johansen, J.L.; Domenici, P.; Steffensen, J.F. Effects of salinity on swimming performance and oxygen consumption rate of shiner perch Cymatogaster aggregata. J. Exp. Mar. Biol. Ecol. 2018, 504, 32–37. [Google Scholar] [CrossRef]
- Hvas, M.; Nilsen, T.O.; Oppedal, F. Oxygen uptake and osmotic balance of atlantic salmon in relation to exercise and salinity acclimation. Front. Mar. Sci. 2018, 5, 368. [Google Scholar] [CrossRef]
- Chabot, D.; Steffensen, J.F.; Farrell, A.P. The determination of standard metabolic rate in fishes: Measuring smr in fishes. J. Fish. Biol. 2016, 88, 81–121. [Google Scholar] [CrossRef]
- Rao, M.M.R. Oxygen consumption of rainbow trout (Salmo gairdneri) in relation to activity and salinity. Can. J. Zool. 1968, 46, 781–786. [Google Scholar] [CrossRef]
- Febry, R.; Lutz, P. Energy partitioning in fish: The activity-related cost of osmoregulation in a euryhaline cichlid. J. Exp. Biol. 1987, 128, 63–85. [Google Scholar]
- Fry, F.E.J. The Effect of Environmental Factors on the Physiology of Fish. In Fish Physiology; Elsevier: Amsterdam, The Netherlands, 1971; Volume 6, pp. 1–98. ISBN 978-0-12-350406-7. [Google Scholar]
- Farmer, G.J.; Beamish, F.W.H. Oxygen consumption of Tilapia nilotica in relation to swimming speed and salinity. J. Fish. Res. Board Can. 1969, 26, 2807–2821. [Google Scholar] [CrossRef]
- Ern, R.; Huong, D.T.T.; Cong, N.V.; Bayley, M.; Wang, T. Effect of salinity on oxygen consumption in fishes: A review: Salinity and oxygen consumption. J. Fish. Biol. 2014, 84, 1210–1220. [Google Scholar] [CrossRef]
- Sardella, B.A.; Brauner, C.J. The effect of elevated salinity on ‘California’ Mozambique tilapia (Oreochromis mossambicus x O. urolepis hornorum) metabolism. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 148, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Norin, T.; Clark, T.D. Measurement and relevance of maximum metabolic rate in fishes: Maximum metabolic rate in fishes. J. Fish. Biol. 2016, 88, 122–151. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, D.; Claireaux, G. The Effects of Environmental Factors on the Physiology of Aerobic Exercise. In Fish Locomotion; Domenici, P., Kapoor, B., Eds.; Science Publishers: Enfield, UK, 2010; pp. 296–332. ISBN 978-1-57808-448-7. [Google Scholar]
- Brix, K.V.; Grosell, M. Evaluation of pre- and post-zygotic mating barriers, hybrid fitness and phylogenetic relationship between Cyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi (Cyprinodontiformes, Teleostei). J. Evol. Biol. 2013, 26, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, A.; Roach, J.L.; Zhang, S.; Galvez, F. Genomic mechanisms of evolved physiological plasticity in killifish distributed along an environmental salinity gradient. Proc. Natl. Acad. Sci. USA 2011, 108, 6193–6198. [Google Scholar] [CrossRef] [PubMed]
- Skovrind, M.; Christensen, E.; Carl, H.; Jacobsen, L.; Møller, P. Marine spawning sites of perch Perca fluviatilis revealed by oviduct-inserted acoustic transmitters. Aquat. Biol. 2013, 19, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Skovrind, M.; Olsen, M.T.; Vieira, F.G.; Pacheco, G.; Carl, H.; Gilbert, M.T.P.; Møller, P.R. Genomic population structure of freshwater-resident and anadromous ide (Leuciscus idus) in north-western Europe. Ecol. Evol. 2016, 6, 1064–1074. [Google Scholar] [CrossRef]
- Christensen, E.A.F.; Svendsen, M.B.S.; Steffensen, J.F. Growth, migration, and physio-chemical environment of European perch (Perca fluviatilis L.) in the western Baltic Sea. 2019; in preparation. [Google Scholar]
- Fischer, H.; Matthäus, W. The importance of the Drogden Sill in the Sound for major Baltic inflows. J. Mar. Syst. 1996, 9, 137–157. [Google Scholar] [CrossRef]
- Jacobsen, L.; Bekkevold, D.; Berg, S.; Jepsen, N.; Koed, A.; Aarestrup, K.; Baktoft, H.; Skov, C. Pike (Esox lucius L.) on the edge: Consistent individual movement patterns in transitional waters of the western Baltic. Hydrobiologia 2016, 784, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Fenchel, T.; Sand-Jensen, K. Naturen i Danmark—Havet; Gyldendal: Copenhagen, Denmark, 2017. [Google Scholar]
- Bœuf, G.; Payan, P. How should salinity influence fish growth? Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 411–423. [Google Scholar] [CrossRef]
- Brett, J.R. The respiratory metabolism and swimming performance of young sockeye salmon. J. Fish. Res. Board Can. 1964, 88, 152–172. [Google Scholar] [CrossRef]
- Steffensen, J.F. Some errors in respirometry of aquatic breathers: How to avoid and correct for them. Fish. Physiol. Biochem. 1989, 6, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, M.B.S.; Bushnell, P.G.; Steffensen, J.F. Design and setup of intermittent-flow respirometry system for aquatic organisms: How to set up an aquatic respirometry system. J. Fish. Biol. 2016, 88, 26–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, E.A.F.; Svendsen, M.B.S.; Steffensen, J.F. Plasma osmolality and oxygen consumption of perch Perca fluviatilis in response to different salinities and temperatures. J. Fish. Biol. 2017, 90, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Tudorache, C.; de Boeck, G.; Claireaux, G. Forced and Preferred Swimming Speeds of Fish: A Methodological Approach. In Swimming Physiology of Fish; Palstra, A.P., Planas, J.V., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 81–108. ISBN 978-3-642-31048-5. [Google Scholar]
- Bushnell, P.G.; Steffensen, J.F.; Schurmann, H.; Jones, D.R. Exercise metabolism in two species of cod in arctic waters. Polar Biol. 1994, 14, 43–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Ojanguren, A.F.; Brana, F. Effects of size and morphology on swimming performance in juvenile brown trout (Salmo trutta L.). Ecol. Freshw. Fish. 2003, 12, 241–246. [Google Scholar] [CrossRef]
- Morgan, J.D.; Iwama, G.K. Energy cost of NaCl transport in isolated gills of cutthroat trout. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1999, 277, R631–R639. [Google Scholar] [CrossRef] [Green Version]
- Kidder, G.W.; Petersen, C.W.; Preston, R.L. Energetics of osmoregulation: I. oxygen consumption by Fundulus heteroclitus. J. Exp. Zool. 2006, 305A, 309–317. [Google Scholar] [CrossRef]
- Kirschner, L.B. Energetics of osmoregulation in fresh water vertebrates. J. Exp. Zool. 1995, 271, 243–252. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; L. Erlbaum Associates: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Segal, M.S.; Beem, E. Effect of pH, ionic charge, and osmolality on cytochromec -mediated caspase-3 activity. Am. J. Physiol. Cell Physiol. 2001, 281, C1196–C1204. [Google Scholar] [CrossRef] [Green Version]
- Nordlie, F.G. The influence of environmental salinity on respiratory oxygen demands in the euryhaline teleost, Ambassis interrupta Bleeker. Comp. Biochem. Physiol. A Physiol. 1978, 59, 271–274. [Google Scholar] [CrossRef]
- Larsen, P.F.; Nielsen, E.E.; Koed, A.; Thomsen, D.S.; Olsvik, P.A.; Loeschcke, V. Interpopulation differences in expression of candidate genes for salinity tolerance in winter migrating anadromous brown trout (Salmo trutta L.). BMC Genet. 2008, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svendsen, M.B.S.; Bushnell, P.G.; Christensen, E.A.F.; Steffensen, J.F. Sources of variation in oxygen consumption of aquatic animals demonstrated by simulated constant oxygen consumption and respirometers of different sizes: Variation in intermittent-flow respirometry. J. Fish. Biol. 2016, 88, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Norin, T.; Malte, H. Repeatability of standard metabolic rate, active metabolic rate and aerobic scope in young brown trout during a period of moderate food availability. J. Exp. Biol. 2011, 214, 1668–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, P.P.; Sun, C.M.; Wu, S.M. Changes of plasma osmolality, chloride concentration and gill Na/K-ATPase activity in tilapia Oreochromis mossambicus during seawater acclimation. Mar. Biol. 1989, 100, 295–299. [Google Scholar] [CrossRef]
- Morgan, J.D.; Sakamoto, T.; Grau, E.G.; Iwama, G.K. Physiological and respiratory responses of the mozambique tilapia (Oreochromis mossambicus) to salinity acclimation. Comp. Biochem. Physiol. A Physiol. 1997, 117, 391–398. [Google Scholar] [CrossRef]
- Van der Linden, A.; Vanaudenhove, M.; Verhoye, M.; De Boeck, G.; Blust, R. Osmoregulation of the common carp (Cyprinus carpio) when exposed to an osmotic challenge assessed in-vivo and non-invasively by diffusion- and T2-weighted magnetic resonance imaging. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1999, 124, 343–352. [Google Scholar] [CrossRef]
- Morgan, J.D.; Iwama, G.K. Cortisol-induced changes in oxygen consumption and ionic regulation in coastal cutthroat trout (Oncorhynchus clarki clarki) parr. Fish. Physiol. Biochem. 1996, 15, 385–394. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, G.; Vlaeminck, A.; Van der Linden, A.; Blust, R. The energy metabolism of common carp (Cyprinus carpio) when exposed to salt stress: An increase in energy expenditure or effects of starvation? Physiol. Biochem. Zool. 2000, 73, 102–111. [Google Scholar] [CrossRef]
- Killen, S.S. Growth trajectory influences temperature preference in fish through an effect on metabolic rate. J. Anim. Ecol. 2014, 83, 1513–1522. [Google Scholar] [CrossRef] [Green Version]
- Roche, D.G.; Binning, S.A.; Bosiger, Y.; Johansen, J.L.; Rummer, J.L. Finding the best estimates of metabolic rates in a coral reef fish. J. Exp. Biol. 2013, 216, 2103–2110. [Google Scholar] [CrossRef] [Green Version]
- Schurmann, H.; Steffensen, J.F. Effects of temperature, hypoxia and activity on the metabolism of juvenile Atlantic cod. J. Fish. Biol. 1997, 50, 1166–1180. [Google Scholar]
- Reidy, S.P.; Nelson, J.A.; Tang, Y.; Kerr, S.R. Post-exercise metabolic rate in Atlantic cod and its dependence upon the method of exhaustion. J. Fish. Biol. 1995, 47, 377–386. [Google Scholar] [CrossRef]
- Hvas, M.; Oppedal, F. Influence of experimental set-up and methodology for measurements of metabolic rates and critical swimming speed in Atlantic salmon Salmo salar. J. Fish. Biol. 2019, 95, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Hollins, J.; Thambithurai, D.; Koeck, B.; Crespel, A.; Bailey, D.M.; Cooke, S.J.; Lindström, J.; Parsons, K.J.; Killen, S.S. A physiological perspective on fisheries-induced evolution. Evol. Appl. 2018, 11, 561–576. [Google Scholar] [CrossRef] [Green Version]
- Stieglitz, J.D.; Mager, E.M.; Hoenig, R.H.; Benetti, D.D.; Grosell, M. Impacts of Deepwater Horizon crude oil exposure on adult mahi-mahi (Coryphaena hippurus) swim performance: Impacts of Deepwater Horizon exposure on mahi-mahi. Environ. Toxicol. Chem. 2016, 35, 2613–2622. [Google Scholar] [CrossRef]
- Marras, S.; Cucco, A.; Antognarelli, F.; Azzurro, E.; Milazzo, M.; Bariche, M.; Butenschön, M.; Kay, S.; Di Bitetto, M.; Quattrocchi, G.; et al. Predicting future thermal habitat suitability of competing native and invasive fish species: From metabolic scope to oceanographic modelling. Conserv. Physiol. 2015, 3, cou059. [Google Scholar] [CrossRef] [Green Version]
- Peck, M.A.; Arvanitidis, C.; Butenschön, M.; Canu, D.M.; Chatzinikolaou, E.; Cucco, A.; Domenici, P.; Fernandes, J.A.; Gasche, L.; Huebert, K.B.; et al. Projecting changes in the distribution and productivity of living marine resources: A critical review of the suite of modelling approaches used in the large European project VECTORS. Estuar. Coast. Shelf Sci. 2018, 201, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Pörtner, H.O.; Peck, M.A. Climate change effects on fishes and fisheries: Towards a cause-and-effect understanding. J. Fish. Biol. 2010, 77, 1745–1779. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Farrell, A.P. Physiology and climate change. Science 2008, 322, 690–692. [Google Scholar] [CrossRef]
- Evans, T.G.; Diamond, S.E.; Kelly, M.W. Mechanistic species distribution modelling as a link between physiology and conservation. Conserv. Physiol. 2015, 3, cov056. [Google Scholar] [CrossRef]
- Wikelski, M.; Cooke, S.J. Conservation physiology. Trends Ecol. Evol. 2006, 21, 38–46. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, D.J.; Axelsson, M.; Chabot, D.; Claireaux, G.; Cooke, S.J.; Corner, R.A.; De Boeck, G.; Domenici, P.; Guerreiro, P.M.; Hamer, B.; et al. Conservation physiology of marine fishes: State of the art and prospects for policy. Conserv. Physiol. 2016, 4, cow046. [Google Scholar] [CrossRef] [PubMed]
- Sandblom, E.; Clark, T.D.; Gräns, A.; Ekström, A.; Brijs, J.; Sundström, L.F.; Odelström, A.; Adill, A.; Aho, T.; Jutfelt, F. Physiological constraints to climate warming in fish follow principles of plastic floors and concrete ceilings. Nat. Commun. 2016, 7, 11447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, D.L.; Overgaard, J.; Wang, T.; Gesser, H.; Malte, H. Temperature effects on aerobic scope and cardiac performance of European perch (Perca fluviatilis). J. Therm. Biol. 2017, 68, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.A.F.; Skovrind, M.; Olsen, M.T.; Carl, H.; Gravlund, P.; Møller, P.R. Hatching success in brackish water of Perca fluviatilis eggs obtained from the western Baltic Sea. Cybium 2016, 6, 133–138. [Google Scholar]
- Nesbø, C.L.; Magnhagen, C.; Jakobsen, K.S. Genetic differentiation among stationary and anadromous perch (Perca fluviatilis) in the Baltic Sea. Hereditas 1999, 129, 241–249. [Google Scholar] [CrossRef]
- George, A.L.; Kuhajda, B.R.; Williams, J.D.; Cantrell, M.A.; Rakes, P.L.; Shute, J.R. Guidelines for propagation and translocation for freshwater fish conservation. Fisher 2009, 34, 529–545. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Metric | Factor | LSTP | HSTP |
|---|---|---|---|
| Ucrit (BL s−1) | 2.30 ± 0.08 | 2.40 ± 0.09 | |
| Salinity 1 | * | NS | |
| Time 2 | NS | NS | |
| Order 3 | NS | NS | |
| Uopt (µg O2 kg−1 BL−1) | 1.68 ± 0.09 | 1.68 ± 0.05 | |
| Salinity | NS | NS | |
| Time | NS | NS | |
| Order | NS | NS | |
| SMR (mg O2 kg−1 h−1) | 70.1 ± 2.5 | 69.4 ± 2.6 | |
| Respirometry method 4 | NS | NS | |
| Salinity | NS | * | |
| Time | * | *** | |
| Order | NS | NS | |
| MMR (mg O2 kg−1 h−1) | 294.8 ± 10.7 | 291.8 ± 9.9 | |
| Respirometry method | NS | NS | |
| Salinity | NS | NS | |
| Time | NS | NS | |
| Order | NS | NS | |
| Aerobic scope(mg O2 kg−1 h−1) | 294.8 ± 10.7 | 222.4 ± 9.7 | |
| Respirometry method | NS | NS | |
| Salinity | NS | NS | |
| Time | NS | NS | |
| Order | NS | NS |
| Population | Respirometry Method | Standard Deviation (%) | Sample Size | Cost of Osmoregulation (in % of SMR) |
|---|---|---|---|---|
| LSTP | Swimming | 8 | 16 | 12 |
| Static | 20 | 16 | 28 | |
| HSTP | Swimming | 7 | 16 | 10 |
| Static | 13 | 8 | 19 |
| Population | Respirometry Method | Standard Deviation (%) | Sample Size |
|---|---|---|---|
| LSTP | Swimming | 8 | 82 |
| Static | 20 | 502 | |
| HSTP | Swimming | 7 | 62 |
| Static | 13 | 212 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christensen, E.A.F.; Stieglitz, J.D.; Grosell, M.; Steffensen, J.F. Intra-Specific Difference in the Effect of Salinity on Physiological Performance in European Perch (Perca fluviatilis) and Its Ecological Importance for Fish in Estuaries. Biology 2019, 8, 89. https://doi.org/10.3390/biology8040089
Christensen EAF, Stieglitz JD, Grosell M, Steffensen JF. Intra-Specific Difference in the Effect of Salinity on Physiological Performance in European Perch (Perca fluviatilis) and Its Ecological Importance for Fish in Estuaries. Biology. 2019; 8(4):89. https://doi.org/10.3390/biology8040089
Chicago/Turabian StyleChristensen, Emil A. F., John D. Stieglitz, Martin Grosell, and John F. Steffensen. 2019. "Intra-Specific Difference in the Effect of Salinity on Physiological Performance in European Perch (Perca fluviatilis) and Its Ecological Importance for Fish in Estuaries" Biology 8, no. 4: 89. https://doi.org/10.3390/biology8040089
APA StyleChristensen, E. A. F., Stieglitz, J. D., Grosell, M., & Steffensen, J. F. (2019). Intra-Specific Difference in the Effect of Salinity on Physiological Performance in European Perch (Perca fluviatilis) and Its Ecological Importance for Fish in Estuaries. Biology, 8(4), 89. https://doi.org/10.3390/biology8040089