Time- and Zinc-Related Changes in Biomechanical Properties of Human Colorectal Cancer Cells Examined by Atomic Force Microscopy

 , ,
, ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Preparation of Zinc-Deficient Medium
2.3. Cell Culture
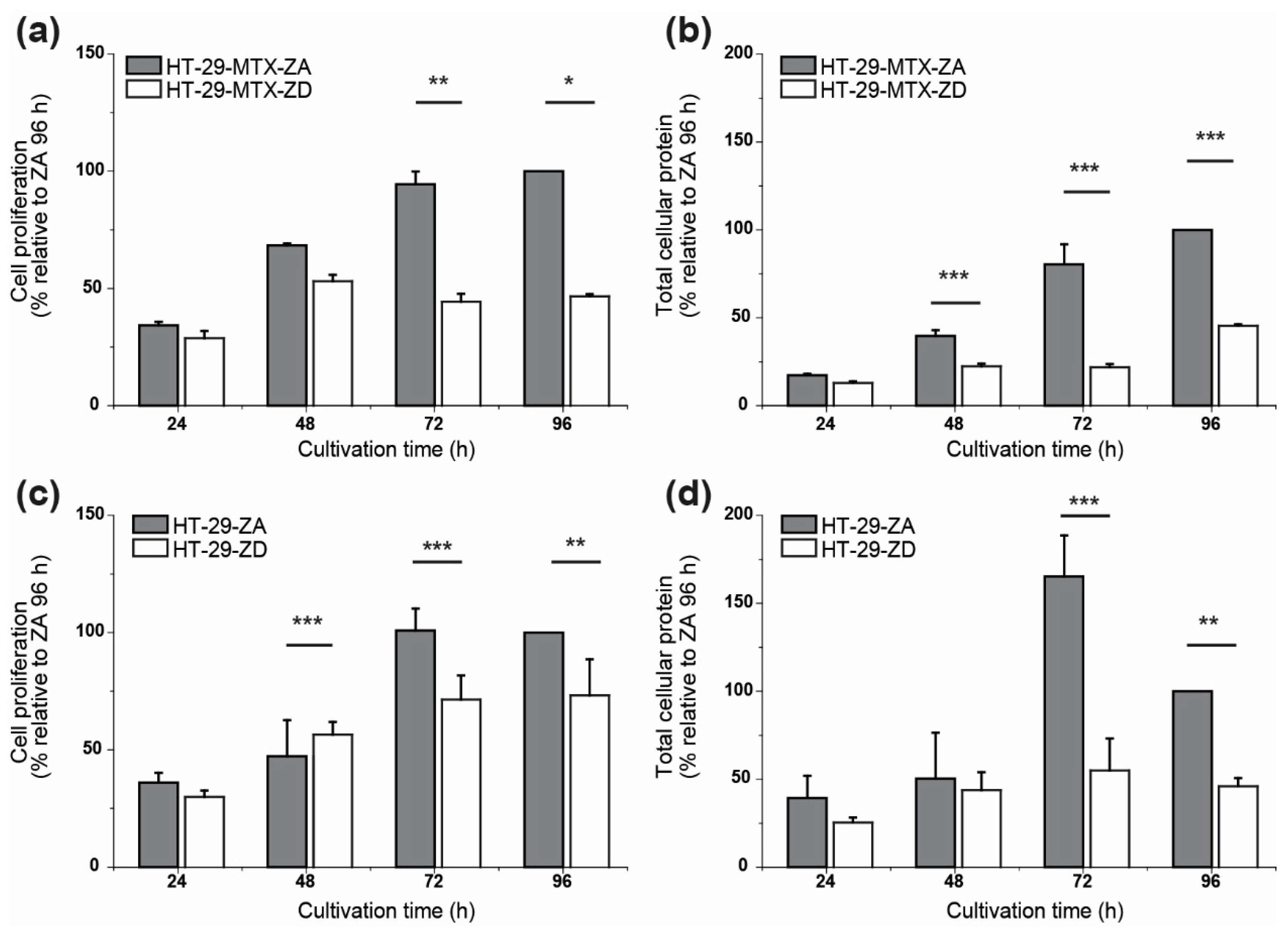
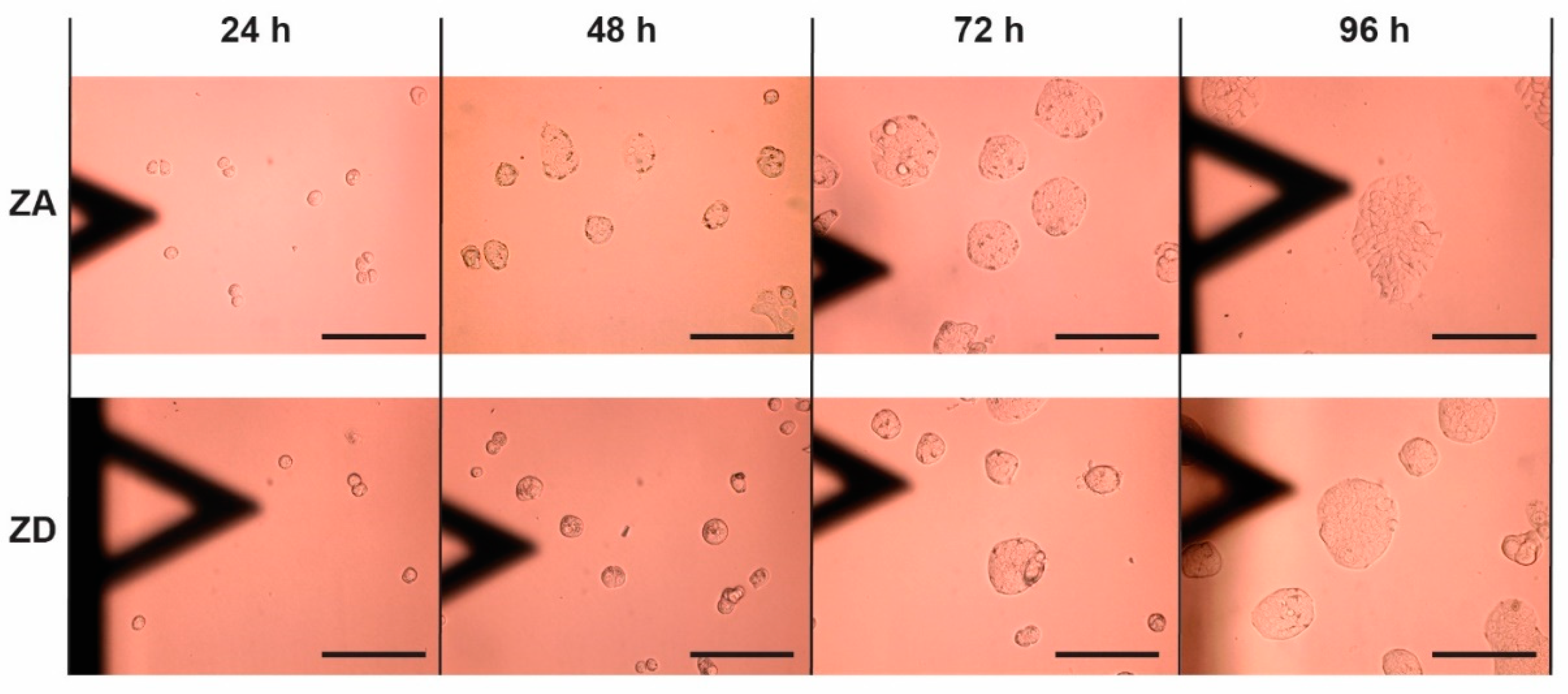
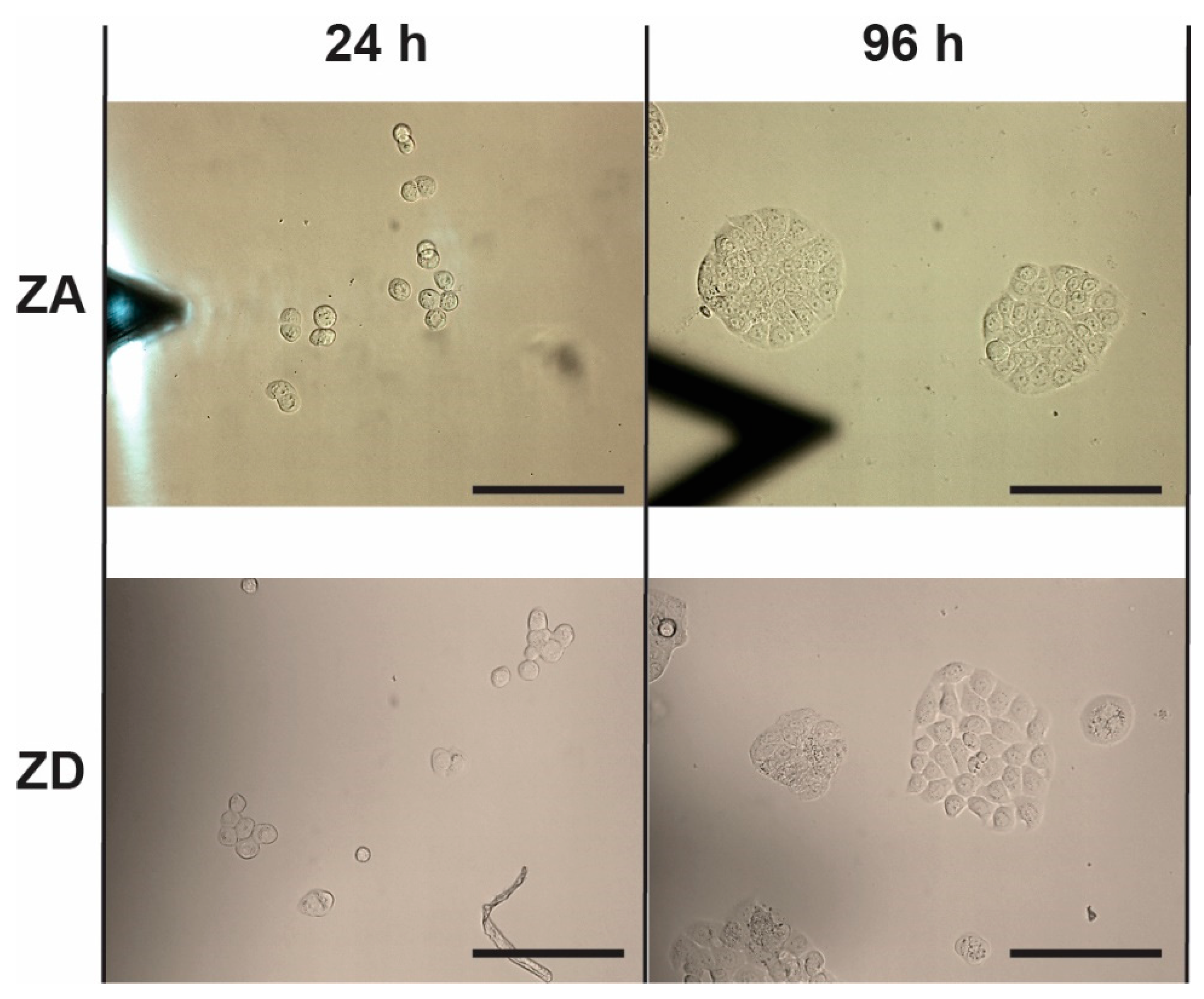
2.4. Cell Proliferation
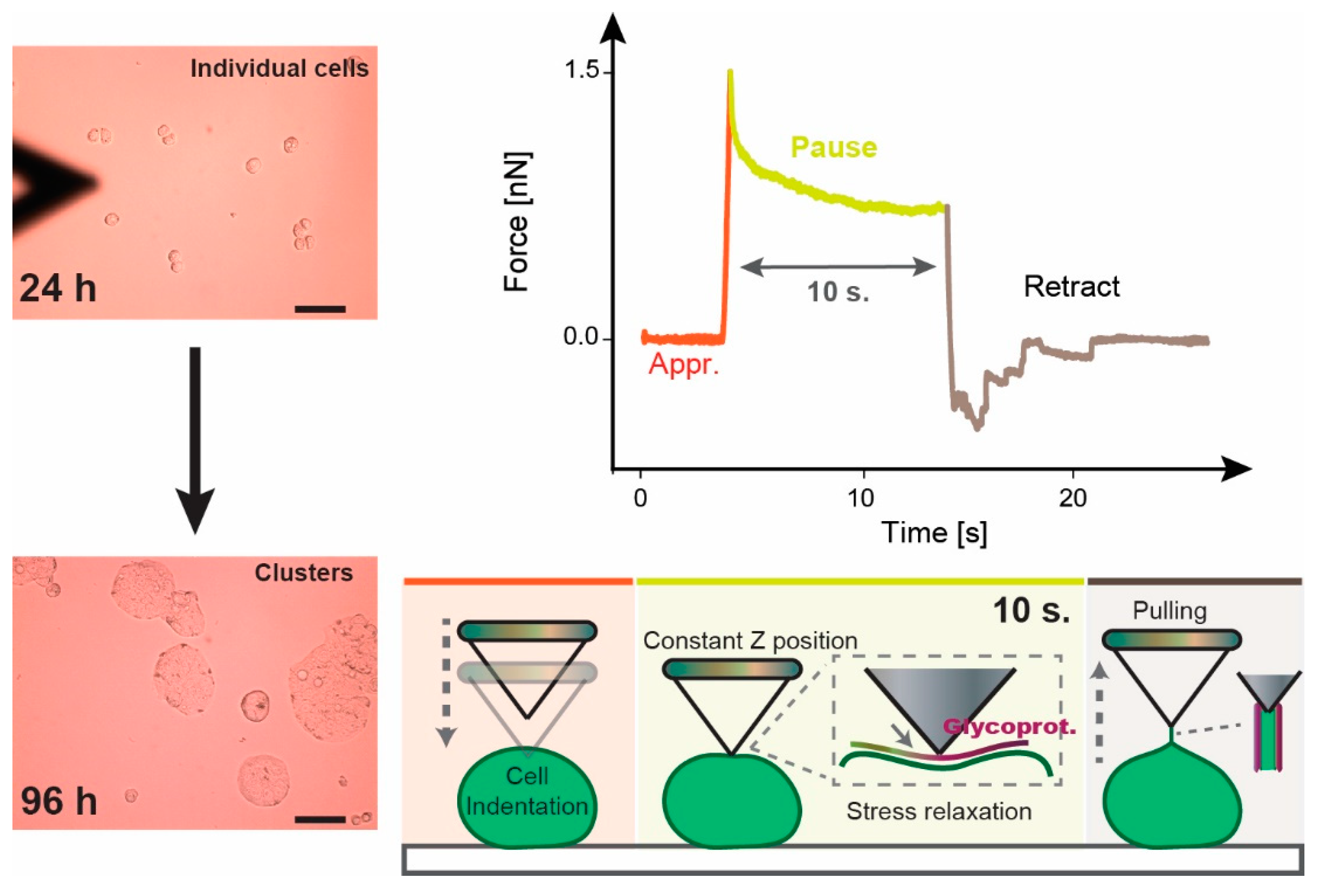
2.5. Atomic Force Microscopy (AFM)
2.6. Data Analysis
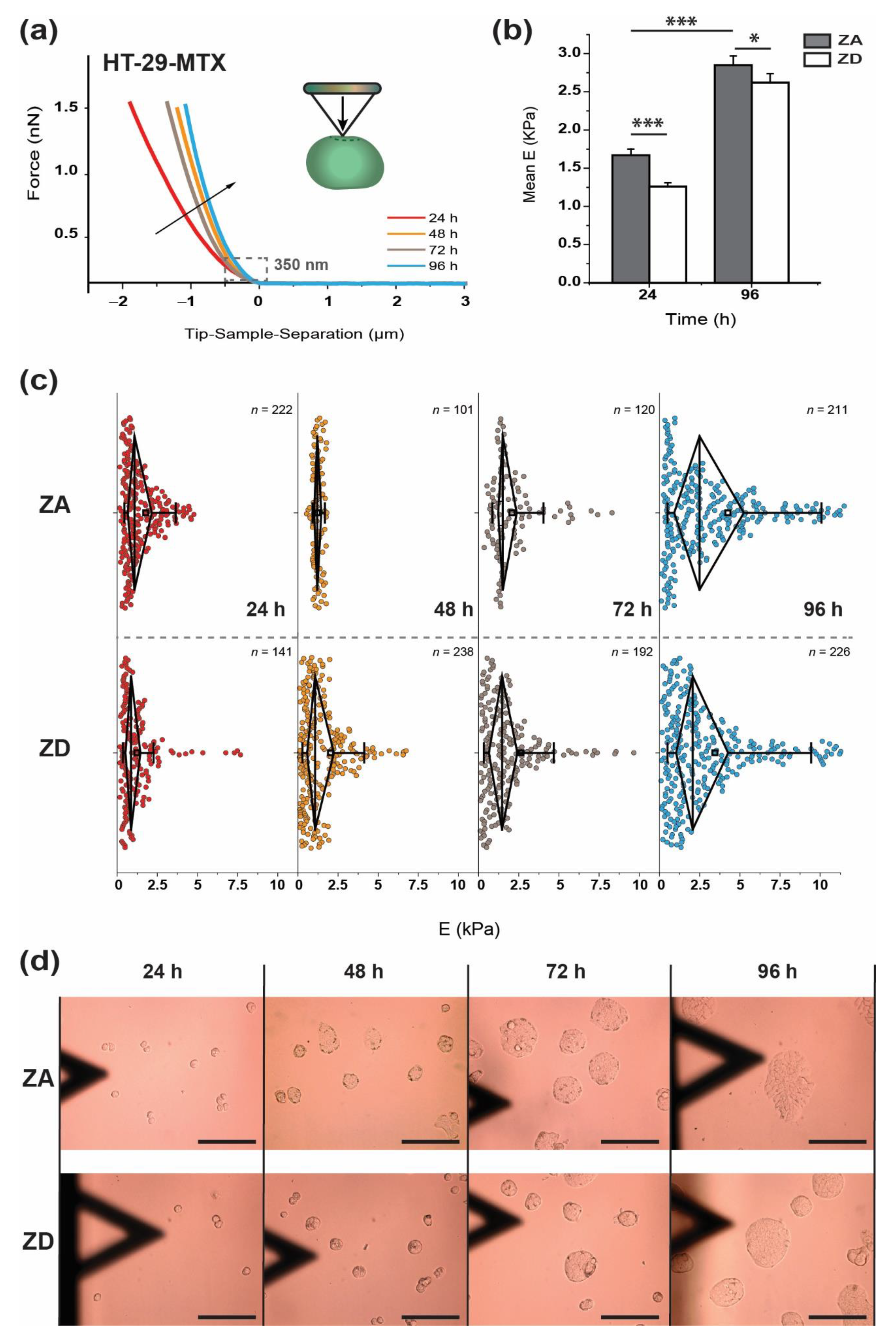
2.6.1. Elastic Modulus E
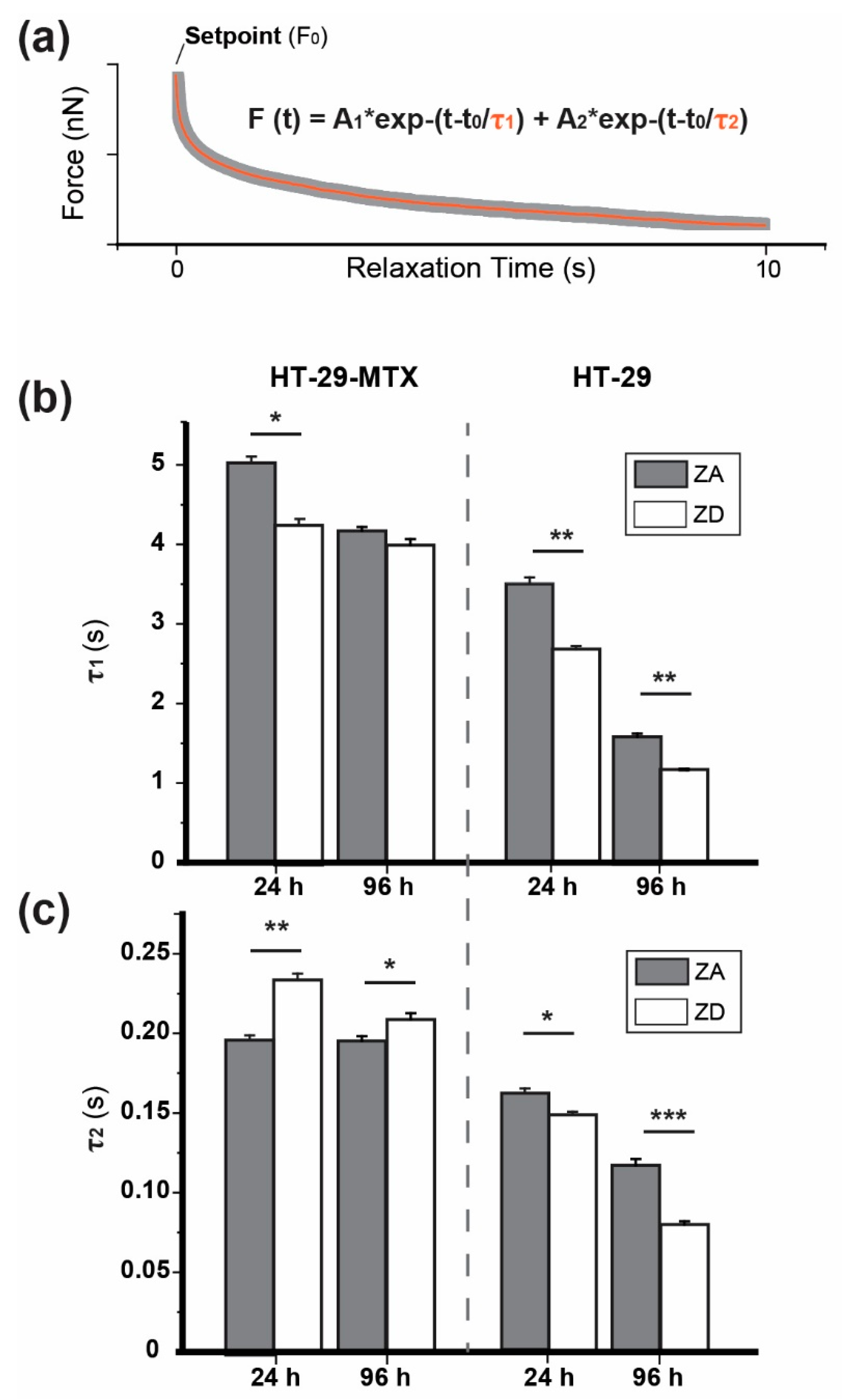
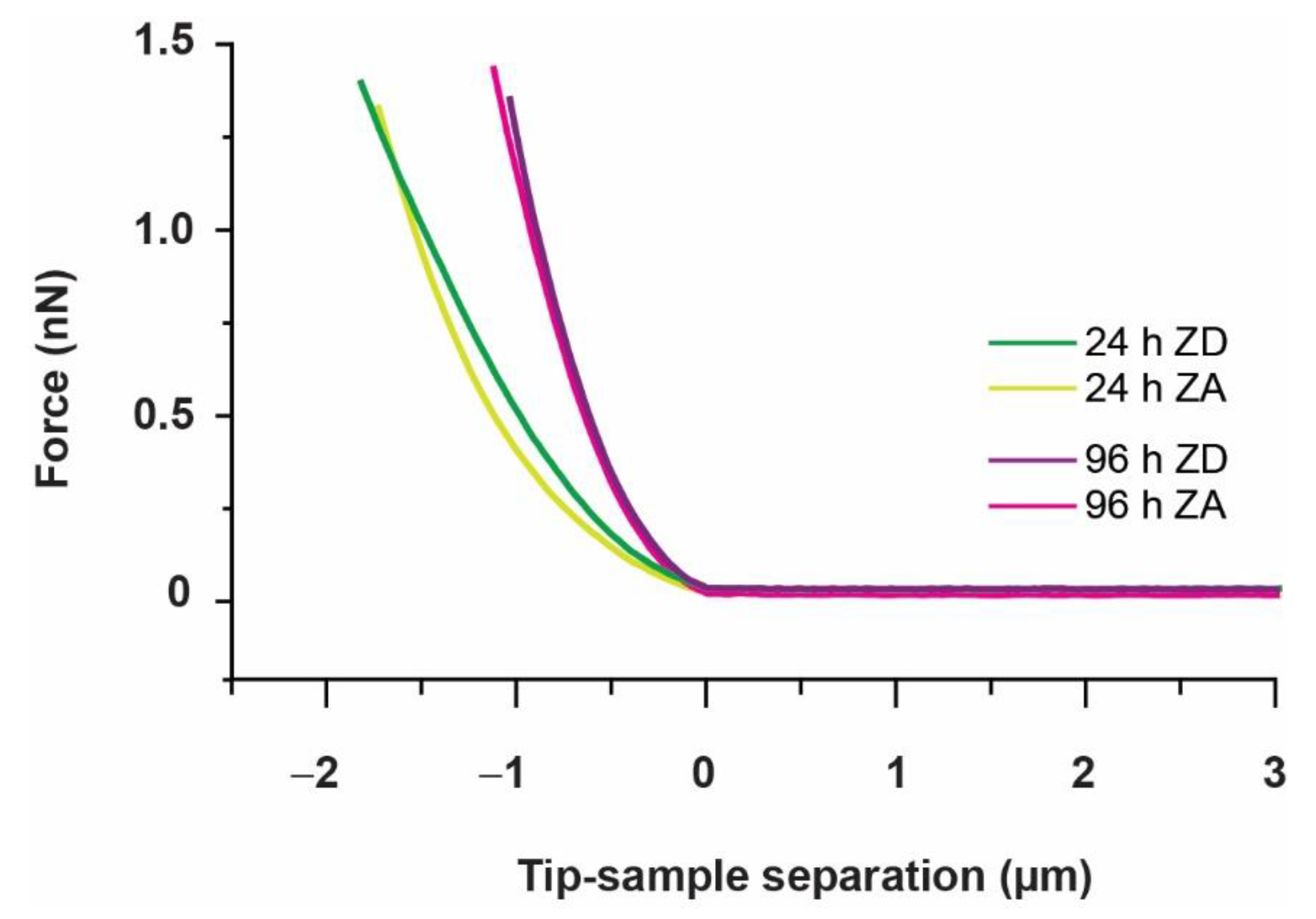
2.6.2. Stress Relaxation
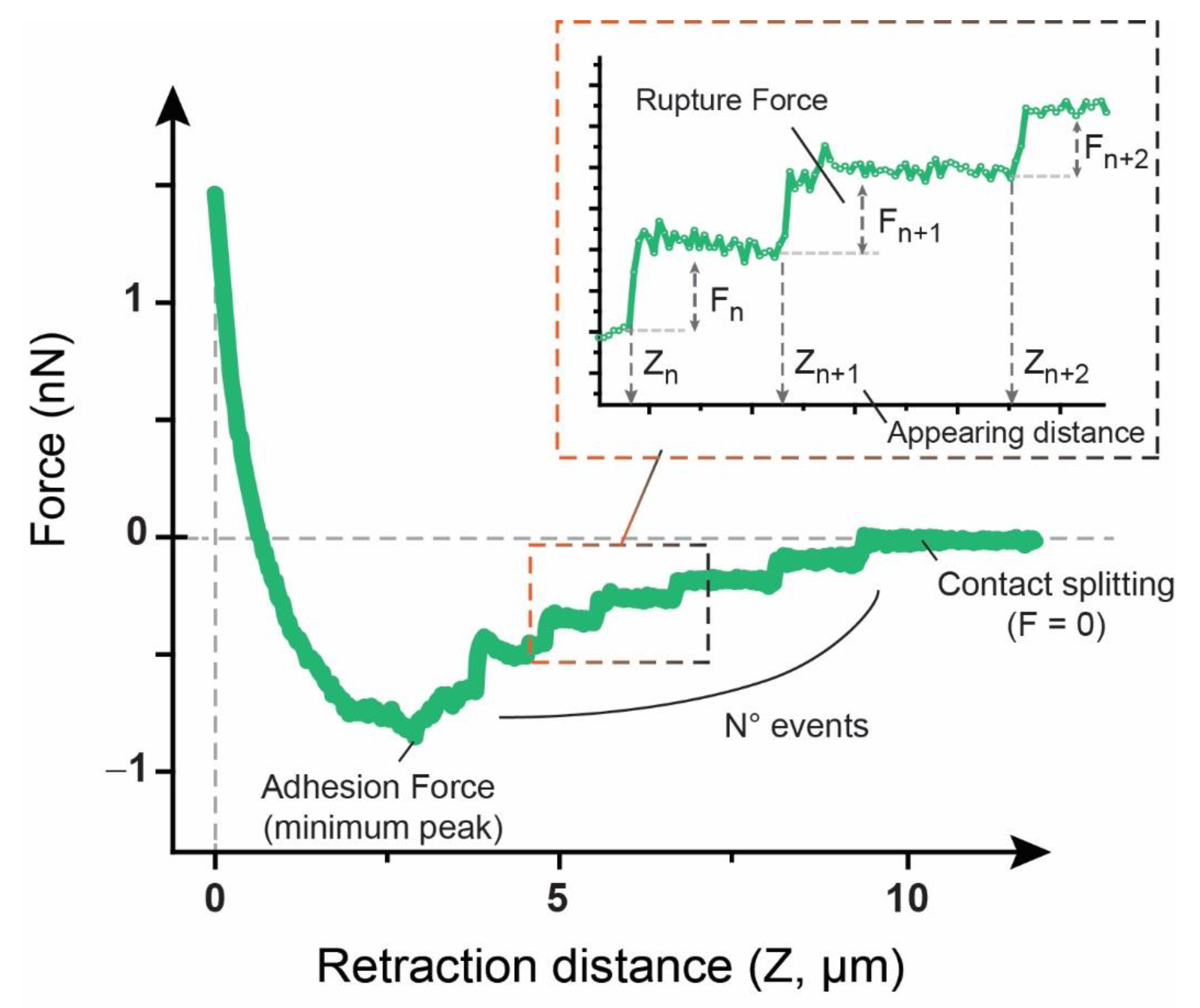
2.6.3. Adhesion Factors
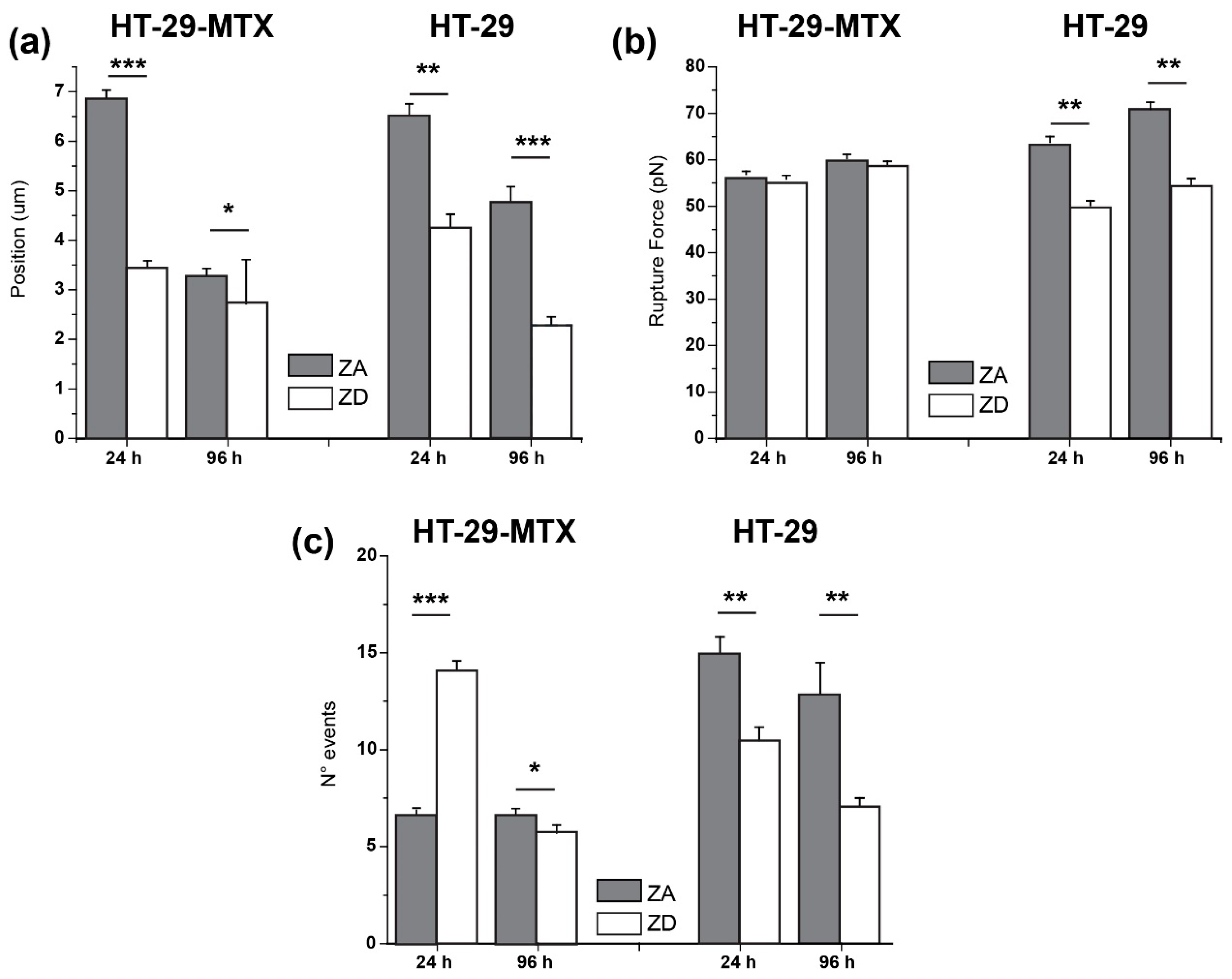
3. Results
3.1. Approach and Pause: Elastic Modulus Determination and Stress Relaxation
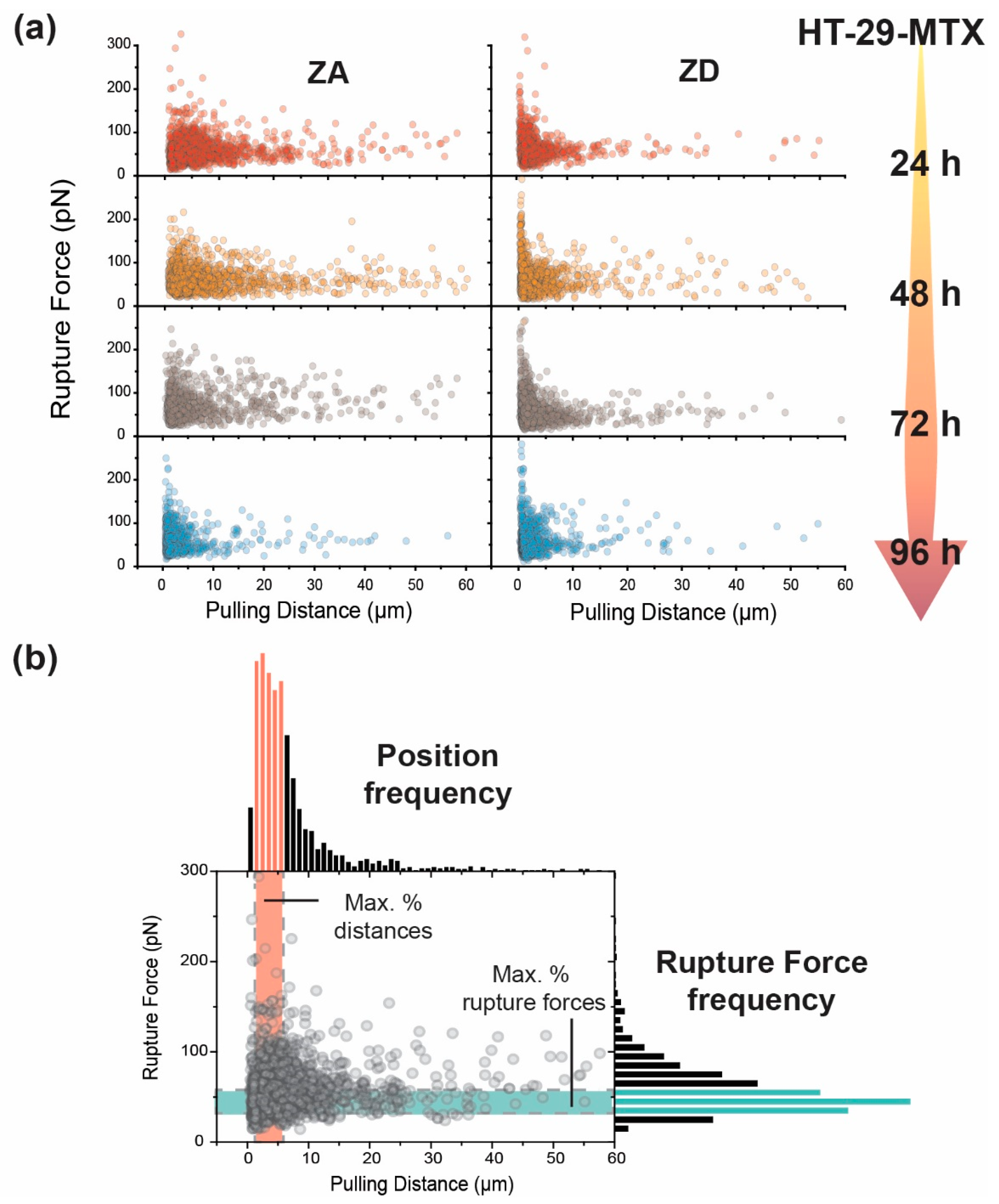
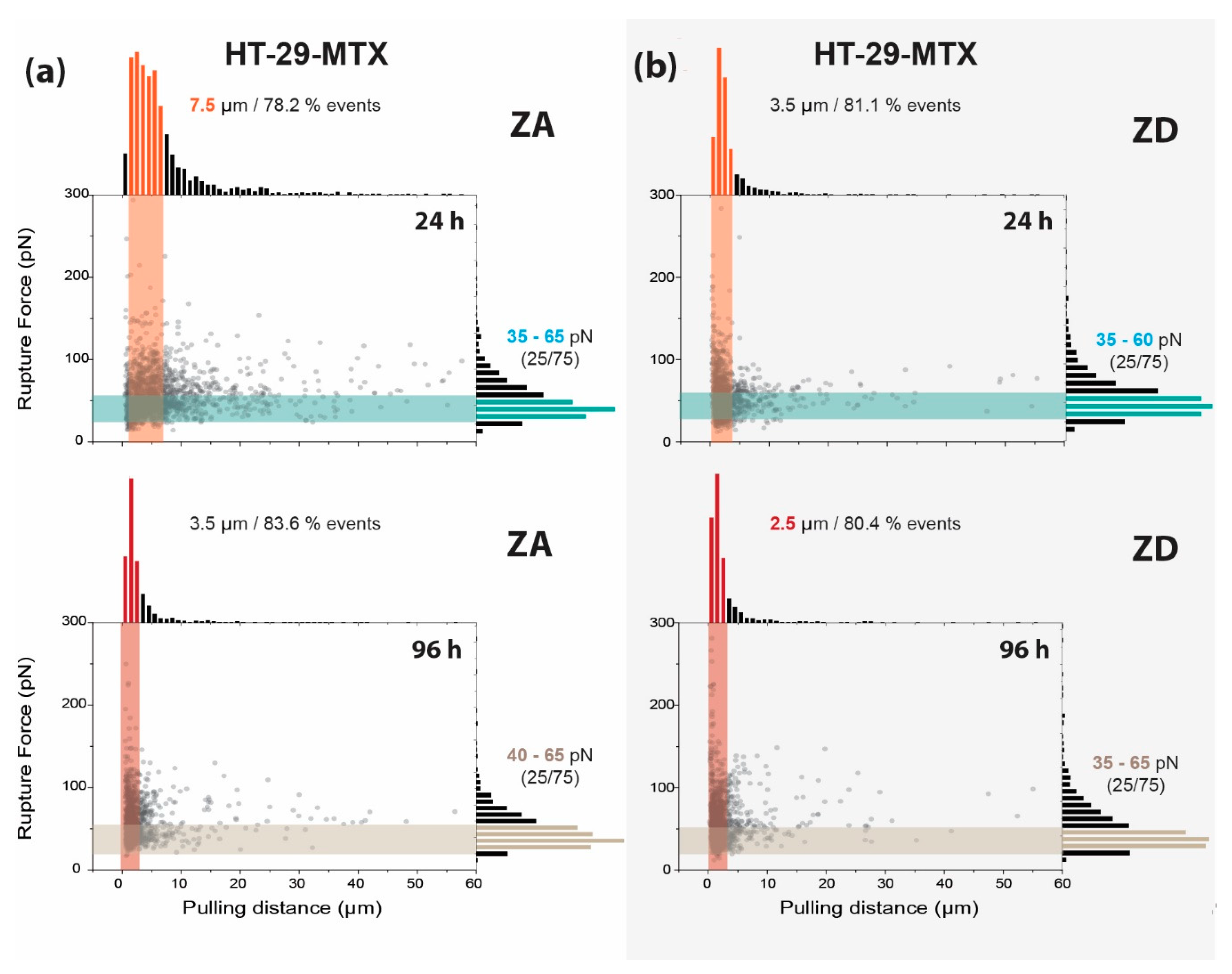
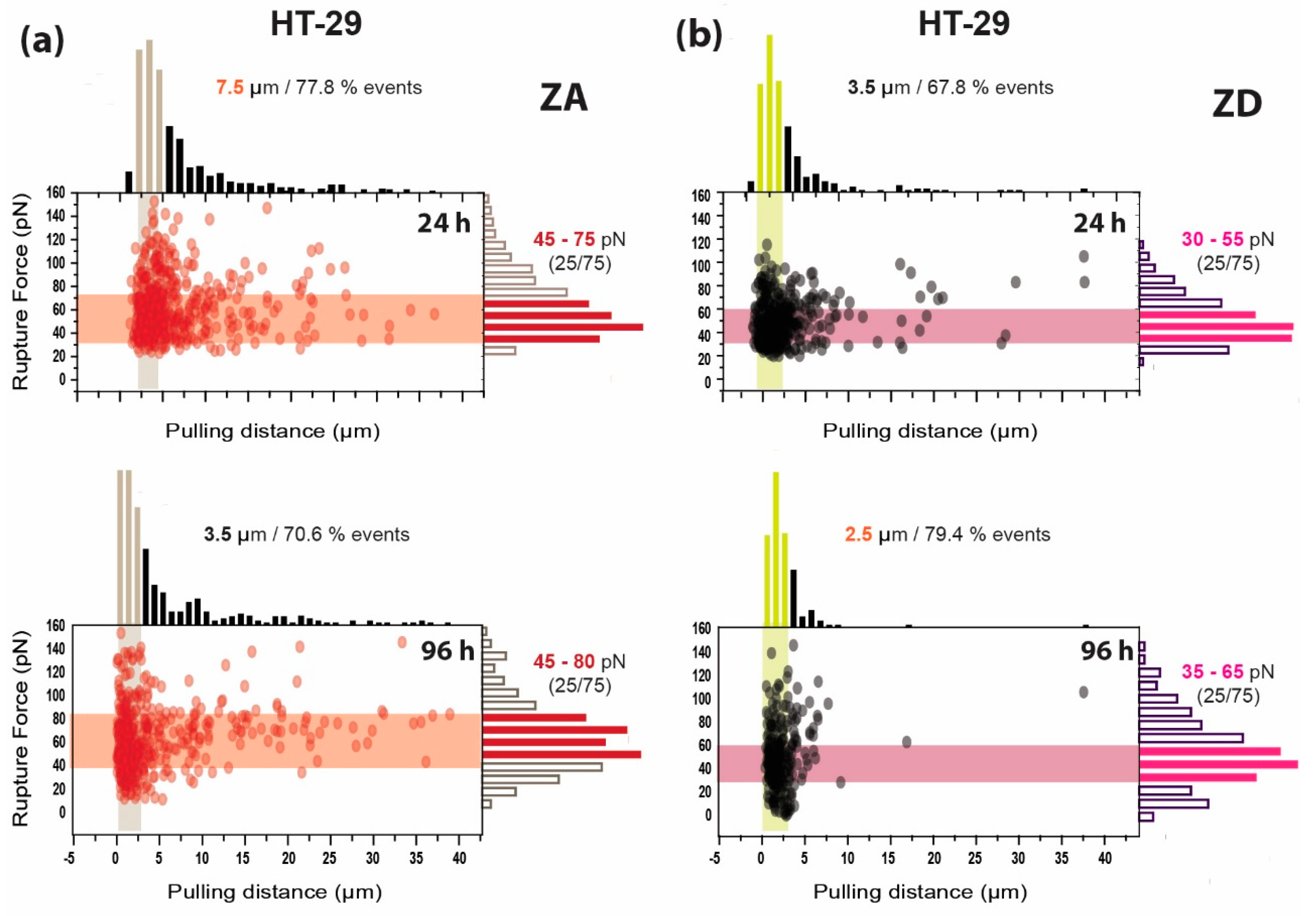
3.2. Adhesion and Rupture Events
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| 24 h | 1.67 ± 0.08 kPa | 1.26 ± 0.05 kPa | 0.91 ± 0.05 kPa | 1.09 ± 0.06 kPa |
| 48 h | 1.25 ± 0.03 kPa | 1.37 ± 0.06 kPa | ||
| 72 h | 1.58 ± 0.07 kPa | 1.62 ± 0.08 kPa | ||
| 96 h | 2.85 ± 0.12 kPa | 2.62 ± 0.12 kPa | 1.28 ± 0.17 kPa | 0.85 ± 0.08 kPa |
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| 24 h | τ1 = 5.02 ± 0.08 s | τ1 = 4.19 ± 0.06 s | τ1 = 3.51 ± 0.08 s | τ1 = 2.68 ± 0.04 s |
| τ2 = 0.19 ± 0.003 s | τ2 = 0.23 ± 0.003 s | τ2 = 0.16 ± 0.003 s | τ2 = 0.15 ± 0.002 s | |
| r2 = 0.9950 | r2 = 0.9958 | r2 = 0.9964 | r2 = 0.9976 | |
| 48 h | τ1 = 3.77 ± 0.05 s | τ1 = 4.33 ± 0.05 s | ||
| τ2 = 0.15 ± 0.004 s | τ2 = 0.16 ± 0.003 s | |||
| r2 = 0.9948 | r2 = 0.9957 | |||
| 72 h | τ1 = 5.83 ± 0.10 s | τ1 = 5.41 ± 0.12 s | ||
| τ2 = 0.24 ± 0.004 s | τ2 = 0.22 ± 0.005 s | |||
| r2 = 0.9958 | r2 = 0.9919 | |||
| 96 h | τ1 = 4.10 ± 0.05 s | τ1 = 3.99 ± 0.06 s | τ1 = 1.60 ± 0.08 s | τ1 = 1.13 ± 0.01 s |
| τ2 = 0.19 ± 0.004 s | τ2 = 0.21 ± 0.004 s | τ2 = 0.12 ± 0.004 s | τ2 = 0.08 ± 0.002 s | |
| r2 = 0.9962 | r2 = 0.9941 | r2 = 0.9839 | r2 = 0.9954 | |
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| 24 h | 0.24 ± 0.02 nN | 0.59 ± 0.03 nN | 0.61 ± 0.03 nN | 0.37 ± 0.02 nN |
| 48 h | 0.34 ± 0.01 nN | 0.31 ± 0.01 nN | ||
| 72 h | 0.44 ± 0.02 nN | 0.39 ± 0.01 nN | ||
| 96 h | 0.32 ± 0.01 nN | 0.24 ± 0.01 nN | 0.47 ± 0.07 nN | 0.33 ± 0.02 nN |
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| Mean Event Position | ||||
| 24 h | 6.86 ± 0.17 µm | 3.48 ± 0.14 µm | 6.49 ± 0.24 µm | 4.33 ± 0.23 µm |
| 48 h | 7.49 ± 0.22 µm | 3.74 ± 0.16 µm | ||
| 72 h | 7.23 ± 0.28 µm | 3.73 ± 0.14 µm | ||
| 96 h | 3.23 ± 0.15 µm | 2.69 ± 0.89 µm | 4.76 ± 0.31 µm | 2.28 ± 0.17 µm |
| Mean rupture Force | ||||
| 24 h | 56.9 ± 1.23 pN | 55.9 ± 0.76 pN | 63.8 ± 1.23 pN | 50.2 ± 1.01 pN |
| 48 h | 61.9 ± 0.65 pN | 59.4 ± 0.87 pN | ||
| 72 h | 73.0 ± 0.99 pN | 58.1 ± 0.84 pN | ||
| 96 h | 60.3 ± 0.83 pN | 58.8 ± 0.89 pN | 70.9 ± 1.49 pN | 54.3 ± 1.66 pN |
References
- Weber, A.; Iturri, J.; Benitez, R.; Zemljic-Jokhadar, S.; Toca-Herrera, J.L. Microtubule disruption changes endothelial cell mechanics and adhesion. Sci. Rep. 2019, 9, 14903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.H.; Aroush, D.R.B.; Asnacios, A.; Chen, W.C.; Dokukin, M.E.; Doss, B.L.; Durand-Smet, P.; Ekpenyong, A.; Guck, J.; Guz, N.V.; et al. A comparison of methods to assess cell mechanical properties. Nat. Methods 2018, 15, 491–498. [Google Scholar] [CrossRef]
- Moeendarbary, E.; Harris, A.R. Cell mechanics: Principles, practices, and prospects. Wiley Interdiscip. Rev. Syst. Biol. Med. 2014, 6, 371–388. [Google Scholar] [CrossRef] [PubMed]
- Yallapu, M.M.; Katti, K.S.; Katti, D.R.; Mishra, S.R.; Khan, S.; Jaggi, M.; Chauhan, S.C. The Roles of Cellular Nanomechanics in Cancer. Med. Res. Rev. 2015, 35, 198–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, S.E.; Jin, Y.S.; Rao, J.; Gimzewski, J.K. Nanomechanical analysis of cells from cancer patients. Nat. Nanotechnol. 2007, 2, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Lim, C.T. Biomechanics approaches to studying human diseases. Trends Biotechnol. 2007, 25, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Xiong, F.; Li, X.; Xiang, B.; Li, Z.; Wu, X.; Guo, C.; Li, X.; Li, Y.; Li, G.; et al. Application of atomic force microscopy in cancer research. J. Nanobiotechnol. 2018, 16, 102. [Google Scholar] [CrossRef] [Green Version]
- Lekka, M. Discrimination Between Normal and Cancerous Cells Using AFM. Bionanoscience 2016, 6, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Stylianou, A.; Lekka, M.; Stylianopoulos, T. AFM assessing of nanomechanical fingerprints for cancer early diagnosis and classification: From single cell to tissue level. Nanoscale 2018, 10, 20930–20945. [Google Scholar] [CrossRef]
- Okegawa, T.; Pong, R.C.; Li, Y.; Hsieh, J.T. The role of cell adhesion molecule in cancer progression and its application in cancer therapy. Acta Biochim. Pol. 2004, 51, 445–457. [Google Scholar] [CrossRef] [Green Version]
- Bendas, G.; Borsig, L. Cancer Cell Adhesion and Metastasis: Selectins, Integrins, and the Inhibitory Potential of Heparins. Int. J. Cell Biol. 2012, 2012, 676731. [Google Scholar] [CrossRef] [PubMed]
- Janiszewska, M.; Primi, M.C.; Izard, T. Cell adhesion in cancer: Beyond the migration of single cells. J. Biol. Chem. 2020, 295, 2495–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Brackenbury, W.J. Membrane potential and cancer progression. Front. Physiol. 2013, 4, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampen, K.R. Membrane proteins: The key players of a cancer cell. J. Membr. Biol. 2011, 242, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, M.C.; de Geus, S.W.; Prevoo, H.A.; Hawinkels, L.J.; van de Velde, C.J.; Kuppen, P.J.; Vahrmeijer, A.L.; Sier, C.F. Selecting Targets for Tumor Imaging: An Overview of Cancer-Associated Membrane Proteins. Biomark Cancer 2016, 8, 119–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.Y.; Lee, C.H.; Chuang, Y.H.; Lee, J.Y.; Chiu, Y.Y.; Wu Lee, Y.H.; Jong, Y.J.; Hwang, J.K.; Huang, S.H.; Chen, L.C.; et al. Membrane protein-regulated networks across human cancers. Nat. Commun. 2019, 10, 3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plodinec, M.; Loparic, M.; Monnier, C.A.; Obermann, E.C.; Zanetti-Dallenbach, R.; Oertle, P.; Hyotyla, J.T.; Aebi, U.; Bentires-Alj, M.; Lim, R.Y.H.; et al. The nanomechanical signature of breast cancer. Nat. Nanotechnol. 2012, 7, 757–765. [Google Scholar] [CrossRef]
- Iturri, J.; Weber, A.; Moreno-Cencerrado, A.; Vivanco, M.d.; Benítez, R.; Leporatti, S.; Toca-Herrera, J.L. Resveratrol-Induced Temporal Variation in the Mechanical Properties of MCF-7 Breast Cancer Cells Investigated by Atomic Force Microscopy. Int. J. Mol. Sci. 2019, 20, 3275. [Google Scholar] [CrossRef] [Green Version]
- Raudenska, M.; Kratochvilova, M.; Vicar, T.; Gumulec, J.; Balvan, J.; Polanska, H.; Pribyl, J.; Masarik, M. Cisplatin enhances cell stiffness and decreases invasiveness rate in prostate cancer cells by actin accumulation. Sci. Rep. 2019, 9, 1660. [Google Scholar] [CrossRef]
- Graham, H.K.; Hodson, N.W.; Hoyland, J.A.; Millward-Sadler, S.J.; Garrod, D.; Scothern, A.; Griffiths, C.E.M.; Watson, R.E.B.; Cox, T.R.; Erler, J.T.; et al. Tissue section AFM: In situ ultrastructural imaging of native biomolecules. Matrix Biol. 2010, 29, 254–260. [Google Scholar] [CrossRef]
- Brauchle, E.; Kasper, J.; Daum, R.; Schierbaum, N.; Falch, C.; Kirschniak, A.; Schäffer, T.E.; Schenke-Layland, K. Biomechanical and biomolecular characterization of extracellular matrix structures in human colon carcinomas. Matrix Biol. 2018, 68, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Sotres, J.; Jankovskaja, S.; Wannerberger, K.; Arnebrant, T. Ex-Vivo Force Spectroscopy of Intestinal Mucosa Reveals the Mechanical Properties of Mucus Blankets. Sci. Rep. 2017, 7, 7270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghajanova, A.H.; Safarzadeh, A. Atomic Force Microscopy in the Study of Cell Membranes Normal Epithelium and Adenocarcinoma Cell of the Large Intestine. J. Cancer Sci. Ther. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Schimpel, C.; Werzer, O.; Frohlich, E.; Leitinger, G.; Absenger-Novak, M.; Teubl, B.; Zimmer, A.; Roblegg, E. Atomic force microscopy as analytical tool to study physico-mechanical properties of intestinal cells. Beilstein J. Nanotechnol. 2015, 6, 1457–1466. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Nieto, M.; Mendoza-Flores, P.; García-Ortiz, D.; Montes-de-Oca, L.M.; Mendoza-Villa, M.; Barrón-González, P.; Espinosa, G.; Menchaca, J.L. Viscoelastic properties of doxorubicin-treated HT-29 cancer cells by atomic force microscopy: The fractional Zener model as an optimal viscoelastic model for cells. Biomech. Model. Mechanobiol. 2020, 19, 801–813. [Google Scholar] [CrossRef]
- Mayne, S.T.; Playdon, M.C.; Rock, C.L. Diet, nutrition, and cancer: Past, present and future. Nat. Rev. Clin. Oncol. 2016, 13, 504–515. [Google Scholar] [CrossRef]
- Maret, W. Zinc Biochemistry: From a Single Zinc Enzyme to a Key Element of Life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Ho, E. Zinc deficiency, DNA damage and cancer risk. J. Nutr. Biochem. 2004, 15, 572–578. [Google Scholar] [CrossRef]
- Dhawan, D.K.; Chadha, V.D. Zinc: A promising agent in dietary chemoprevention of cancer. Indian J. Med. Res. 2010, 132, 676–682. [Google Scholar]
- Dani, V.; Goel, A.; Vaiphei, K.; Dhawan, D.K. Chemopreventive potential of zinc in experimentally induced colon carcinogenesis. Toxicol. Lett. 2007, 171, 10–18. [Google Scholar] [CrossRef]
- Maares, M.; Haase, H. A Guide to Human Zinc Absorption: General Overview and Recent Advances of In Vitro Intestinal Models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, H.; Ellinger, S.; Linseisen, J.; Neuhäuser-Berthold, M.; Richter, M. Revised D-A-CH-reference values for the intake of zinc. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. 2020, 61, 126536. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Giovannucci, E.L.; Smith-Warner, S.A.; Wu, K.; Fuchs, C.S.; Pollak, M.; Willett, W.C.; Ma, J. A prospective study of intakes of zinc and heme iron and colorectal cancer risk in men and women. Cancer Causes Control 2011, 22, 1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepien, M.; Jenab, M.; Freisling, H.; Becker, N.P.; Czuban, M.; Tjønneland, A.; Olsen, A.; Overvad, K.; Boutron-Ruault, M.C.; Mancini, F.R.; et al. Pre-diagnostic copper and zinc biomarkers and colorectal cancer risk in the European Prospective Investigation into Cancer and Nutrition cohort. Carcinogenesis 2017, 38, 699–707. [Google Scholar] [CrossRef]
- Li, P.; Xu, J.; Shi, Y.; Ye, Y.; Chen, K.; Yang, J.; Wu, Y. Association between zinc intake and risk of digestive tract cancers: A systematic review and meta-analysis. Clin. Nutr. 2014, 33, 415–420. [Google Scholar] [CrossRef]
- Barresi, V.; Valenti, G.; Spampinato, G.; Musso, N.; Castorina, S.; Rizzarelli, E.; Condorelli, D.F. Transcriptome analysis reveals an altered expression profile of zinc transporters in colorectal cancer. J. Cell. Biochem. 2018, 119, 9707–9719. [Google Scholar] [CrossRef]
- Bafaro, E.; Liu, Y.; Xu, Y.; Dempski, R.E. The emerging role of zinc transporters in cellular homeostasis and cancer. Signal Transduct. Target. Ther. 2017, 2, 17029. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Amin, K.; Sato, B.G.; Samuelsson, S.J.; Sambucetti, L.; Haroon, Z.A.; Laderoute, K.; Murphy, B.J. The metal-responsive transcription factor-1 protein is elevated in human tumors. Cancer Biol. Ther. 2010, 9, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Ninsontia, C.; Phiboonchaiyanan, P.P.; Chanvorachote, P. Zinc induces epithelial to mesenchymal transition in human lung cancer H460 cells via superoxide anion-dependent mechanism. Cancer Cell Int. 2016, 16, 48. [Google Scholar] [CrossRef] [PubMed]
- Hogstrand, C.; Kille, P.; Ackland, M.L.; Hiscox, S.; Taylor, K.M. A mechanism for epithelial-mesenchymal transition and anoikis resistance in breast cancer triggered by zinc channel ZIP6 and STAT3 (signal transducer and activator of transcription 3). Biochem. J. 2013, 455, 229–237. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, H.; Xu, Z.; Cheng, X. Zinc dysregulation in cancers and its potential as a therapeutic target. Cancer Biol. Med. 2020, 17, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Stovall, D.B.; Wang, W.; Sui, G. Advances of Zinc Signaling Studies in Prostate Cancer. Int. J. Mol. Sci. 2020, 21, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumulec, J.; Masarik, M.; Adam, V.; Eckschlager, T.; Provaznik, I.; Kizek, R. Serum and Tissue Zinc in Epithelial Malignancies: A Meta-Analysis. PLoS ONE 2014, 9, e99790. [Google Scholar] [CrossRef] [PubMed]
- Grattan, B.J.; Freake, H.C. Zinc and cancer: Implications for LIV-1 in breast cancer. Nutrients 2012, 4, 648–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christudoss, P.; Selvakumar, R.; Pulimood, A.B.; Fleming, J.J.; Mathew, G. Zinc and zinc related enzymes in precancerous and cancerous tissue in the colon of dimethyl hydrazine treated rats. Asian Pac. J. Cancer Prev. 2012, 13, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.S.; Bao, B. Molecular Mechanisms of Zinc as a Pro-Antioxidant Mediator: Clinical Therapeutic Implications. Antioxidants 2019, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Finamore, A.; Massimi, M.; Conti Devirgiliis, L.; Mengheri, E. Zinc deficiency induces membrane barrier damage and increases neutrophil transmigration in Caco-2 cells. J. Nutr. 2008, 138, 1664–1670. [Google Scholar] [CrossRef] [Green Version]
- Tejeda-Mora, H.; Stevens, L.; Gröllers, M.; Katan, A.; van de Steeg, E.; van der Heiden, M. AFM based elasticity of intestinal epithelium correlate with barrier function under drug action. bioRxiv 2019. [Google Scholar] [CrossRef]
- Ciasca, G.; Papi, M.; Minelli, E.; Palmieri, V.; De Spirito, M. Changes in cellular mechanical properties during onset or progression of colorectal cancer. World J. Gastroenterol. 2016, 22, 7203–7214. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Ophir, I.; Shaul, Y.B. Induced differentiation in HT29, a human colon adenocarcinoma cell line. J. Cell Sci. 1999, 112, 2657–2666. [Google Scholar] [PubMed]
- Fogh, J. Human tumor lines for cancer research. Cancer Investig. 1986, 4, 157–184. [Google Scholar] [CrossRef] [PubMed]
- Schroy, P.C.; Rustgi, A.K.; Ikonomu, E.; Liu, X.P.; Polito, J.; Andry, C.; O’Keane, J.C. Growth and intestinal differentiation are independently regulated in HT29 colon cancer cells. J. Cell. Physiol. 1994, 161, 111–123. [Google Scholar] [CrossRef]
- Lesuffleur, T.; Porchet, N.; Aubert, J.P.; Swallow, D.; Gum, J.R.; Kim, Y.S.; Real, F.X.; Zweibaum, A. Differential expression of the human mucin genes MUC1 to MUC5 in relation to growth and differentiation of different mucus-secreting HT-29 cell subpopulations. J. Cell Sci. 1993, 106, 771–783. [Google Scholar]
- Behrens, I.; Stenberg, P.; Artursson, P.; Kissel, T. Transport of Lipophilic Drug Molecules in a New Mucus-Secreting Cell Culture Model Based on HT29-MTX Cells. Pharm. Res. 2001, 18, 1138–1145. [Google Scholar] [CrossRef]
- Hennebicq-Reig, S.; Tetaert, D.; Soudan, B.; Kim, I.; Huet, G.; Briand, G.; Richet, C.; Demeyer, D.; Degand, P. O-Glycosylation and cellular differentiation in a subpopulation of mucin-secreting HT-29 cell line. Exp. Cell Res. 1997, 235, 100–107. [Google Scholar] [CrossRef]
- Maares, M.; Keil, C.; Straubing, S.; Robbe-Masselot, C.; Haase, H. Zinc Deficiency Disturbs Mucin Expression, O-Glycosylation and Secretion by Intestinal Goblet Cells. Int. J. Mol. Sci. 2020, 21, 6149. [Google Scholar] [CrossRef]
- Kindermann, B.; Doöring, F.; Pfaffl, M.; Daniel, H. Identification of Genes Responsive to Intracellular Zinc Depletion in the Human Colon Adenocarcinoma Cell Line HT-291. J. Nutr. 2004, 134, 57–62. [Google Scholar] [CrossRef]
- Krezel, A.; Maret, W. Zinc-buffering capacity of a eukaryotic cell at physiological pZn. J. Biol. Inorg. Chem. 2006, 11, 1049–1062. [Google Scholar] [CrossRef]
- Maares, M.; Keil, C.; Koza, J.; Straubing, S.; Schwerdtle, T.; Haase, H. In Vitro Studies on Zinc Binding and Buffering by Intestinal Mucins. Int. J. Mol. Sci. 2018, 19, 2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maares, M.; Keil, C.; Thomsen, S.; Gunzel, D.; Wiesner, B.; Haase, H. Characterization of Caco-2 cells stably expressing the protein-based zinc probe eCalwy-5 as a model system for investigating intestinal zinc transport. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. 2018, 49, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Iturri, J.; Benitez, R.; Toca-Herrera, J.L. Measuring biomaterials mechanics with atomic force microscopy. 1. Influence of the loading rate and applied force (pyramidal tips). Microsc. Res. Tech. 2019, 82, 1392–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Flores, S.; Benitez, R.; Vivanco, M.; Toca-Herrera, J.L. Stress relaxation and creep on living cells with the atomic force microscope: A means to calculate elastic moduli and viscosities of cell components. Nanotechnology 2010, 21, 445101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alker, W.; Schwerdtle, T.; Schomburg, L.; Haase, H. A Zinpyr-1-based Fluorimetric Microassay for Free Zinc in Human Serum. Int. J. Mol. Sci. 2019, 20, 4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demichelis, A.; Divieto, C.; Mortati, L.; Pavarelli, S.; Sassi, G.; Sassi, M.P. Toward the realization of reproducible Atomic Force Microscopy measurements of elastic modulus in biological samples. J. Biomech. 2015, 48, 1099–1104. [Google Scholar] [CrossRef]
- Sumarokova, M.; Iturri, J.; Weber, A.; Maares, M.; Keil, C.; Luis Toca-Herrera, J. Influencing the adhesion properties and wettability of mucin protein films by variation of the environmental pH. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Lawson, M.J.; Butler, R.N.; Goland, G.J.; Jarrett, I.G.; Roberts-Thomson, I.C.; Partick, E.J.; Dreosti, I.E. Zinc deficiency is associated with suppression of colonocyte proliferation in the distal large bowel of rats. Biol. Trace Elem. Res. 1988, 18, 115–121. [Google Scholar] [CrossRef]
- Wang, X.; Valenzano, M.C.; Mercado, J.M.; Zurbach, E.P.; Mullin, J.M. Zinc supplementation modifies tight junctions and alters barrier function of Caco-2 human intestinal epithelial layers. Dig. Dis. Sci. 2012, 58, 77–87. [Google Scholar] [CrossRef]
- Cohen, L.; Sekler, I.; Hershfinkel, M. The zinc sensing receptor, ZnR/GPR39, controls proliferation and differentiation of colonocytes and thereby tight junction formation in the colon. Cell Death Dis. 2014, 5, e1307. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Lee, N.G.; Lee, K.H.; Seo, J.T.; Choi, K.Y. The ERK pathway involves positive and negative regulations of HT-29 colorectal cancer cell growth by extracellular zinc. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1181–G1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.S.; Ahn, Y.; Kim, J.A.; Yun, M.S.; Seong, B.L.; Choi, K.Y. Extracellular zinc stimulates ERK-dependent activation of p21(Cip/WAF1) and inhibits proliferation of colorectal cancer cells. Br. J. Pharmacol. 2002, 137, 597–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.Y.; Park, K.S.; Kim, J.A.; Choi, K.Y. Differential modulation of zinc-stimulated p21(Cip/WAF1) and cyclin D1 induction by inhibition of PI3 kinase in HT-29 colorectal cancer cells. Exp. Mol. Med. 2002, 34, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.K.; Kwon, Y.; Jang, M.; Park, M.; Kim, J.; Cho, S.; Jang, D.G.; Lee, W.B.; Jung, S.H.; Choi, H.J.; et al. β-catenin activation down-regulates cell-cell junction-related genes and induces epithelial-to-mesenchymal transition in colorectal cancers. Sci. Rep. 2019, 9, 18440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, K.; Zhang, Y.; Niu, J.; Nie, Z.; Liu, Q.; Lv, C. Zinc promotes cell apoptosis via activating the Wnt-3a/β-catenin signaling pathway in osteosarcoma. J. Orthop. Surg. Res. 2020, 15, 57. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Han, J.; Jiang, J.; Shi, S.; Ma, X.; Liu, X.; Wang, C.; Nie, X.; He, Y.; Jiang, S.; et al. The downregulation of Wnt/β-catenin signaling pathway is associated with zinc deficiency-induced proliferative deficit of C17.2 neural stem cells. Brain Res. 2015, 1615, 61–70. [Google Scholar] [CrossRef]
- Medico, E.; Russo, M.; Picco, G.; Cancelliere, C.; Valtorta, E.; Corti, G.; Buscarino, M.; Isella, C.; Lamba, S.; Martinoglio, B.; et al. The molecular landscape of colorectal cancer cell lines unveils clinically actionable kinase targets. Nat. Commun. 2015, 6, 7002. [Google Scholar] [CrossRef]
- Dutton, J.S.; Hinman, S.S.; Kim, R.; Wang, Y.; Allbritton, N.L. Primary Cell-Derived Intestinal Models: Recapitulating Physiology. Trends Biotechnol. 2019, 37, 744–760. [Google Scholar] [CrossRef] [Green Version]









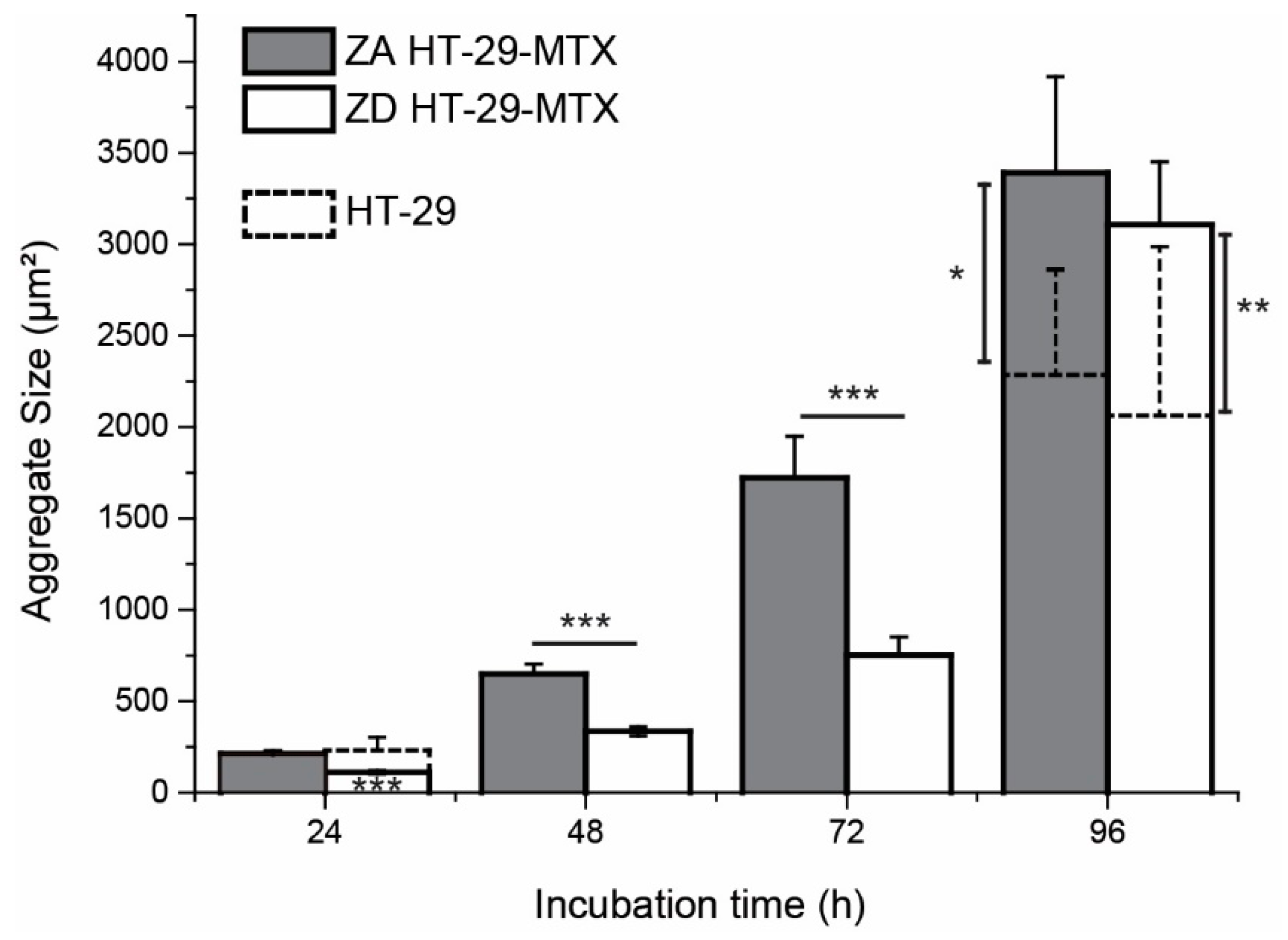
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| 24 h | 210.7 ± 14.7 µm2 | 111.0 ± 9.6 µm2 | 203.7 ± 26.2 µm2 | 231.3 ± 71.2 µm2 |
| 48 h | 648.9 ± 54.2 µm2 | 334.8 ± 25.6 µm2 | ||
| 72 h | 1721.8 ± 227.2 µm2 | 751.2 ± 100.1 µm2 | ||
| 96 h | 3390.9 ± 526.4 µm2 | 3108.1 ± 344.2 µm2 | 2286 ± 571.7 µm2 | 2064.9 ± 923.5 µm2 |
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| 24 h | 1.67 ± 0.08 kPa | 1.26 ± 0.05 kPa | 0.91 ± 0.05 kPa | 1.09 ± 0.06 kPa |
| 96 h | 2.85 ± 0.28 kPa | 2.62 ± 0.12 kPa | 1.28 ± 0.17 kPa | 0.85 ± 0.08 kPa |
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| 24 h | τ1 = 5.02 ± 0.08 s | τ1 = 4.19 ± 0.06 s | τ1 = 3.51 ± 0.08 s | τ1 = 2.68 ± 0.04 s |
| τ2 = 0.19 ± 0.003 s | τ2 = 0.23 ± 0.003 s | τ2 = 0.16 ± 0.003 s | τ2 = 0.15 ± 0.002 s | |
| r2 = 0.9950 | r2 = 0.9958 | r2 = 0.9964 | r2 = 0.9976 | |
| 96 h | τ1 = 4.10 ± 0.05 s | τ1 = 3.99 ± 0.06 s | τ1 = 1.60 ± 0.08 s | τ1 = 1.13 ± 0.01 s |
| τ2 = 0.19 ± 0.004 s | τ2 = 0.21 ± 0.004 s | τ2 = 0.12 ± 0.004 s | τ2 = 0.08 ± 0.002 s | |
| r2 = 0.9962 | r2 = 0.9941 | r2 = 0.9839 | r2 = 0.9954 | |
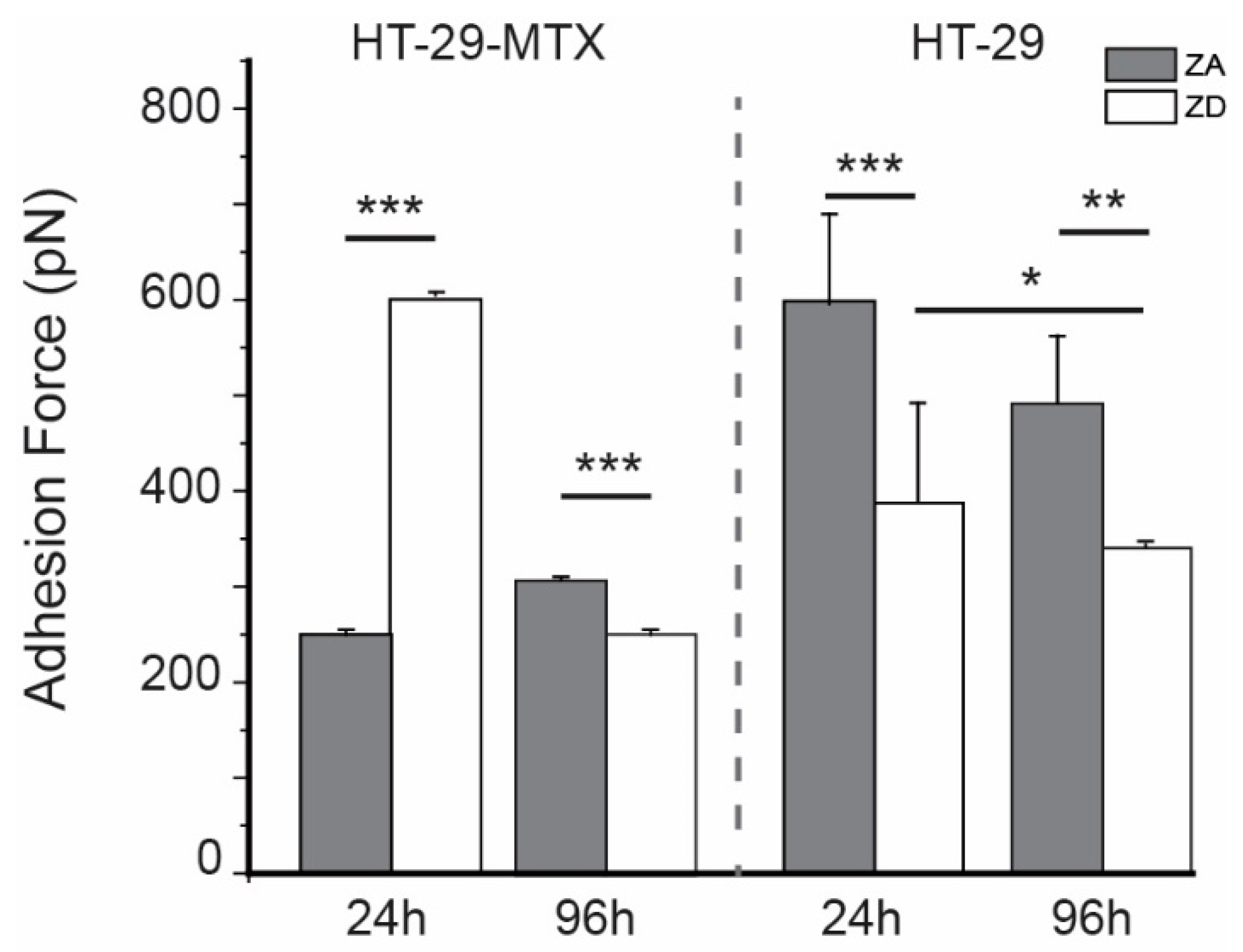
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| 24 h | 0.29 ± 0.01 nN | 0.59 ± 0.03 nN | 0.71 ± 0.24 nN | 0.38 ± 0.09 nN |
| 96 h | 0.31 ± 0.02 nN | 0.24 ± 0.01 nN | 0.60 ± 0.07 nN | 0.33 ± 0.02 nN |
| Incubation | HT-29-MTX | HT-29 | ||
|---|---|---|---|---|
| ZA | ZD | ZA | ZD | |
| Mean Event Position | ||||
| 24 h | 6.86 ± 0.17 µm | 3.48 ± 0.14 µm | 6.49 ± 0.24 µm | 4.33 ± 0.23 µm |
| 96 h | 3.23 ± 0.15 µm | 2.69 ± 0.89 µm | 4.76 ± 0.31 µm | 2.28 ± 0.17 µm |
| Mean Rupture Force | ||||
| 24 h | 56.9 ± 1.23 pN | 55.9 ± 0.76 pN | 63.8 ± 1.23 pN | 50.2 ± 1.01 pN |
| 96 h | 60.3 ± 0.83 pN | 58.8 ± 0.89 pN | 70.9 ± 1.49 pN | 54.3 ± 1.66 pN |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maares, M.; Keil, C.; Löher, L.; Weber, A.; Andorfer-Sarr, A.; Haase, H.; Iturri, J.; Toca-Herrera, J.L. Time- and Zinc-Related Changes in Biomechanical Properties of Human Colorectal Cancer Cells Examined by Atomic Force Microscopy. Biology 2020, 9, 468. https://doi.org/10.3390/biology9120468
Maares M, Keil C, Löher L, Weber A, Andorfer-Sarr A, Haase H, Iturri J, Toca-Herrera JL. Time- and Zinc-Related Changes in Biomechanical Properties of Human Colorectal Cancer Cells Examined by Atomic Force Microscopy. Biology. 2020; 9(12):468. https://doi.org/10.3390/biology9120468
Chicago/Turabian StyleMaares, Maria, Claudia Keil, Leif Löher, Andreas Weber, Amsatou Andorfer-Sarr, Hajo Haase, Jagoba Iturri, and José L. Toca-Herrera. 2020. "Time- and Zinc-Related Changes in Biomechanical Properties of Human Colorectal Cancer Cells Examined by Atomic Force Microscopy" Biology 9, no. 12: 468. https://doi.org/10.3390/biology9120468
APA StyleMaares, M., Keil, C., Löher, L., Weber, A., Andorfer-Sarr, A., Haase, H., Iturri, J., & Toca-Herrera, J. L. (2020). Time- and Zinc-Related Changes in Biomechanical Properties of Human Colorectal Cancer Cells Examined by Atomic Force Microscopy. Biology, 9(12), 468. https://doi.org/10.3390/biology9120468