From One Ejaculate to Another: Transference of Sperm Traits via Seminal Plasma Supplementation in the Ram




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Methods
2.1.1. Animals and Semen Collection
2.1.2. Assessment of Sperm Kinematic Traits
2.1.3. Seminal Plasma Isolation
2.2. Determining Seminal Plasma Concentration for Inter-Ejaculate Supplementation
2.3. Inter-Ejaculate Seminal Plasma Supplementation
2.3.1. Donor Seminal Plasma Collection
2.3.2. Ejaculate Level Effects of Seminal Plasma Supplementation
2.4. Statistical Analysis
3. Results
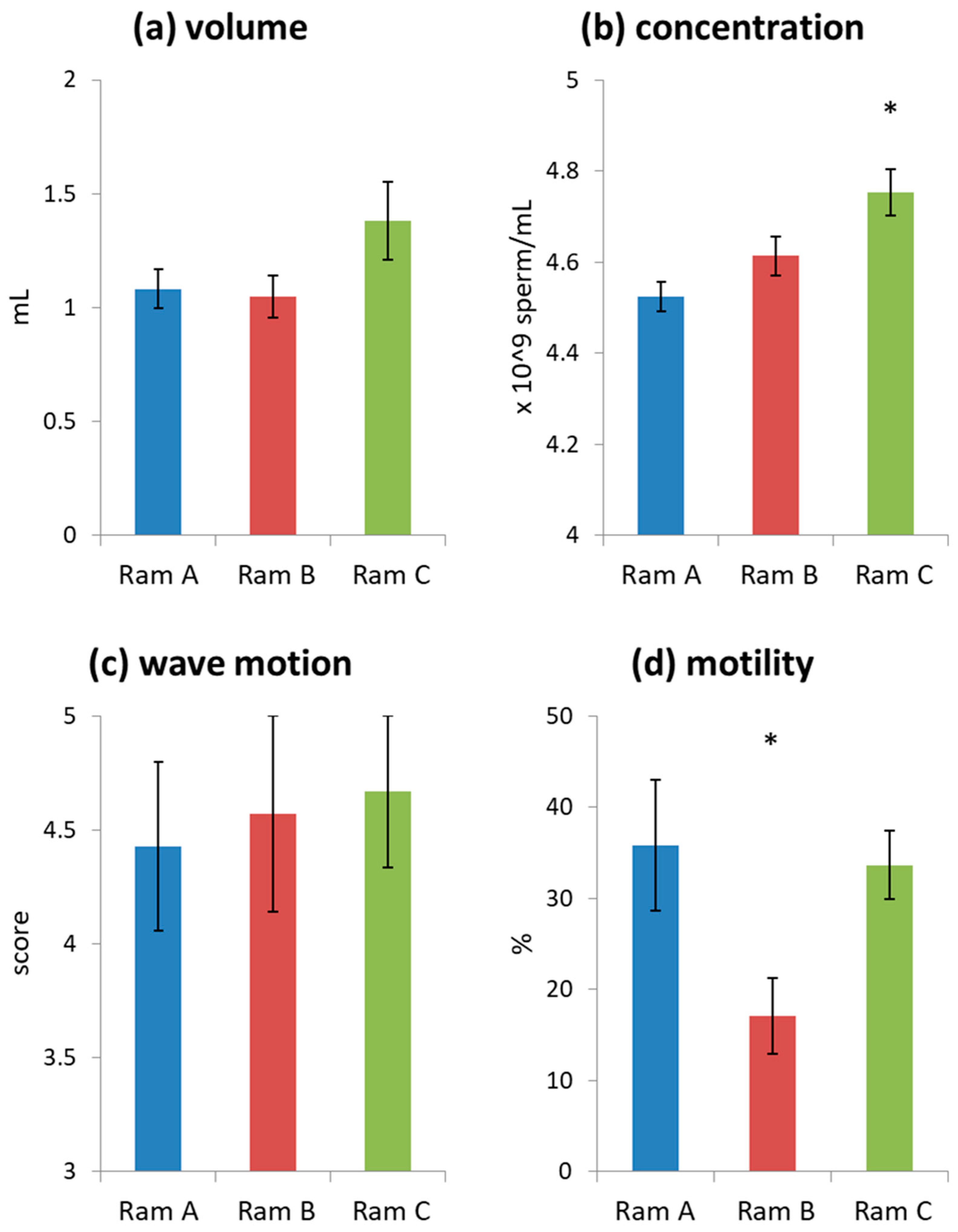
3.1. Ejaculate Characteristics
3.2. Ejaculate Level Effects of Seminal Plasma Supplementation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parker, G.A.; Pizzari, T. Sperm competition and ejaculate economics. Biol. Rev. 2010, 85, 897–934. [Google Scholar] [CrossRef]
- Kilgallon, S.J.; Simmons, L.W. Image content influences men’s semen quality. Biol. Lett. 2005, 1, 253–255. [Google Scholar] [CrossRef] [Green Version]
- Joseph, P.N.; Sharma, R.K.; Agarwal, A.; Sirot, L.K. Men ejaculate larger volumes of semen, more motile sperm, and more quickly when exposed to images of novel women. Evol. Psychol. Sci. 2015, 1, 195–200. [Google Scholar] [CrossRef]
- Hess, R.A.; de Franca, L.R. Spermatogenesis and cycle of the seminiferous epithelium. In Molecular Mechanisms in Spermatogenesis; Cheng, C.Y., Ed.; Springer: New York, NY, USA, 2008; pp. 1–15. [Google Scholar] [CrossRef]
- Franca, L.R.; Avelar, G.F.; Almeida, F.F. Spermatogenesis and sperm transit through the epididymis in mammals with emphasis on pigs. Theriogenology 2005, 63, 300–318. [Google Scholar] [CrossRef]
- Baker, M.A. The ‘omics revolution and our understanding of sperm cell biology. Asian J. 2011, 13, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.C.; Sirot, L.; Wigby, S. The seminal symphony: How to compose an ejaculate. Trends Ecol. Evol. 2013, 28, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Bromfield, J.J. Seminal fluid and reproduction: Much more than previously thought. J. Assist. Reprod Gen. 2014, 31, 627–636. [Google Scholar] [CrossRef] [Green Version]
- Crean, A.J.; Adler, M.I.; Bonduriansky, R. Seminal fluid and mate choice: New predictions. Trends Ecol. Evol. 2016, 31, 253–255. [Google Scholar] [CrossRef]
- Leahy, T.; Rickard, J.P.; Bernecic, N.C.; Druart, X.; de Graaf, S.P. Ram seminal plasma and its functional proteomic assessment. Reproduction 2019, 157, R243–R256. [Google Scholar] [CrossRef]
- Druart, X.; Rickard, J.P.; Mactier, S.; Kohnke, P.L.; Kershaw-Young, C.M.; Bathgate, R.; Gibb, Z.; Crossett, B.; Tsikis, G.; Labas, V.; et al. Proteomic characterization and cross species comparison of mammalian seminal plasma. J. Proteom. 2013, 91, 13–22. [Google Scholar] [CrossRef]
- Leahy, T.; Marti, J.I.; Evans, G.; Maxwell, W.M.C. Seasonal variation in the protective effect of seminal plasma on frozen-thawed ram spermatozoa. Anim. Reprod. Sci. 2010, 119, 147–153. [Google Scholar] [CrossRef]
- Rickard, J.P.; Pini, T.; Soleilhavoup, C.; Cognie, J.; Bathgate, R.; Lynch, G.W.; Evans, G.; Maxwell, W.M.; Druart, X.; de Graaf, S.P. Seminal plasma aids the survival and cervical transit of epididymal ram spermatozoa. Reproduction 2014, 148, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Bedford, J.M. The functions--or not--of seminal plasma? Biol. Reprod. 2015, 92, 18. [Google Scholar] [CrossRef]
- Kanwar, K.C.; Yanagimachi, R.; Lopata, A. Effects of human seminal plasma on fertilizing capacity of human spermatozoa. Fertil. Steril. 1979, 31, 321–327. [Google Scholar] [CrossRef]
- Leahy, T.; De Graaf, S.P. Seminal plasma and its effect on ruminant spermatozoa during processing. Reprod. Domest. Anim. 2012, 47, 207–213. [Google Scholar] [CrossRef]
- Rickard, J.P.; Schmidt, R.E.; Maddison, J.W.; Bathgate, R.; Lynch, G.W.; Druart, X.; De Graaf, S.P. Variation in seminal plasma alters the ability of ram spermatozoa to survive cryopreservation. Reprod. Fert. Dev. 2016, 28, 516–523. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Gago, R.; Alvarez-Rodriguez, M.; Alonso, M.E.; Gonzalez, J.R.; Alegre, B.; Dominguez, J.C.; Martinez-Pastor, F. Thawing boar semen in the presence of seminal plasma improves motility, modifies subpopulation patterns and reduces chromatin alterations. Reprod. Fert. Dev. 2017, 29, 1576–1584. [Google Scholar] [CrossRef]
- Morrell, J.M.; Georgakas, A.; Lundeheim, N.; Nash, D.; Davies Morel, M.C.; Johannisson, A. Effect of heterologous and homologous seminal plasma on stallion sperm quality. Theriogenology 2014, 82, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Pizzari, T. Sperm competition: The need for speed. Elife 2017, 6. [Google Scholar] [CrossRef]
- Rickard, J.P.; Leahy, T.; Soleilhavoup, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Lynch, G.W.; Druart, X.; de Graaf, S.P. The identification of proteomic markers of sperm freezing resilience in ram seminal plasma. J. Proteom. 2015, 126, 303–311. [Google Scholar] [CrossRef]
- Soleilhavoup, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Kohnke, P.L.; Dacheux, J.L.; Guerin, Y.; Gatti, J.L.; De Graaf, S.P.; Druart, X. Ram seminal plasma proteome and its impact on liquid preservation of spermatozoa. J. Proteom. 2014, 109, 245–260. [Google Scholar] [CrossRef]
- Qu, F.; Ying, X.; Guo, W.; Guo, Q.; Chen, G.; Liu, Y.; Ding, Z. The role of Zn-alpha2 glycoprotein in sperm motility is mediated by changes in cyclic AMP. Reproduction 2007, 134, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brokaw, C.J. Regulation of sperm flagellar motility by calcium and cAMP-dependent phosphorylation. J. Cell Biochem. 1987, 35, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, R. Artificial insemination: The state of the art. Theriogenology 2003, 59, 571–584. [Google Scholar] [CrossRef]
- Moore, S.G.; Hasler, J.F. A 100-Year Review: Reproductive technologies in dairy science. J. Dairy Sci. 2017, 100, 10314–10331. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Green, C.; Rickard, J.P.; de Graaf, S.P.; Crean, A.J. From One Ejaculate to Another: Transference of Sperm Traits via Seminal Plasma Supplementation in the Ram. Biology 2020, 9, 33. https://doi.org/10.3390/biology9020033
Green C, Rickard JP, de Graaf SP, Crean AJ. From One Ejaculate to Another: Transference of Sperm Traits via Seminal Plasma Supplementation in the Ram. Biology. 2020; 9(2):33. https://doi.org/10.3390/biology9020033
Chicago/Turabian StyleGreen, Christine, Jessica P. Rickard, Simon P. de Graaf, and Angela J. Crean. 2020. "From One Ejaculate to Another: Transference of Sperm Traits via Seminal Plasma Supplementation in the Ram" Biology 9, no. 2: 33. https://doi.org/10.3390/biology9020033
APA StyleGreen, C., Rickard, J. P., de Graaf, S. P., & Crean, A. J. (2020). From One Ejaculate to Another: Transference of Sperm Traits via Seminal Plasma Supplementation in the Ram. Biology, 9(2), 33. https://doi.org/10.3390/biology9020033