Morphological Features of the Anther Development in Tomato Plants with Non-Specific Male Sterility

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
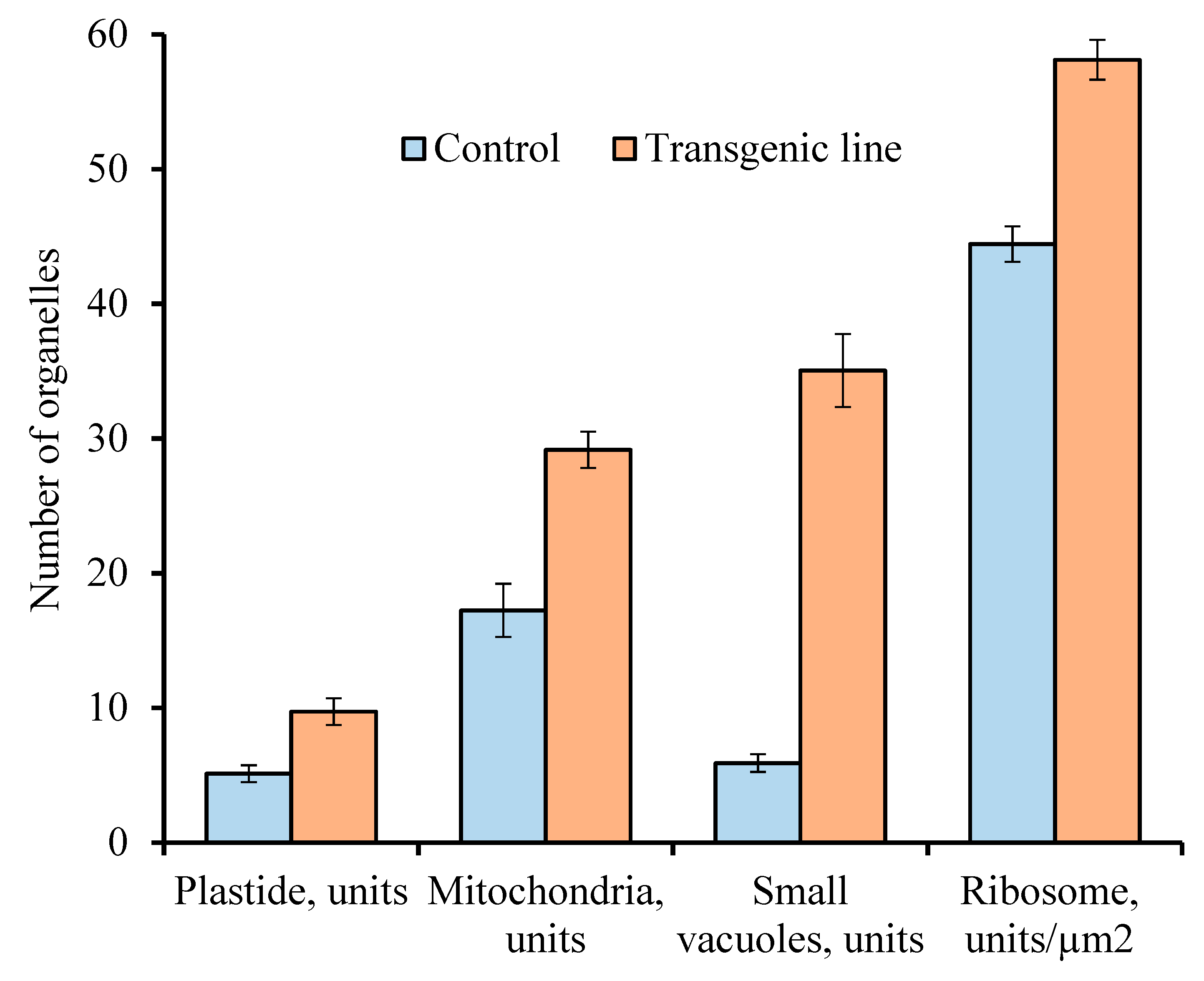{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
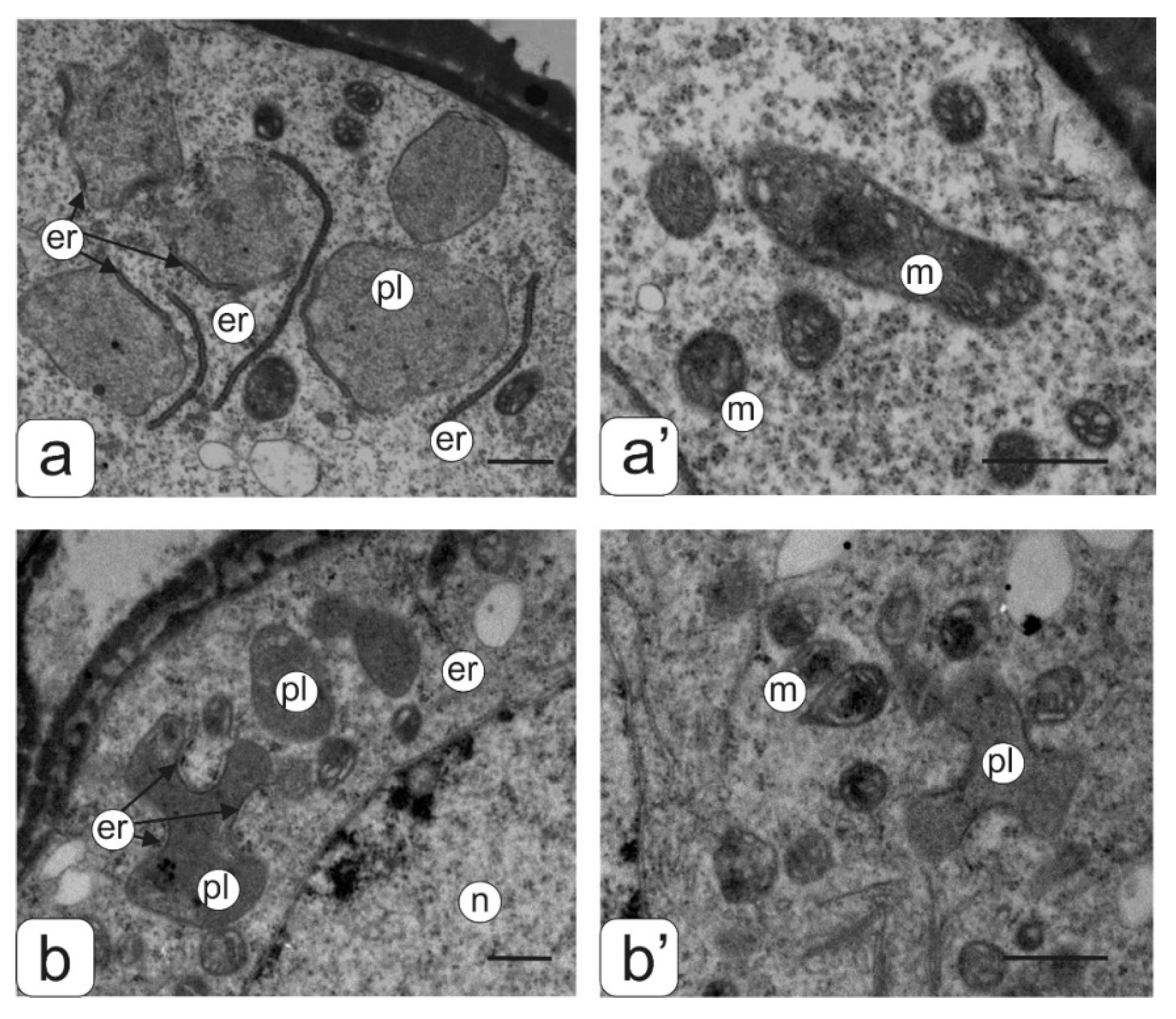
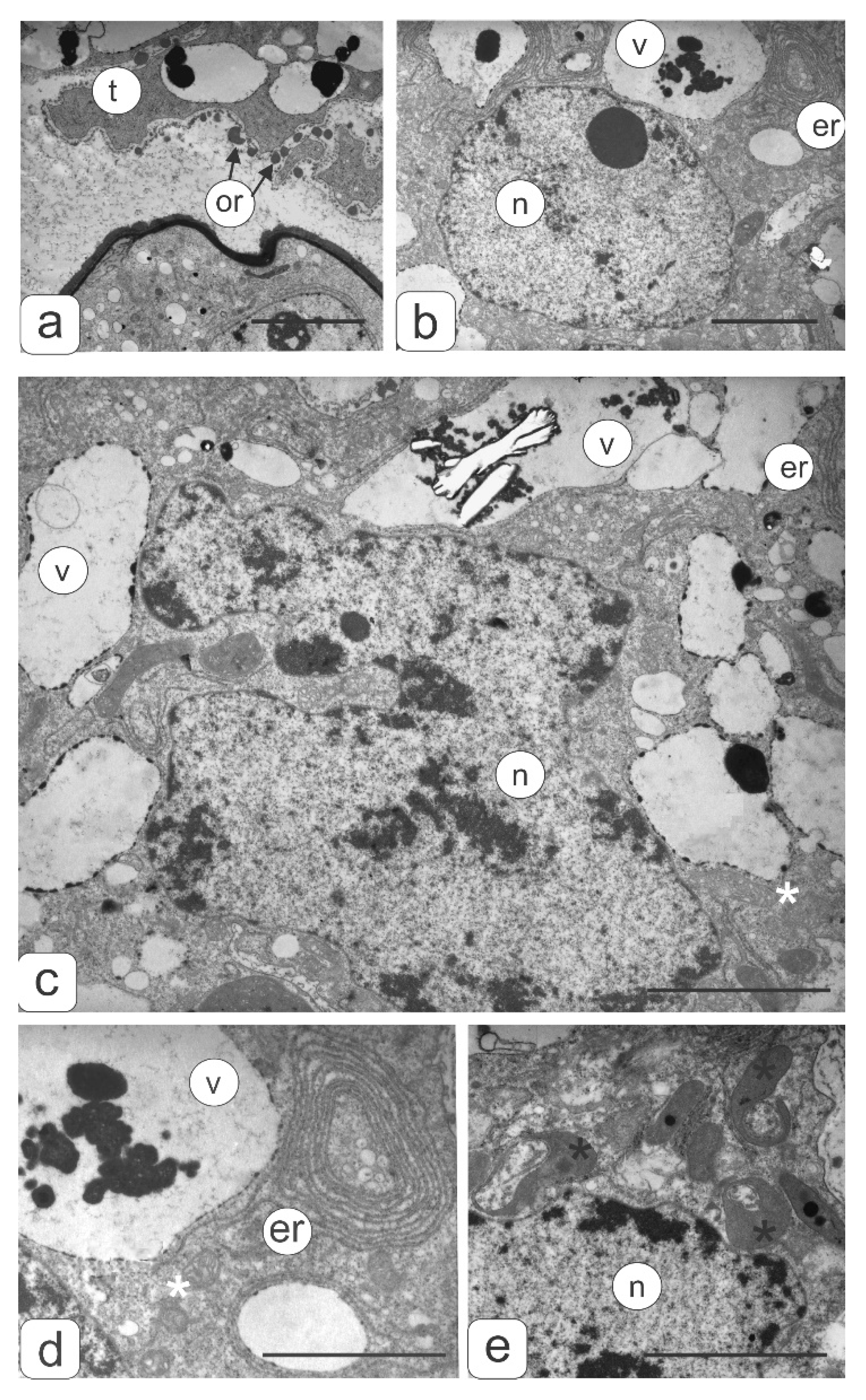
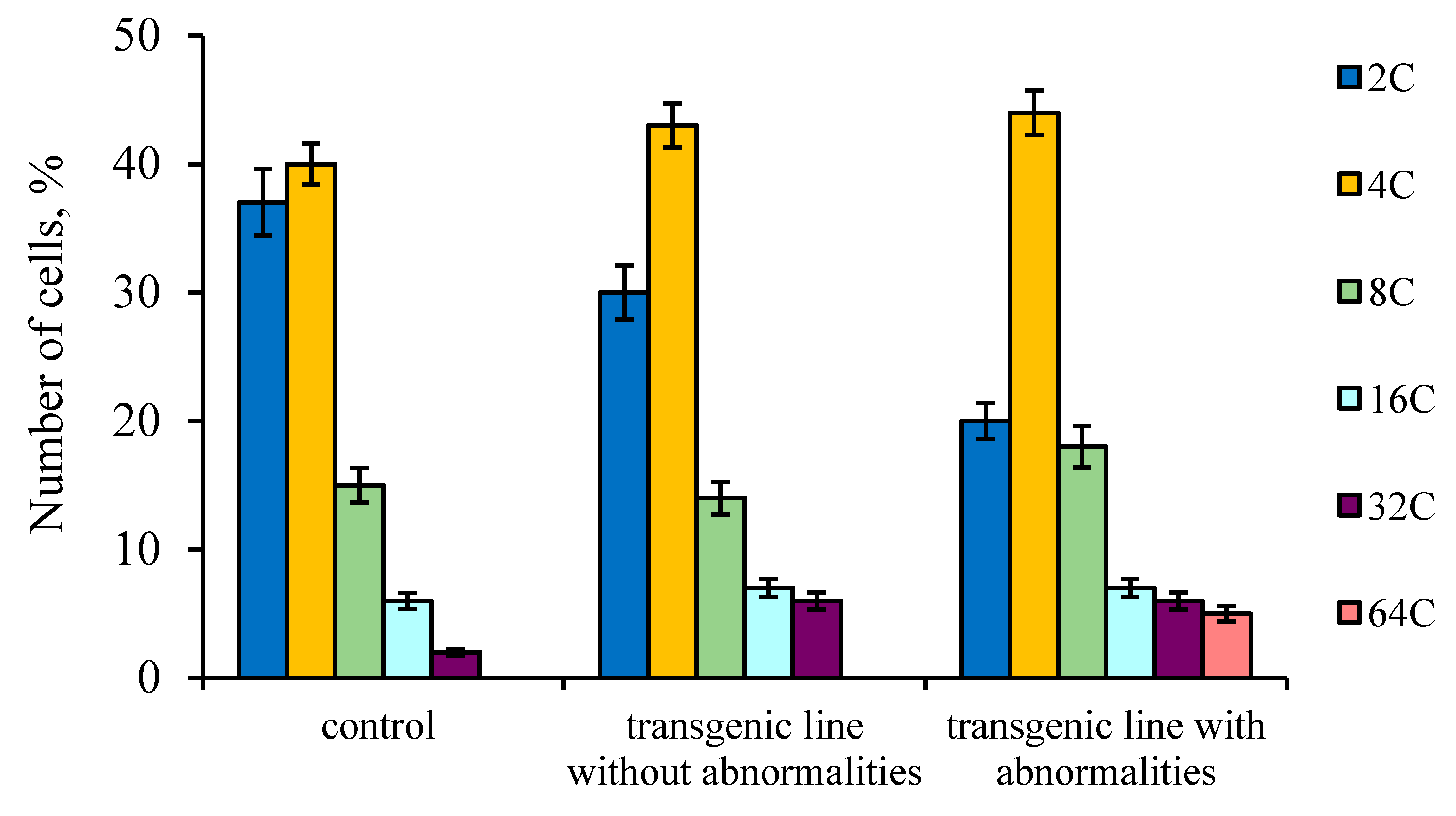
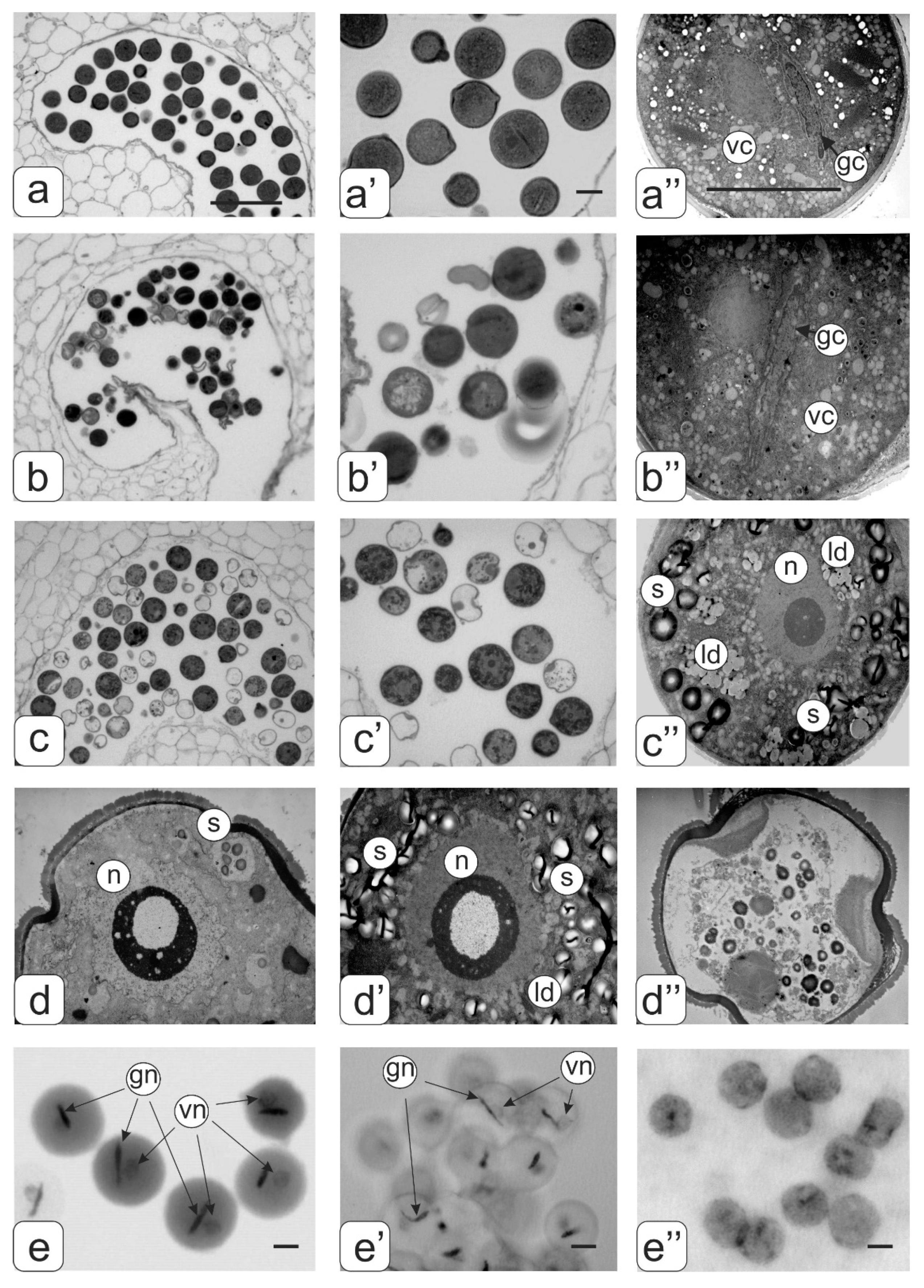
2. Results
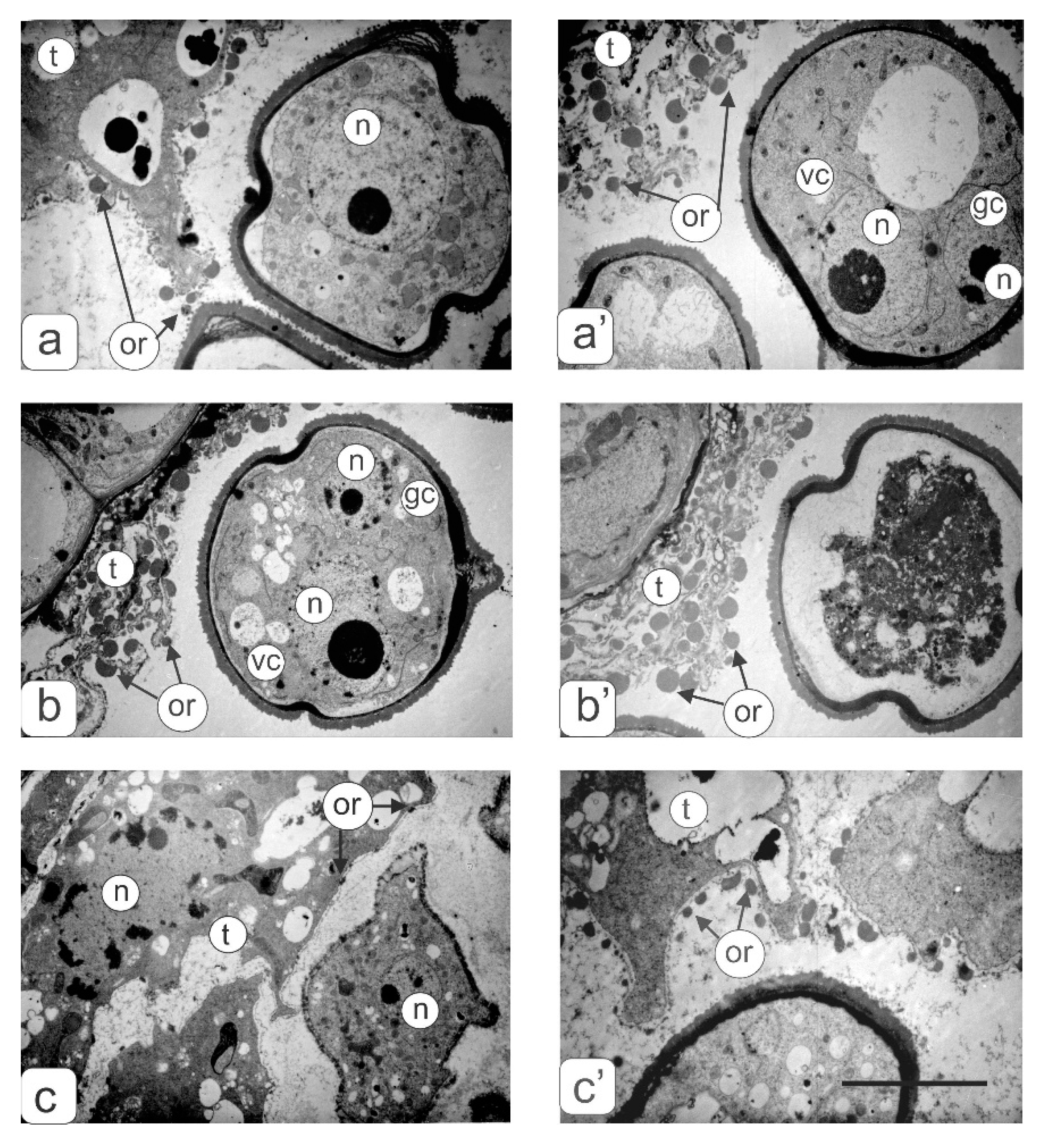
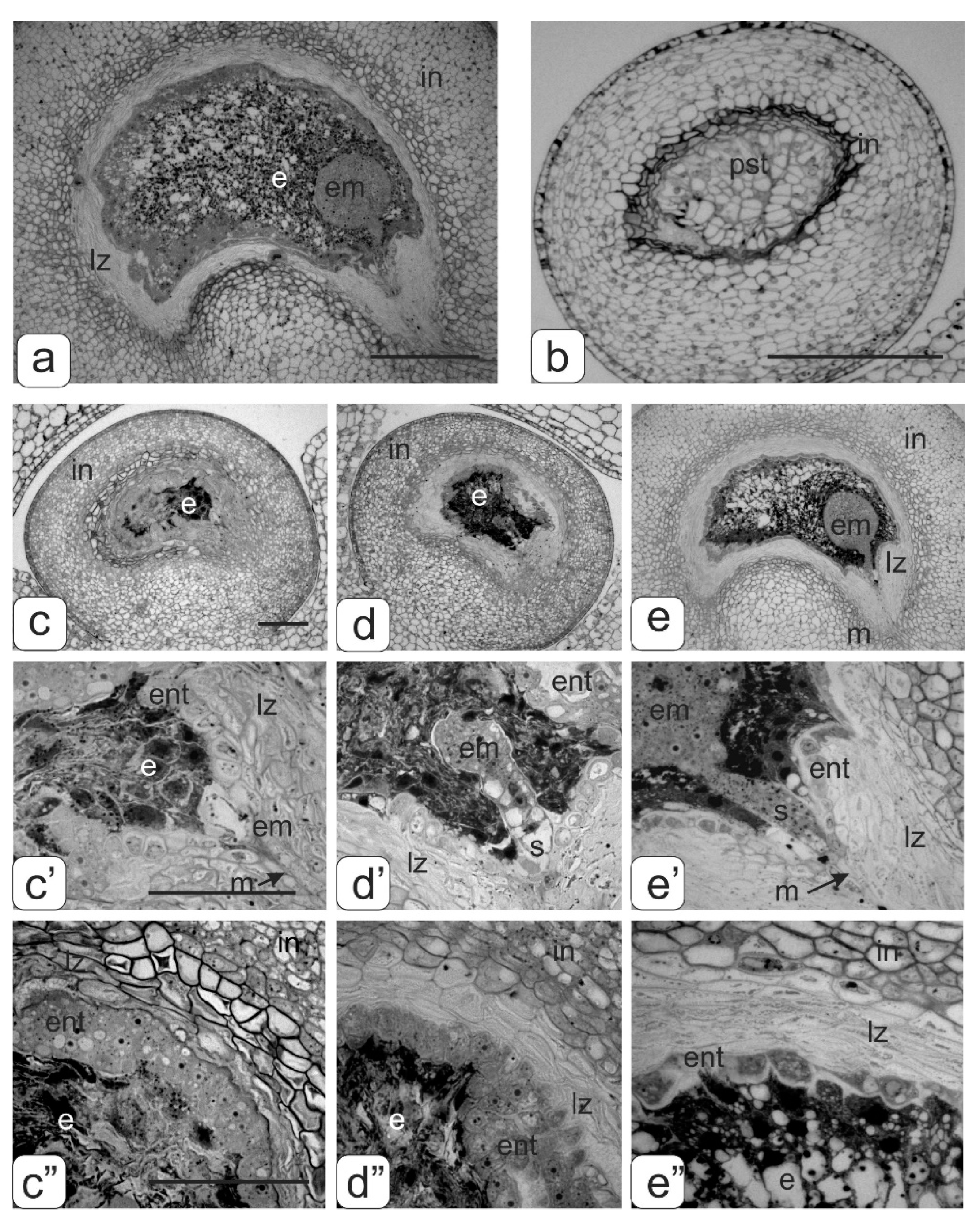
- underdevelopment or complete atrophy of one or more pollen sacs (Figure 4a);
- merging of two pollen sacs into one common sac and degradation of the other two (Figure 4b);
- coalescence of two stamens along the entire length and atrophy of pollen sacs in these merged anthers (Figure 4c);
- integration of two stamens into one with complete atrophy of two pollen sacs from each of them as evidenced by two vascular bundles (Figure 4d).
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Light Microscopy and Transmission Electron Microscopy
4.3. Cytophotometry and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chandler, J.W. The hormonal regulation of flower development. J. Plant Growth Regul. 2011, 30, 242–254. [Google Scholar] [CrossRef]
- Wilson, Z.A.; Zhang, D.B. From Arabidopsis to rice: Pathways in pollen development. J. Exp. Bot. 2009, 60, 1479–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Liu, Y.G. Male sterility and fertility restoration in crops. Ann. Rev. Plant Biol. 2014, 65, 570–606. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Kang, J.H.; Zhao, M.; Kwon, J.K.; Choi, H.S.; Bae, J.H.; Lee, H.A.; Joung, Y.H.; Choi, D.; Kang, B.C. Tomato Male sterile 1035 is essential for pollen development and meiosis in anthers. J. Exp. Bot. 2014, 65, 6693–6709. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.; Hao, M.; Mei, D.; Zaman, Q.U.; Sang, S.; Wang, H.; Wang, W.; Fu, L.; Cheng, H.; Hu, Q. Transcriptome and hormone comparison of three cytoplasmic male sterile systems in Brassica napus. Int. J. Mol. Sci. 2018, 19, 4022. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.L.; Xie, L.F.; Mao, H.Z.; Puah, C.S.; Yang, W.C.; Jiang, L.; Sundaresan, V.; Ye, D. Tapetum determinant1 is required for cell specialization in the Arabidopsis anther. Plant Cell 2003, 15, 2792–2804. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.H.; Han, M.J.; Lee, Y.S.; Kim, Y.W.; Hwang, I.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; An, G. Rice Undeveloped Tapetum1 is a major regulator of early tapetum development. Plant Cell 2005, 17, 2705–2722. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zhou, C.; Yang, J. Molecular phenotypes associated with anomalous stamen development in Alternanthera philoxeroides. Front. Plant Sci. 2015, 6, 6–242. [Google Scholar] [CrossRef] [Green Version]
- Lozano, R.; Gimenez, E.; Cara, B.; Capel, J.; Angosto, T. Genetic analysis of reproductive development in tomato. Int. J. Dev. Biol. 2009, 53, 1635–1648. [Google Scholar] [CrossRef]
- Tchórzewska, D.; Deryło, K.; Błaszczyk, L.; Winiarczyk, K. Tubulin cytoskeleton during microsporogenesis in the male-sterile genotype of Allium sativum and fertile Allium ampeloprasum L. Plant Reprod. 2015, 28, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Tchórzewska, D.; Deryło, K.; Winiarczyk, K. Cytological and biophysical comparative analysis of cell structures at the microsporogenesis stage in sterile and fertile Allium species. Planta 2017, 245, 137–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchórzewska, D.; Luchowski, R.; Gruszecki, W.I.; Winiarczyk, K. Comparative studies of live tapetum cells in sterile garlic (Allium sativum) and fertile leek (Allium ampeloprasum) using the fluorescence lifetime imaging analytical method. S. Afr. J. Bot. 2018, 117, 222–231. [Google Scholar] [CrossRef]
- Bhandari, N.N. The microsporangium. In Embryology of Angiosperms; Johri, B.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 53–121. [Google Scholar]
- Khaliluev, M.R.; Chaban, I.A.; Kononenko, N.V.; Baranova, E.N.; Dolgov, S.V.; Kharchenko, P.N.; Polyakov, V.Y. Abnormal floral meristem development in transgenic tomato plants do not depend on the expression of genes encoding defense-related PR-proteins and antimicrobial peptides. Russ. J. Dev. Biol. 2014, 45, 22–33. [Google Scholar] [CrossRef]
- Chaban, I.; Khaliluev, M.; Baranova, E.; Kononenko, N.; Dolgov, S.; Smirnova, E. Abnormal development of floral meristem triggers defective morphogenesis of generative system in transgenic tomatoes. Protoplasma 2018, 255, 1597–1611. [Google Scholar] [CrossRef] [PubMed]
- Jullien, P.E.; Berger, F. Parental genome dosage imbalance deregulates imprinting in Arabidopsis. PLoS Genet. 2010, 6, e1000885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilkes, B.P.; Spielman, M.; Weizbauer, R.; Watson, B.; Burkart-Waco, D.; Scott, R.J.; Comai, L. The maternally expressed WRKY transcription factor TTG2 controls lethality in interploidy crosses of Arabidopsis. PLoS Biol. 2008, 6, e308. [Google Scholar] [CrossRef] [Green Version]
- Chaban, I.; Baranova, E.; Kononenko, N.; Khaliluev, M.; Smirnova, E. Distinct differentiation characteristics of endothelium determine its ability to form pseudoembryos in tomato ovules. Int. J. Mol. Sci. 2020, 21, 12. [Google Scholar]
- Gorman, S.W.; McCormick, S. Male sterility in tomato. Crit. Rev. Plant Sci. 1997, 16, 31–53. [Google Scholar] [CrossRef]
- Sawhney, V.K.; Bhadula, S.K. Microsporogenesis in the normal and male sterile stamenless-2 mutant of tomato (Lycopersicon esculentum). Can. J. Bot. 1988, 66, 2013–2021. [Google Scholar] [CrossRef]
- Roberts, M.R.; Boyes, E.; Scott, R.J. An investigation of the role of the anther tapetum during microspore development using genetic cell ablation. Sex. Plant Reprod. 1995, 8, 299–307. [Google Scholar] [CrossRef]
- Lei, X.; Liu, B. Tapetum-dependent male meiosis progression in plants: Increasing evidence emerges. Front. Plant Sci. 2019, 10, 1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, M.; Vulkova, Z.; Gorinova, N.; Izhar, S.; Firon, N.; Jacquemin, J.-M.; Atanassov, A.; Stoeva, P. Characterisation of a cytoplasmic male-sterile hybrid line between Lycopersicon peruvianum Mill.× Lycopersicon pennellii Corr. and its crosses with cultivated tomato. Theor. Appl. Gen. 1999, 98, 825–830. [Google Scholar] [CrossRef]
- Stoeva-Popova, P.K.; Dimaculangan, D.; Radkova, M.; Vulkova, Z. Towards cytoplasmic male sterility in cultivated tomato. J. Agric. Food Environ. Sci. 2007, 1, 1–13. [Google Scholar]
- Polowick, P.L.; Sawhney, V.K. An ultrastructural study of pollen development in tomato (Lycopersicon esculentum). 11. Pollen maturation. Can. J. Bot. 1993, 71, 1048–1055. [Google Scholar] [CrossRef]
- Polowick, P.L.; Sawhney, V.K. Ultrastructural changes in the cell wall, nucleus and cytoplasm of pollen mother cells during meiotic prophase in Lycopersicon esculenturm Mill. Protoplasma 1992, 169, 139–147. [Google Scholar] [CrossRef]
- Pacini, E.; Franchi, G.G.; Hesse, M. The tapetum: Its form, function, and possible phylogeny in Embryophyta. Plant Syst. Evol. 1985, 149, 155–185. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, Y.; Timofejeva, L.; Chen, C.; Grossniklaus, U.; Ma, H. Regulation of Arabidopsis tapetum development and function by DYSFUNCTIONAL TAPETUM1 (DYT1) encoding a putative bHLH transcription factor. Development 2006, 133, 3085–3095. [Google Scholar] [CrossRef] [Green Version]
- Papini, A.; Mosti, S.; Brighigna, L. Programmed-cell-death events during tapetum development of angiosperms. Protoplasma 1999, 207, 213–221. [Google Scholar] [CrossRef]
- Li, N.; Zhang, D.S.; Liu, H.S.; Yin, C.S.; Li, X.X.; Liang, W.Q.; Yuan, Z.; Xu, B.; Chu, H.-W.; Wang, J.; et al. The Rice Tapetum Degeneration Retardation gene is required for tapetum degradation and anther development. Plant Cell 2006, 18, 2999–3014. [Google Scholar] [CrossRef] [Green Version]
- Kawanabe, T.; Ariizumi, T.; Kawai-Yamada, M.; Uchimiya, H.; Toriyama, K. Abolition of the tapetum suicide program ruins microsporogenesis. Plant Cell Physiol. 2006, 47, 784–787. [Google Scholar] [CrossRef] [Green Version]
- Flawell, R. A model fo the mechanism of cytoplasmic male sterility in plants with special reference to maize. Plant Sci. Lett. 1974, 3, 259–263. [Google Scholar]
- Horn, R.; Gupta, K.J.; Colombo, N. Mitochondrion role in molecular basis of cytoplasmic male sterility. Mitochondrion 2014, 19, 198–205. [Google Scholar] [CrossRef]
- Zhang, D.; Luo, X.; Zhu, L. Cytological analysis and genetic control of rice anther development. J. Genetics Genomics 2011, 38, 379–390. [Google Scholar] [CrossRef]
- Batygina, T.B.; Vasilyeva, V.E. Periodization in the development of flowering plant reproductive structures: Critical periods. Acta Biol. Crac. Ser. Bot. 2003, 45, 27–36. [Google Scholar]
- Roth, M.; Florez-Rueda, A.M.; Griesser, S.; Paris, M.; Städler, T. Incidence and developmental timing of endosperm failure in post-zygotic isolation between wild tomato lineages. Ann. Bot. 2018, 121, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, C.; Baulcombe, D.C.; Chen, Z.J. Maternal siRNAs as regulators of parental genome imbalance and gene expression in endosperm of Arabidopsis seeds. Proc. Natl. Acad. Sci. USA 2012, 109, 5529–5534. [Google Scholar] [CrossRef] [Green Version]
- Li, D.-D.; Xue, J.-S.; Zhu, J.; Yang, Z.-N. Gene regulatory network for tapetum development in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1559. [Google Scholar] [CrossRef] [Green Version]
- Shivanna, K.R. Pollen Biology and Biotechnology; Science Publishers Inc.: Enfield, UK, 2003; p. 316. [Google Scholar]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaban, I.A.; Kononenko, N.V.; Gulevich, A.A.; Bogoutdinova, L.R.; Khaliluev, M.R.; Baranova, E.N. Morphological Features of the Anther Development in Tomato Plants with Non-Specific Male Sterility. Biology 2020, 9, 32. https://doi.org/10.3390/biology9020032
Chaban IA, Kononenko NV, Gulevich AA, Bogoutdinova LR, Khaliluev MR, Baranova EN. Morphological Features of the Anther Development in Tomato Plants with Non-Specific Male Sterility. Biology. 2020; 9(2):32. https://doi.org/10.3390/biology9020032
Chicago/Turabian StyleChaban, Inna A., Neonila V. Kononenko, Alexander A. Gulevich, Liliya R. Bogoutdinova, Marat R. Khaliluev, and Ekaterina N. Baranova. 2020. "Morphological Features of the Anther Development in Tomato Plants with Non-Specific Male Sterility" Biology 9, no. 2: 32. https://doi.org/10.3390/biology9020032
APA StyleChaban, I. A., Kononenko, N. V., Gulevich, A. A., Bogoutdinova, L. R., Khaliluev, M. R., & Baranova, E. N. (2020). Morphological Features of the Anther Development in Tomato Plants with Non-Specific Male Sterility. Biology, 9(2), 32. https://doi.org/10.3390/biology9020032