Effects of Fish Predators on the Mass-Related Energetics of a Keystone Freshwater Crustacean



Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predator | Prey Species | Metabolic Response | Source |
|---|---|---|---|
| AQUATIC | |||
| Octopus | Jasus edwardsii (decapod) | NEG 2 | [31] |
| Octopus | Gobius paganellus (fish) | NEG 2 | [32] |
| Seastar | Fissurella limbata (mollusk) | POS 2 | [33] |
| Odonate larvae | Ischnura elegans (odonate) | NO 2,3 | [34] |
| Odonate larvae | Libellula quadrimaculata (odonate) | NO 2,3 | [34] |
| Odonate larvae | Sympetrum sanguineum (odonate) | NO 2,3 | [34] |
| Odonate larvae | Rana temporaria (tadpole) | POS/NEG 2,3,4 | [23] |
| Odonate larvae | Bufo arabicus (tadpole) | NEG/NO 2,3,4 | [35] |
| Fish | Gammarus minus (amphipod) | POS/NEG 5,6 | [13] |
| Fish | Daphnia magna (cladoceran) | NO 2 | [36] |
| Fish | D. magna (cladoceran) | POS 2 | [37] |
| Fish | D. magna (cladoceran) | NO 2 | [38] |
| Fish | D. magna (cladoceran) | NEG 2 | [39] |
| Fish | Panopeus herbstii (decapod) | NO 2 | [40] |
| Fish | Dreissena polymorpha (mollusk) | NEG/NO/POS 2,4 | [41] |
| Fish | Enallagma cyathigerum (odonate) | POS 2 | [42] |
| Fish | E. vesperum (odonate) | POS 2 | [43] |
| Fish | Carassius auratus (fish) | NEG 2,3 | [44] |
| Fish | C. carassius (fish) | NEG 2 | [45] |
| Fish | Fundulus majalis (fish) | POS 2 | [46] |
| Fish | Galaxias maculatus (fish) | NEG/NO 2,7 | [47] |
| Fish | Parabramis pekinensis (fish) | POS 2 | [24] |
| Fish | Pimephales promelas (fish) | POS 2 | [39] |
| Fish | Poecilia reticulata (fish) | NEG 6 | [48] |
| Fish | P. reticulata (fish) | NEG/NO 2,8 | [48] |
| Fish | P. reticulata (fish) | POS 6 | [49] |
| Fish | Pseudorasbora parva (fish) | POS 2 | [50] |
| Fish | Spinibarbus sinensis (fish) | POS 2,3 | [51] |
| Fish | Zacco platypus (fish) | POS 6 | [52] |
| TERRESTRIAL | |||
| Spider | Hasarius adansoni (spider) | POS/NEG 2,4 | [53] |
| Spider | Melanoplus fenurrubrum (grasshopper) | POS 2 | [54] |
| Mantid insect | Argiope keyserlingi (spider) | POS 2 | [55] |
| Pentatomid insect | Leptinotarsa decemlineata (beetle) | POS/NEG/NO 2,8 | [56,57] |
| Pentatomid insect | Manduca sexta (caterpillar) | POS 2,3 | [58] |
| Reptile | Teleiogryllus commodus (cricket) | POS 2 | [14] |
| Rat | Tenebrio molitor (beetle) | NEG 6 | [59,60] |
| Bird | Parus major (bird) | NO 2 | [61] |
| Frightening stimulus | Sylvilagus aquaticus (mammal) | NEG 2 | [62] |
| Mammal | Cervus elaphus (mammal) | POS 2 | [63] |
| Mammal | Erinaceus europaeus (mammal) | POS 2 | [64] |
| Mammal | E. europaeus (mammal) | NEG 2 | [65] |
2. Materials and Methods
2.1. Study System
2.2. Amphipod Collection
2.3. Measuring Food Assimilation Rate in the Laboratory
2.4. Measuring Fat Content
2.5. Estimating Scaling Exponents of Food Assimilation Rate in the Field
2.6. Measuring Growth Rates
2.7. Measuring Rates of Resting Metabolism
2.8. Estimating Energetic Costs of Maintenance, Growth and Reproduction
2.9. Scaling Analyses
3. Results
3.1. Organization of Results
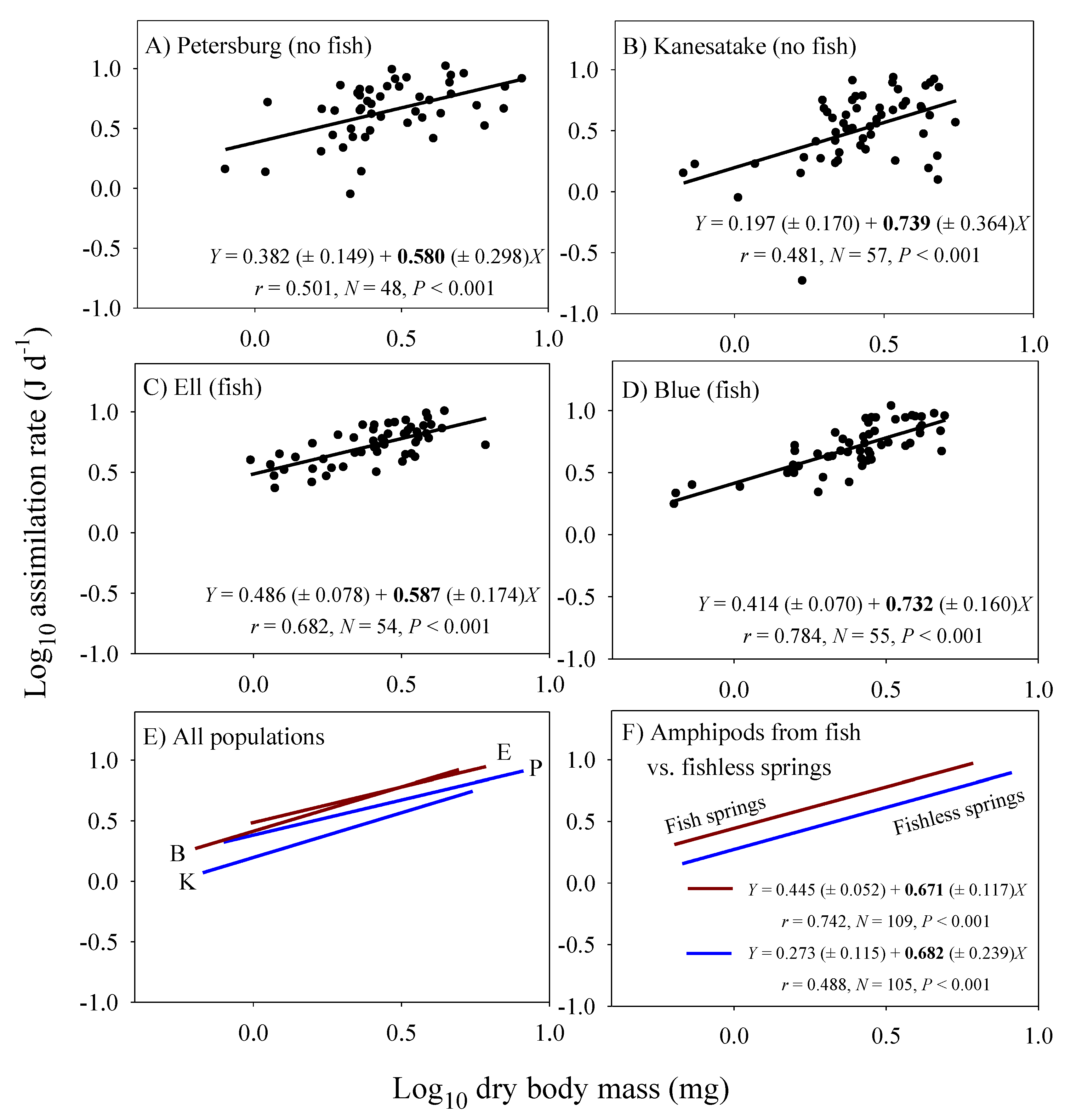
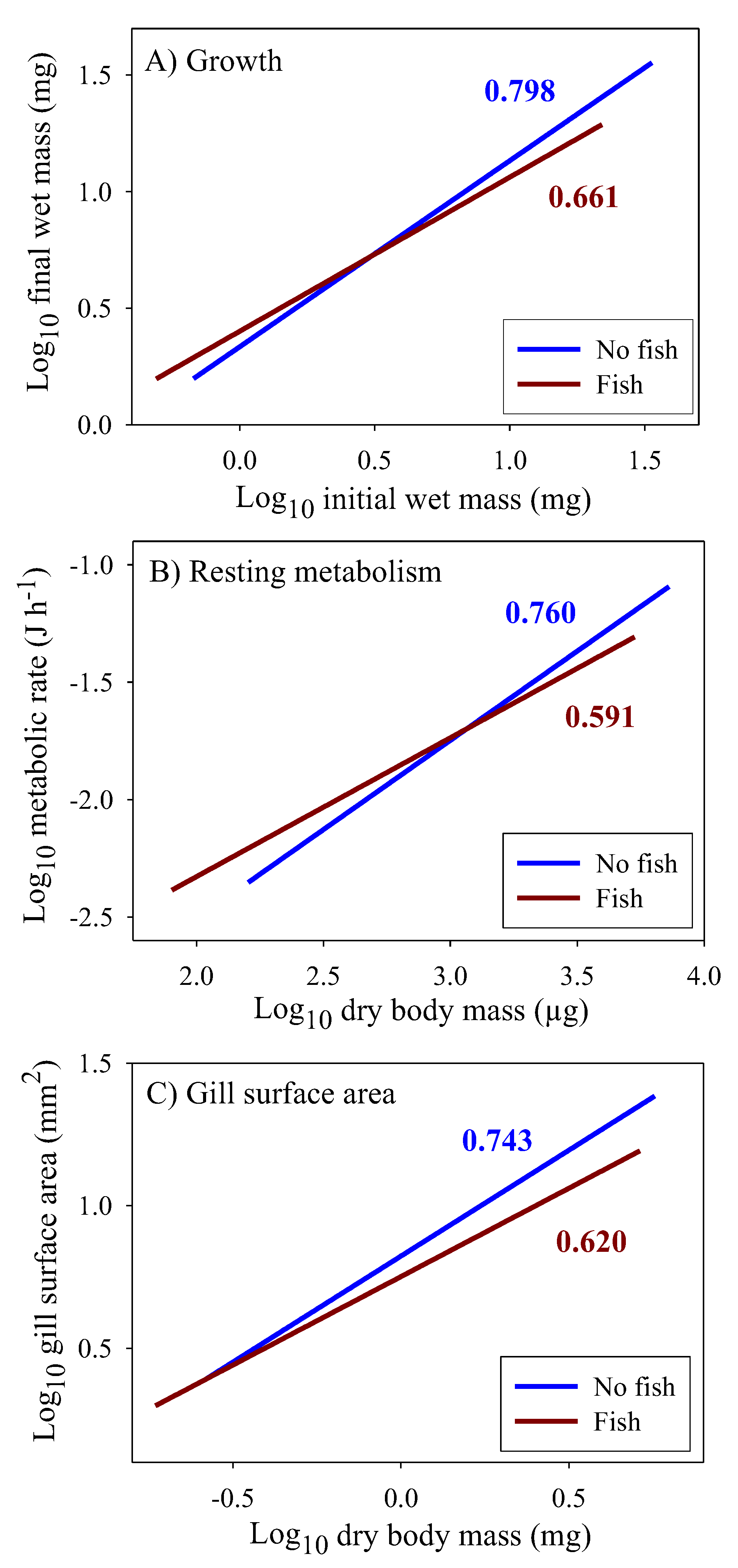
3.2. Scaling of Food Assimilation Rate in the Laboratory
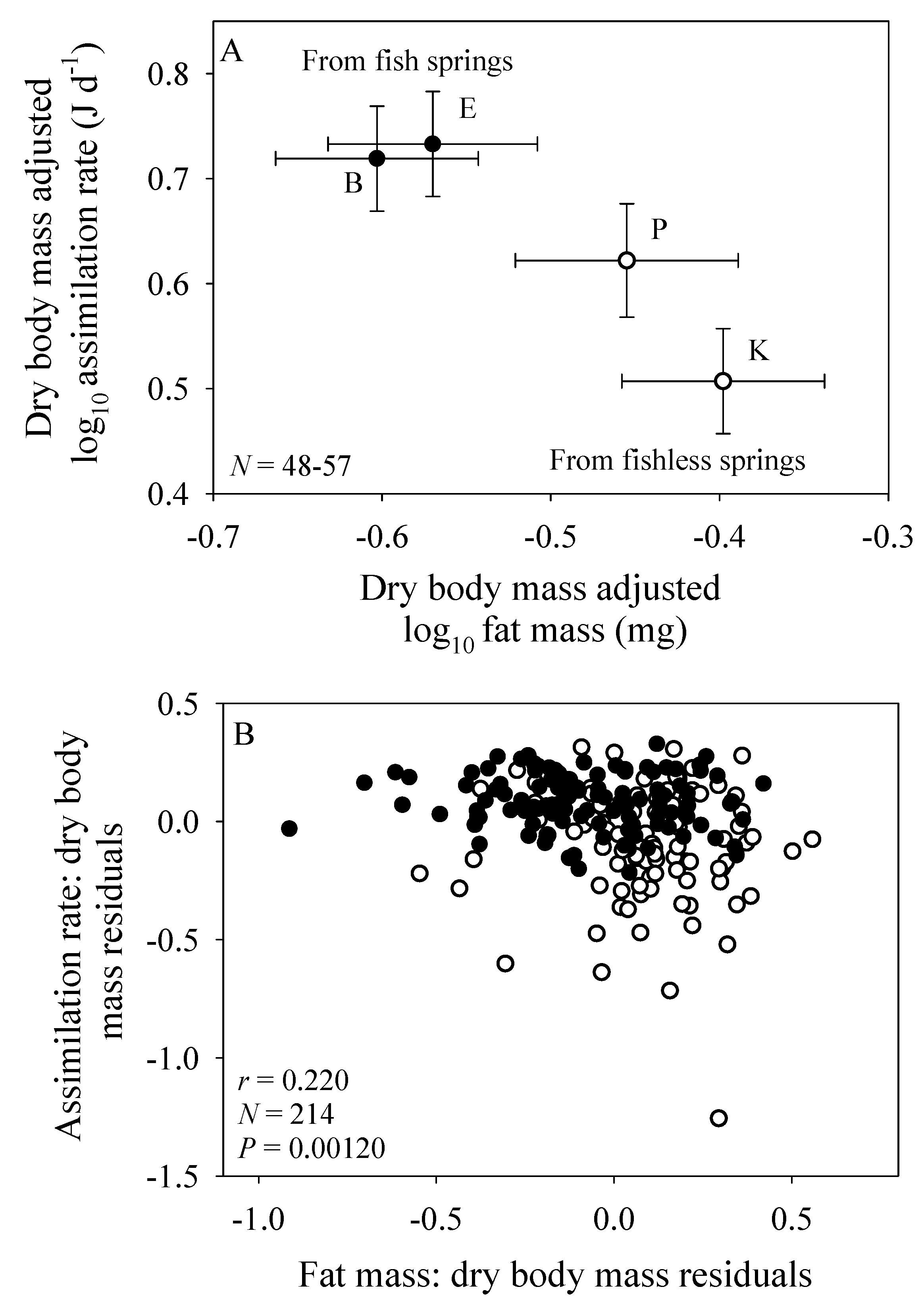
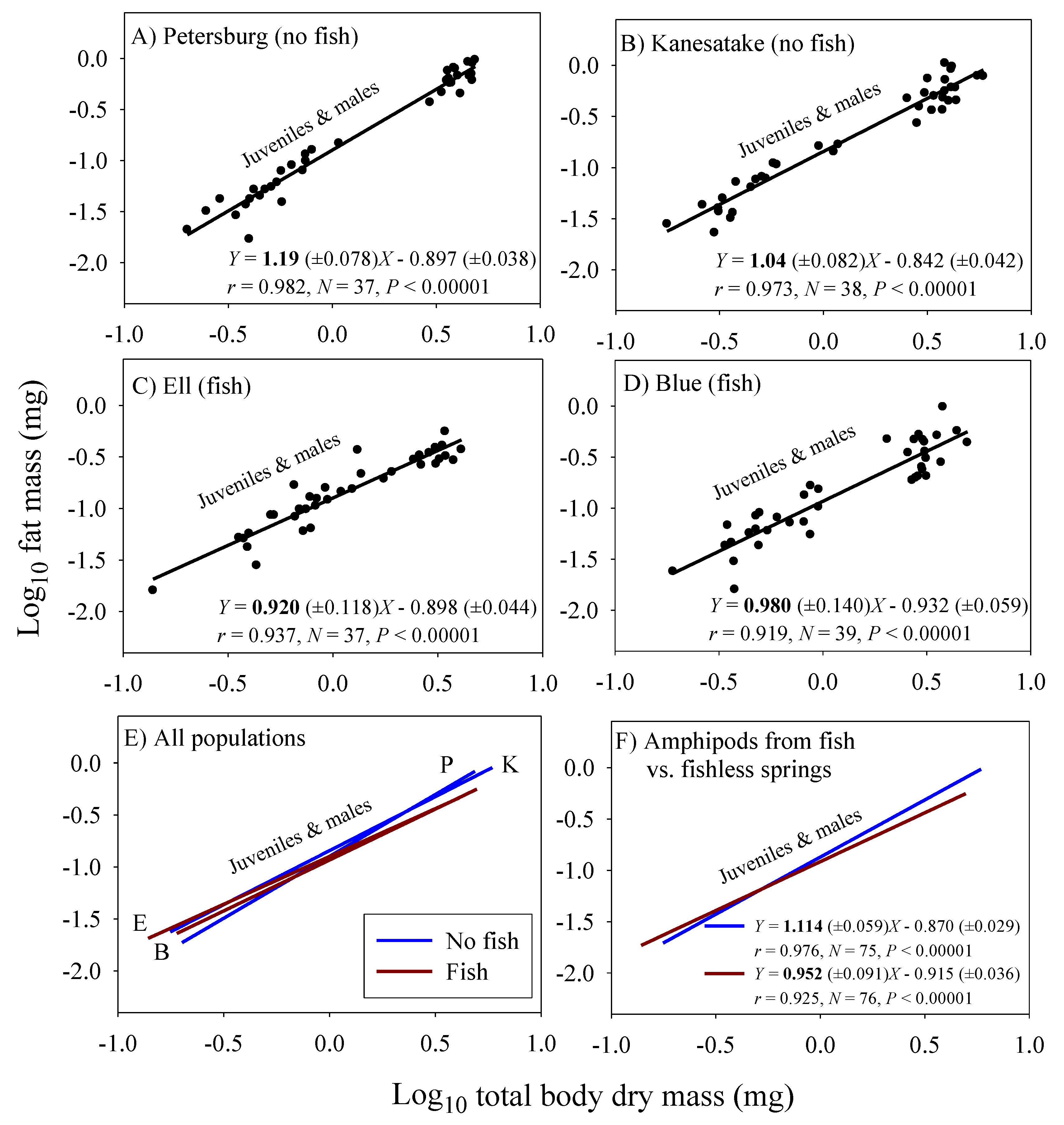
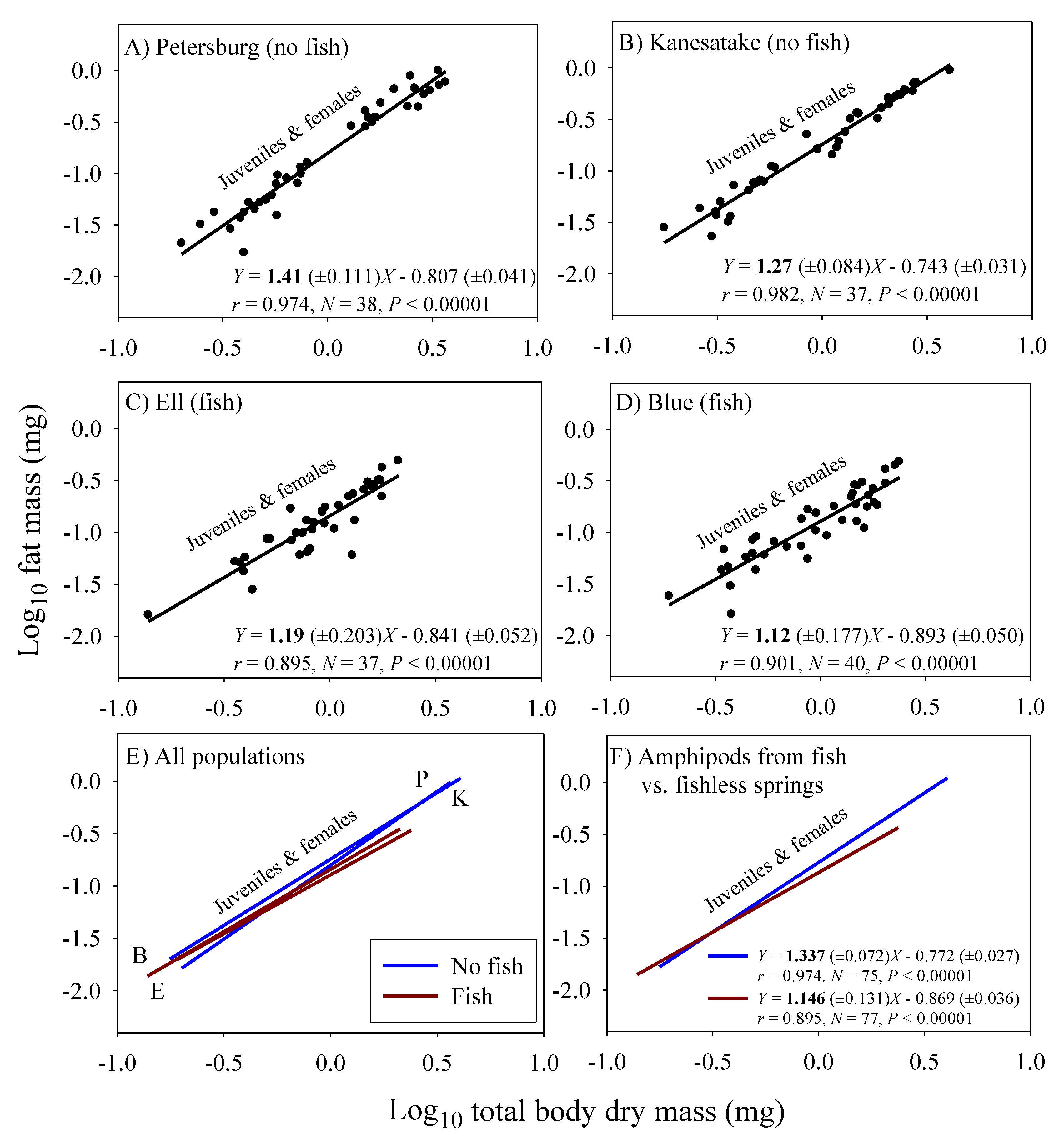
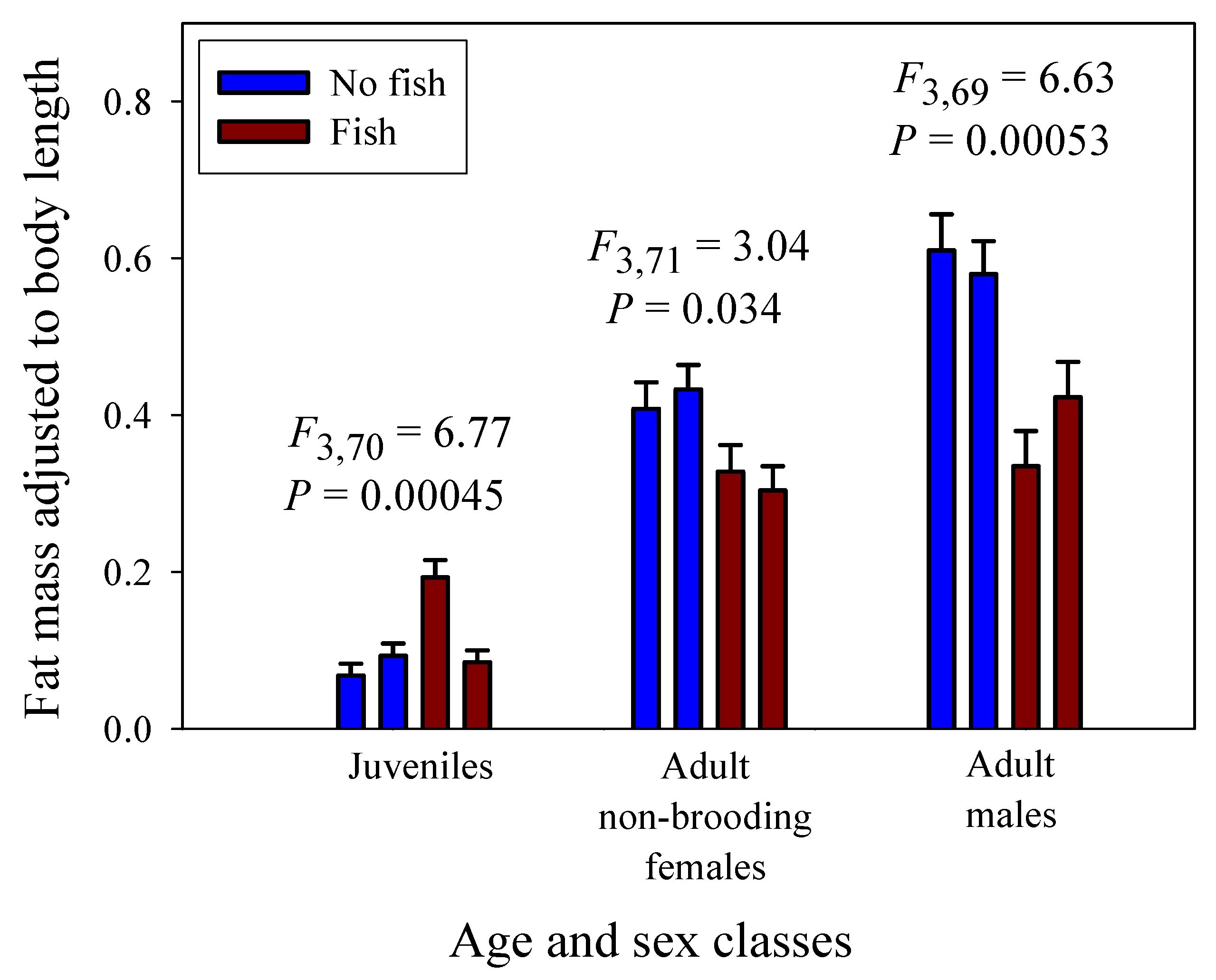
3.3. Scaling of Fat Mass of Field-Collected Amphipods, with Inferences about the Scaling of Feeding Rate in Nature
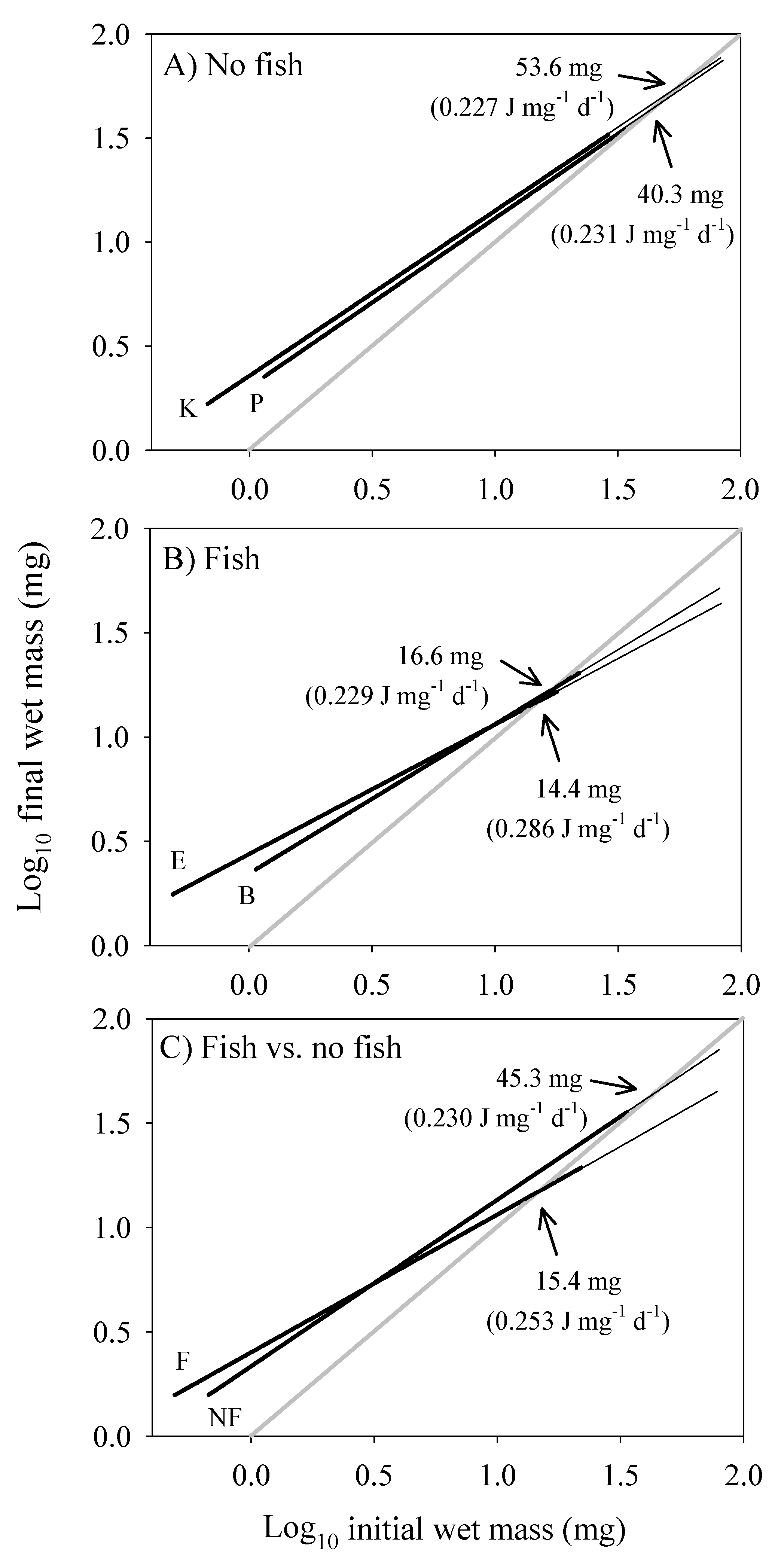
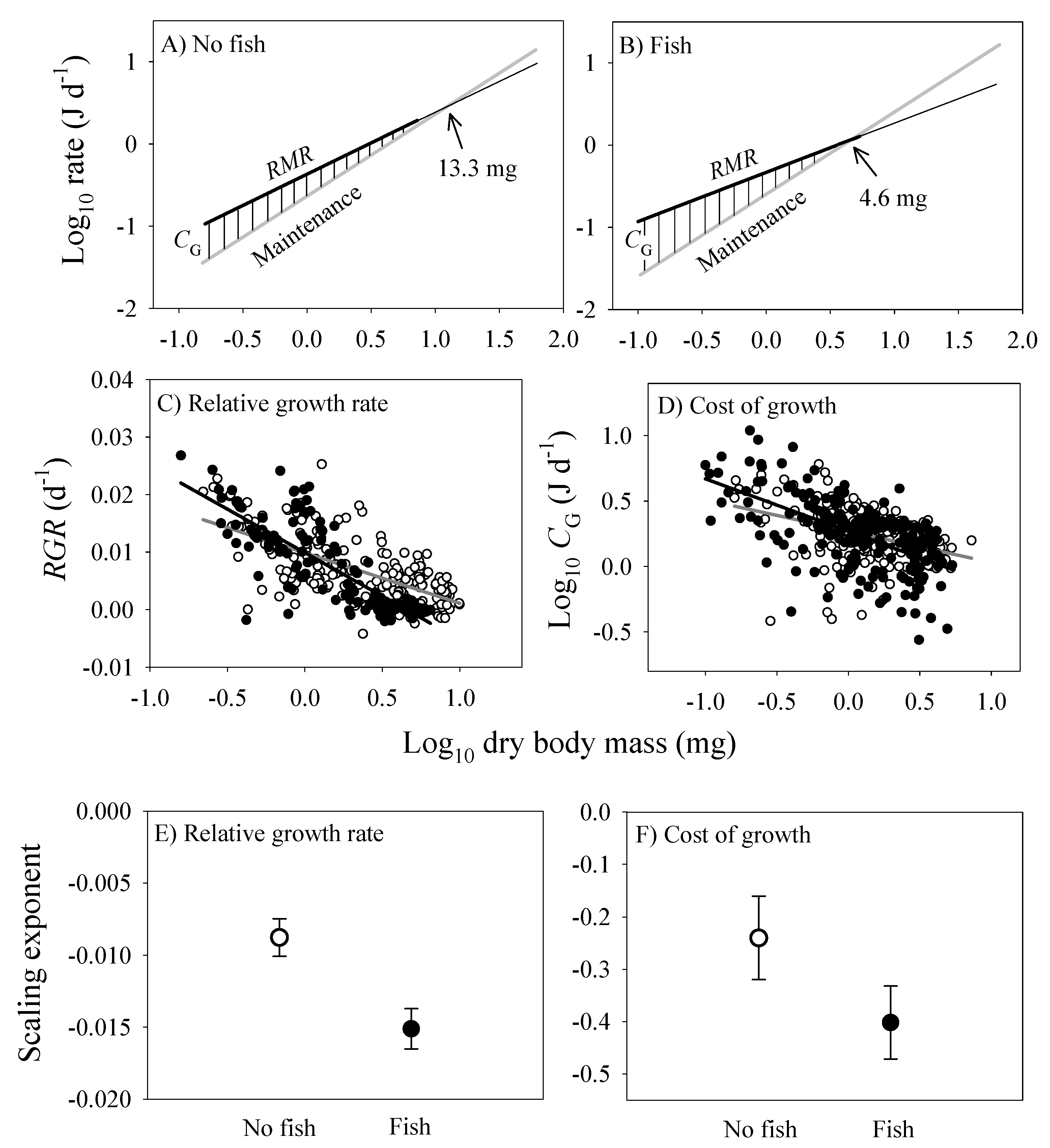
3.4. Scaling of Costs of Growth and Maintenance
3.5. Scaling of Cost of Reproduction of Field-Collected Brooding Females
3.6. Synthetic Allometry of Energy Intake and Use
4. Discussion
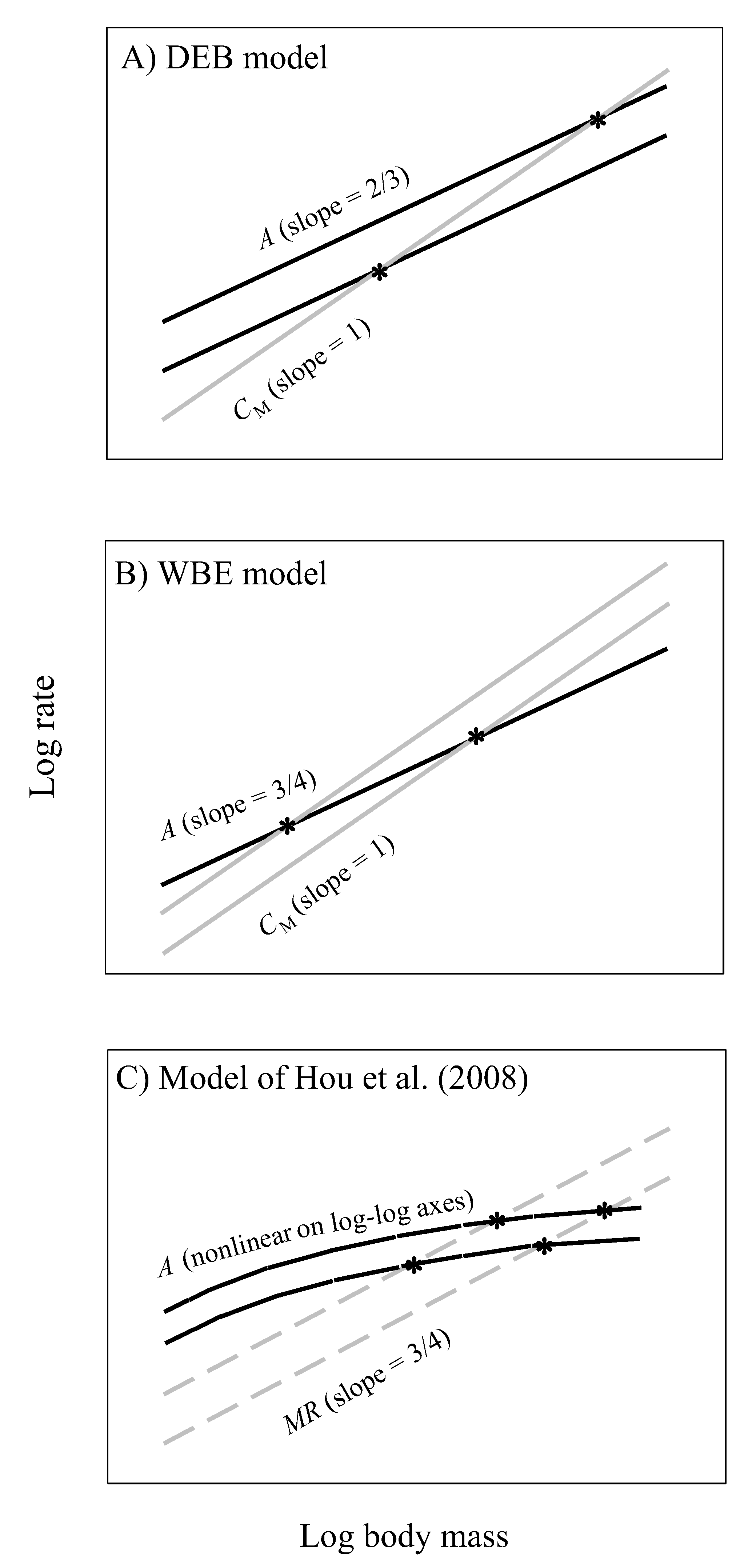
4.1. Relevance to Physical and Geometric Models of Biological Scaling
4.2. Bottom-Up (Resource Supply) Versus Top-Down (Predation) Effects
4.3. Degree of Congruence of Energetic Allometric Responses by Amphipods to Fish Predators
4.4. Mechanisms Underlying Effects of Fish Predators on the Energetic Allometry of Amphipod Prey
4.5. Implications for Growth Models and Other Energy-Based Biological and Ecological Theories
4.5.1. Specific Implications for Growth Models Based on Energetic Scaling Relationships
4.5.2. General Implications for Energy-Based Biological and Ecological Theories
5. Conclusions
- To fully understand the effects of predators on their prey, it is important to consider all of the life stages of prey organisms. Ontogenetic body-mass scaling analyses, as carried out in our study, are one useful way to do this.
- Given the importance of energy for all biological processes, an examination of how predators affect the ontogeny of various energetically significant traits, and their scaling with body mass, can provide valuable insights into age- and size-specific energy-allocation strategies (also see [22,130,150,151]).
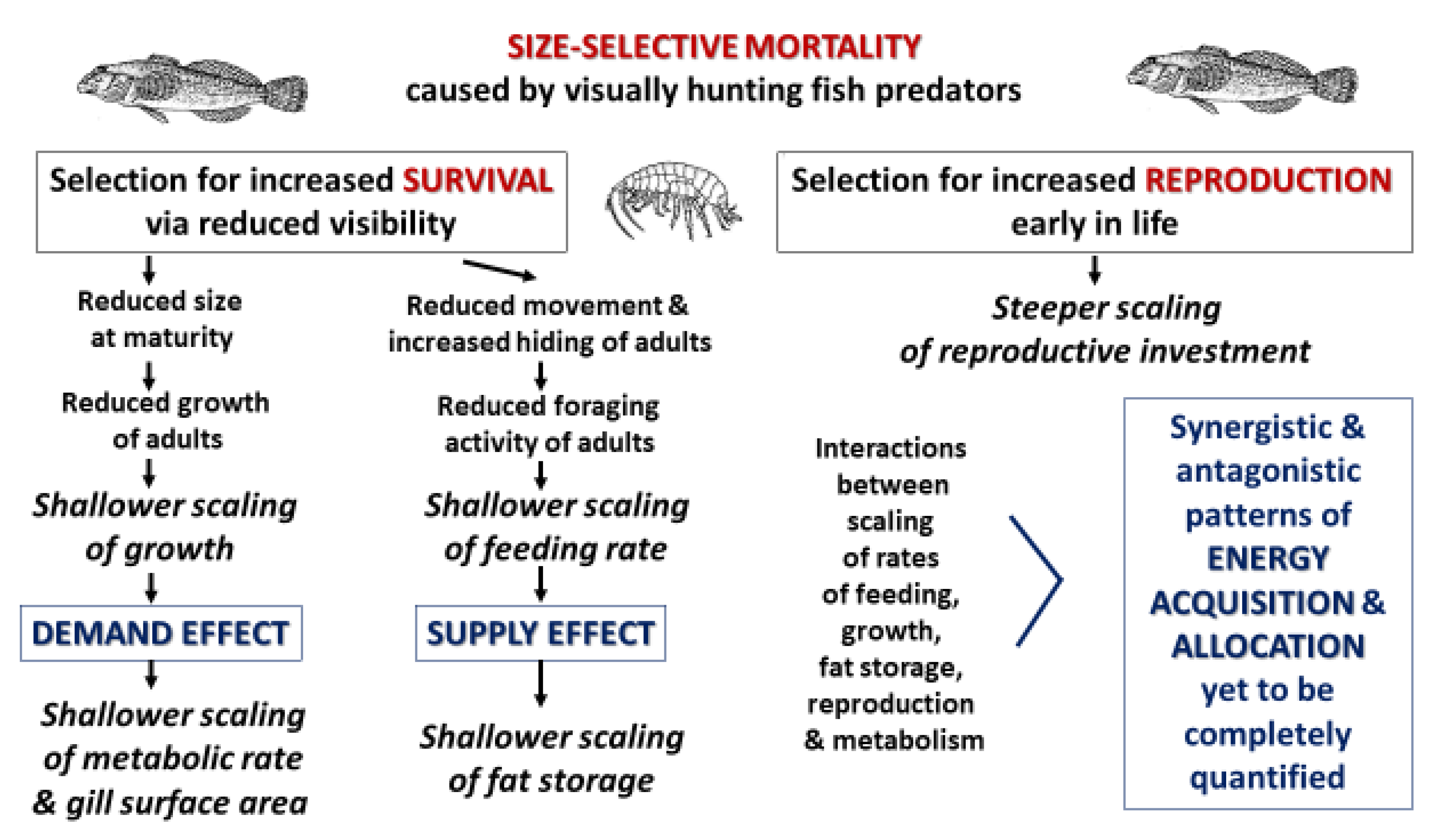
- Various biological processes, such as energy intake and use for various vital functions (e.g., metabolism, growth, locomotion and reproduction) are interdependent in synergistic or antagonistic ways, and thus should be examined with an integrated, holistic perspective. By doing so, our study of the freshwater amphipod Gammarus minus has revealed the ‘symmorphic allometry’ (parallel scaling) of two different clusters of energetically significant traits based on somatic versus reproductive investments that have shifted in coordinated ways in response to the presence of fish predators. With regard to somatic investment, predation risk has caused shallower, congruent scaling of the rates of assimilation, growth and metabolism, fat content, and oxygen uptake capacity (as indicated by gill surface area). Accordingly, adults are smaller and leaner, and have slower rates of growth and metabolism and smaller gills, relative to their body mass, in springs with versus without fish predators. In contrast, traits related to reproductive investment (e.g., total mass and number of egg per brood) scale more steeply in springs with versus without fish predators. Consequently, predation risk intensifies the trade-off between somatic and reproductive investments in G. minus.
- Our results add to growing evidence that biological scaling is ecologically sensitive and evolutionarily malleable, and not merely physically constrained by body design (e.g., [27,29,132,144,152,153,154,155,156,157]). They also show that biological scaling can be affected by both top-down (predation) and bottom-up (resource supply) factors.
- Our findings support the view that growth is not merely the passive result of the body-mass scaling of assimilation rate and maintenance costs, as determined by physical constraints, but is actively regulated to maximize evolutionary fitness in local environments [28]. A comprehensive understanding of biological scaling and other aspects of living systems should be based on not only energy, but also information (also see [28,149,158]).
- Our study of the scaling of various energetically significant traits related to somatic investment focuses on juveniles and adult males to avoid complications regarding egg production. Future research should also examine adult females, because their metabolic responses to predation risk may differ from that of males (see e.g., [22]). In the presence of predators, adult females of G. minus invest more in egg production earlier in life and at smaller body sizes, which may draw energy away from somatic growth and fat storage (cf. [130]). This may, in part, explain why females are smaller than males (in addition to sexual selection for larger size in males to increase their mating success [159,160,161,162]). How the different reproductive strategies of females and males affects their responses to fish predators should be explored.
- We have presented preliminary results on a complex issue. To clarify further our understanding of anti-predator prey responses, we recommend additional research objectives. First, the scaling of key energetic processes, such as food consumption, metabolism and locomotor activity, should be estimated in nature and not just in the laboratory (see e.g., [65]). Second, interactions between various anti-predator responses by prey (e.g., changes in their rates of feeding, metabolism, growth, reproduction and behavioral activity: see e.g., [13,23,24,38,44,45,48,49,130,163]), and reciprocal effects of these responses on the vulnerability of prey to predation should be investigated (following [24,61,164]). Third, effects of other environmental factors (e.g., temperature, habitat, parasites, and food quantity and quality) on prey responses to predator risk deserve further attention (see e.g., [6,38,43,58,116,119,120,165,166,167,168,169,170,171,172,173,174,175,176,177]). Fourth, the relative roles of evolutionary adaptation and phenotypically plastic acclimation involved in prey responses to predators require elucidation (see e.g., [13,48,166,168,178,179,180]).
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B
| Spring Population | Linearregression Equation | Regression Statistics | ||
|---|---|---|---|---|
| r | N | P | ||
| Petersburg | MD = −0.478 (± 0.037) + 0.957 (± 0.034)MW | 0.988 | 92 | <0.00001 |
| Kanesatake | MD = −0.502 (± 0.025) + 0.992 (± 0.022)MW | 0.993 | 116 | <0.00001 |
| Ell | MD = −0.489 (± 0.032) + 0.975 (± 0.034)MW | 0.987 | 91 | <0.00001 |
| Blue | MD = −0.509 (± 0.048) + 0.977 (± 0.047)MW | 0.983 | 63 | <0.00001 |
Appendix C
References
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Lima, S.L. Nonlethal effects in the ecology of predator-prey interactions. BioScience 1998, 48, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to death? The effects of intimidation and consumption in predator–prey interactions. Ecology 2005, 86, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Clinchy, M.; Sheriff, M.J.; Zanette, L.Y. Predator-induced stress and the ecology of fear. Funct. Ecol. 2013, 27, 56–65. [Google Scholar] [CrossRef]
- Sheriff, M.J.; Thaler, J.S. Ecophysiological effects of predation risk; an integration across disciplines. Oecologia 2014, 176, 607–611. [Google Scholar] [CrossRef] [Green Version]
- Reznick, D.A.; Bryga, H.; Endler, J.A. Experimentally induced life-history evolution in a natural population. Nature 1990, 346, 357–359. [Google Scholar] [CrossRef]
- Reznick, D.N.; Travis, J. Experimental studies of evolution and eco-evo dynamics in guppies (Poecilia reticulata). Annu. Rev. Ecol. Evol. Syst. 2019, 50, 335–354. [Google Scholar] [CrossRef]
- Wellborn, G.A. Size-biased predation and prey life histories: A comparative study of freshwater amphipod populations. Ecology 1994, 75, 2104–2117. [Google Scholar] [CrossRef]
- Tollrian, R.; Harvell, C.D. The Ecology and Evolution of Inducible Defenses; Princeton University Press: Princetion, NJ, USA, 1999; pp. 1–383. [Google Scholar]
- Werner, E.E.; Peacor, S.D. A review of trait-mediated indirect interactions in ecological communities. Ecology 2003, 84, 1083–1100. [Google Scholar] [CrossRef] [Green Version]
- Hawlena, D.; Schmitz, O.J. Physiological stress as a fundamental mechanism linking predation to ecosystem functioning. Am. Nat. 2010, 176, 537–556. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.E.; Blumstein, D.T. (Eds.) Escaping from Predators: An Integrative View of Escape Decisions; Cambridge University Press: Cambridge, UK, 2015; pp. 1–441. [Google Scholar]
- Glazier, D.S.; Butler, E.M.; Lombardi, S.A.; Deptola, T.J.; Reese, A.J.; Satterthwaite, E.V. Ecological effects on metabolic scaling: Amphipod responses to fish predators in freshwater springs. Ecol. Monogr. 2011, 81, 599–618. [Google Scholar] [CrossRef]
- Lagos, P.A.; Herberstein, M.E. Are males more scared of predators? Differential change in metabolic rate between males and females under predation risk. Physiol. Behav. 2017, 173, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.L.; Dodson, S.I. Predation, body size, and composition of plankton: The effect of a marine planktivore on lake plankton illustrates theory of size, competition, and predation. Science 1965, 150, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, M.G. Size-selective predation on Daphnia by rainbow trout and yellow perch. Trans. Am. Fish. Soc. 1967, 96, 1–10. [Google Scholar] [CrossRef]
- Werner, E.E.; Hall, D.J. Optimal foraging and the size selection of prey by the bluegill sunfish (Lepomis macrochirus). Ecology 1974, 55, 1042–1052. [Google Scholar] [CrossRef]
- Vince, S.; Valiela, I.; Backus, N.; Teal, J.M. Predation by the salt marsh killifish Fundulus heteroclitus (L.) in relation to prey size and habitat structure: Consequences for prey distribution and abundance. J. Exp. Mar. Biol. Ecol. 1976, 23, 255–266. [Google Scholar] [CrossRef]
- Law, R. Optimal life histories under age-specific predation. Am. Nat. 1979, 114, 399–417. [Google Scholar] [CrossRef]
- Newman, R.M.; Waters, T.F. Size-selective predation on Gammarus pseudolimnaeus by trout and sculpins. Ecology 1984, 65, 1535–1545. [Google Scholar] [CrossRef]
- Allen, J.D. Size-specific predation on marine invertebrate larvae. Biol. Bull. 2008, 214, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Meuthen, D.; Ferrari, M.C.; Lane, T.; Chivers, D.P. Predation risk induces age-and sex-specific morphological plastic responses in the fathead minnow Pimephales promelas. Sci. Rep. 2019, 9, 15738. [Google Scholar] [CrossRef]
- Steiner, U.K.; Van Buskirk, J. Predator-induced changes in metabolism cannot explain the growth/predation risk tradeoff. PLoS ONE 2009, 4, e6160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, C.; Fu, S.J.; Wu, Q.Y.; Cao, Z.D. Predation threat modifies relationships between metabolism and behavioural traits but not their ecological relevance in Chinese bream. Mar. Freshw. Behav. Physiol. 2017, 50, 329–344. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Sibly, R.M.; Brown, J.H.; Kodric-Brown, A. (Eds.) Metabolic Ecology: A Scaling Approach; John Wiley & Sons: New York, NY, USA, 2012; pp. 1–375. [Google Scholar]
- Glazier, D.S. Metabolic scaling in complex living systems. Systems 2014, 2, 451–540. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S. Is metabolic rate a universal ‘pacemaker’ for biological processes? Biol. Rev. 2015, 90, 377–407. [Google Scholar] [CrossRef]
- Harrison, J.F. Do performance–safety tradeoffs cause hypometric metabolic scaling in animals? Trends Ecol. Evol. 2017, 32, 653–664. [Google Scholar] [CrossRef]
- Glazier, D.S. Resource supply and demand both affect metabolic scaling: A response to Harrison. Trends Ecol. Evol. 2018, 33, 237–238. [Google Scholar] [CrossRef]
- Briceño, F.A.; Polymeropoulos, E.T.; Fitzgibbon, Q.P.; Dambacher, J.M.; Pecl, G.T. Changes in metabolic rate of spiny lobster under predation risk. Mar. Ecol. Prog. Ser. 2018, 598, 71–84. [Google Scholar] [CrossRef]
- Paul, N.; Novais, S.C.; Lemos, M.F.; Kunzmann, A. Chemical predator signals induce metabolic suppression in rock goby (Gobius paganellus). PLoS ONE 2018, 13, e0209286. [Google Scholar] [CrossRef] [Green Version]
- Manzur, T.; Vidal, F.; Pantoja, J.F.; Fernández, M.; Navarrete, S.A. Behavioural and physiological responses of limpet prey to a seastar predator and their transmission to basal trophic levels. J. Anim. Ecol. 2014, 83, 923–933. [Google Scholar] [CrossRef] [Green Version]
- Kolar, V.; Boukal, D.S.; Sentis, A. Predation risk and habitat complexity modify intermediate predator feeding rates and energetic efficiencies in a tri-trophic system. Freshwater Biol. 2019, 64, 1480–1491. [Google Scholar] [CrossRef]
- Barry, M.J.; Syal, S. Metabolic responses of tadpoles to chemical predation cues. Hydrobiologia 2013, 700, 267–276. [Google Scholar] [CrossRef]
- Stibor, H.; Machacek, J. The influence of fish-exuded chemical signals on the carbon budget of Daphnia. Limnol. Oceanogr. 1998, 43, 997–1000. [Google Scholar] [CrossRef]
- Beckerman, A.P.; Wieski, K.; Baird, D.J. Behavioural versus physiological mediation of life history under predation risk. Oecologia 2007, 152, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, K.; Stoks, R.; De Meester, L. Enhanced anti-predator defence in the presence of food stress in the water flea Daphnia magna. Funct. Ecol. 2010, 24, 322–329. [Google Scholar] [CrossRef]
- Robison, A.L.; Chapman, T.; Bidwell, J.R. Predation cues influence metabolic rate and sensitivity to other chemical stressors in fathead minnows (Pimephales promelas) and Daphnia pulex. Ecotoxicology 2018, 27, 55–68. [Google Scholar] [CrossRef]
- Toscano, B.J.; Monaco, C.J. Testing for relationships between individual crab behavior and metabolic rate across ecological contexts. Behav. Ecol. Sociobiol. 2015, 69, 1343–1351. [Google Scholar] [CrossRef]
- Antoł, A.; Kierat, J.; Czarnoleski, M. Sedentary prey facing an acute predation risk: Testing the hypothesis of inducible metabolite emission suppression in zebra mussels, Dreissena polymorpha. Hydrobiologia 2018, 810, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Slos, S.; Stoks, R. Predation risk induces stress proteins and reduces antioxidant defense. Funct. Ecol. 2008, 22, 637–642. [Google Scholar] [CrossRef]
- Culler, L.E.; McPeek, M.A.; Ayres, M.P. Predation risk shapes thermal physiology of a predaceous damselfly. Oecologia 2014, 176, 653–660. [Google Scholar] [CrossRef]
- Ling, H.; Fu, S.J.; Zeng, L.Q. Predator stress decreases standard metabolic rate and growth in juvenile crucian carp under changing food availability. Comp. Bioch. Physiol. A Molec. Integr. Physiol. 2019, 231, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, I.J.; Aho, J.; Vornanen, M.; Huuskonen, H. Phenotypic plasticity and predator effects on morphology and physiology of crucian carp in nature and in the laboratory. J. Fish Biol. 1997, 50, 781–798. [Google Scholar] [CrossRef]
- Woodley, C.M.; Peterson, M.S. Measuring responses to simulated predation threat using behavioral and physiological metrics: The role of aquatic vegetation. Oecologia 2003, 136, 155–160. [Google Scholar] [PubMed]
- Milano, D.; Lozada, M.; Zagarese, H.E. Predator-induced reaction patterns of landlocked Galaxias maculatus to visual and chemical cues. Aquat. Ecol. 2010, 44, 741–748. [Google Scholar] [CrossRef]
- Handelsman, C.A.; Broder, E.D.; Dalton, C.M.; Ruell, E.W.; Myrick, C.A.; Reznick, D.N.; Ghalambor, C.K. Predator-induced phenotypic plasticity in metabolism and rate of growth: Rapid adaptation to a novel environment. Integr. Comp. Biol. 2013, 53, 975–988. [Google Scholar] [CrossRef]
- Auer, S.K.; Dick, C.A.; Metcalfe, N.B.; Reznick, D.N. Metabolic rate evolves rapidly and in parallel with the pace of life history. Nat. Commun. 2018, 9, 14. [Google Scholar] [CrossRef]
- Sunardi; Asaeda, T.; Manatunge, J. Physiological responses of topmouth gudgeon, Pseudorasbora parva, to predator cues and variation of current velocity. Aquat. Ecol. 2007, 41, 111–118. [Google Scholar]
- Xu, J.J.; Fu, S.J.; Fu, C. Physiological and behavioral stress responses to predators are altered by prior predator experience in juvenile qingbo (Spinibarbus sinensis). Biol. Open 2019, 8, bio041012. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Fu, S.J.; Yuan, X.Z.; Cao, Z.D. Predator-driven intra-species variation in locomotion, metabolism and water velocity preference in pale chub (Zacco platypus) along a river. J. Exp. Biol. 2015, 218, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Okuyama, T. Metabolic responses to predation risk in a jumping spider. J. Zool. 2015, 297, 9–14. [Google Scholar] [CrossRef]
- Hawlena, D.; Schmitz, O.J. Herbivore physiological response to predation risk and implications for ecosystem nutrient dynamics. Proc. Natl. Acad. Sci. USA 2010, 107, 15503–15507. [Google Scholar] [CrossRef] [Green Version]
- Ameri, M.; Kemp, D.J.; Barry, K.L.; Herberstein, M.E. Predatory chemical cues decrease attack time and increase metabolic rate in an orb-web spider. J. Exp. Biol. 2019, 222, jeb.212738. [Google Scholar] [CrossRef] [PubMed]
- Tigreros, N.; Wang, E.H.; Thaler, J.S. Prey nutritional state drives divergent behavioural and physiological responses to predation risk. Funct. Ecol. 2018, 32, 982–989. [Google Scholar] [CrossRef]
- Tigreros, N.; Wang, E.H.; Thaler, J.S. Maternal effects across life stages: Larvae experiencing predation risk increase offspring provisioning. Ecol. Entomol. 2019, 44, 738–744. [Google Scholar] [CrossRef]
- Thaler, J.S.; Contreras, H.; Davidowitz, G. Effects of predation risk and plant resistance on Manduca sexta caterpillar feeding behaviour and physiology. Ecol. Entomol. 2014, 39, 210–216. [Google Scholar] [CrossRef]
- Krams, I.; Kivleniece, I.; Kuusik, A.; Krama, T.; Mänd, R.; Rantala, M.J.; Znotiņa, S.; Freeberg, T.M.; Mänd, M. Predation promotes survival of beetles with lower resting metabolic rates. Entomol. Exp. Appl. 2013, 148, 94–103. [Google Scholar] [CrossRef]
- Krams, I.; Kivleniece, I.; Kuusik, A.; Krama, T.; Freeberg, T.M.; Mänd, R.; Vrublevska, J.; Rantala, M.J.; Mänd, M. Predation selects for low resting metabolic rate and consistent individual differences in anti-predator behavior in a beetle. Acta Ethol. 2013, 16, 163–172. [Google Scholar] [CrossRef]
- Mathot, K.J.; Abbey-Lee, R.N.; Kempenaers, B.; Dingemanse, N.J. Do great tits (Parus major) suppress basal metabolic rate in response to increased perceived predation danger? A field experiment. Physiol. Behav. 2016, 164, 400–406. [Google Scholar] [CrossRef]
- Smith, E.N.; Sims, K.; Vich, J.F. Oxygen consumption of frightened swamp rabbits, Sylvilagus aquaticus. Comp. Biochem. Physiol. A Physiol. 1981, 70, 533–536. [Google Scholar] [CrossRef]
- Chabot, D.; Gagnon, P.; Dixon, E.A. Effect of predator odors on heart rate and metabolic rate of wapiti (Cervus elaphus canadensis). J. Chem. Ecol. 1996, 22, 839–868. [Google Scholar] [CrossRef]
- Ward, J.F.; MacDonald, D.W.; Doncaster, C.P.; Mauget, C. Physiological response of the European hedgehog to predator and nonpredator odour. Physiol. Behav. 1996, 60, 1469–1472. [Google Scholar] [CrossRef]
- Pettett, C.E.; Johnson, P.J.; Moorhouse, T.P.; Hambly, C.; Speakman, J.R.; Macdonald, D.W. Daily energy expenditure in the face of predation: Hedgehog energetics in rural landscapes. J. Exp. Biol. 2017, 220, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Holsinger, J.R. The Freshwater Amphipod Crustaceans (Gammaridae) in North America; United States Environmental Protection Agency: Cincinnati, OH, USA, 1976; pp. 1–89.
- Culver, D.C.; Kane, T.C.; Fong, D.W. Adaptation and Natural Selection in Caves: The Evolution of Gammarus minus; Harvard University Press: Cambridge, MA, USA, 1995; pp. 1–235. [Google Scholar]
- Glazier, D.S.; Gooch, J.L. Macroinvertebrate assemblages in Pennsylvania (USA) springs. Hydrobiologia 1987, 150, 33–43. [Google Scholar] [CrossRef]
- Glazier, D.S.; Horne, M.T.; Lehman, M.E. Abundance, body composition and reproductive output of Gammarus minus (Crustacea: Amphipoda) in ten cold springs differing in pH and ionic content. Freshwater Biol. 1992, 28, 149–163. [Google Scholar] [CrossRef]
- Glazier, D.S.; Paul, D.A. Ecology of ontogenetic body-mass scaling of gill-surface area in a freshwater crustacean. J. Exp. Biol. 2017, 220, 2120–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazier, D.S. Variation in offspring investment within and among populations of Gammarus minus Say (Crustacea: Amphipoda) in ten mid-Appalachian springs (USA). Arch. Hydrobiol. 1999, 146, 257–284. [Google Scholar] [CrossRef]
- Glazier, D.S. Springs as model ecosystems for ecology and evolutionary biology: A case study of Gammarus minus Say (Amphipoda) in mid-Appalachian springs differing in pH and ionic content. In Studies in Crenobiology. The Biology of Springs and Springbrooks; Botosaneanu, L., Ed.; Backhuys: Leiden, The Netherlands, 1998; pp. 49–62. [Google Scholar]
- Glazier, D.S.; Deptola, T.J. The amphipod Gammarus minus has larger eyes in freshwater springs with numerous fish predators. Invert. Biol. 2011, 130, 60–67. [Google Scholar] [CrossRef]
- Glazier, D.S. Springs. In Reference Module in Earth Systems and Environmental Sciences; Elias, S.A., Ed.; Elsevier: Waltham, MA, USA, 2014; pp. 1–78. [Google Scholar]
- Glazier, D.S. The fauna of North American temperate cold springs: Patterns and hypotheses. Freshwater Biol. 1991, 26, 527–542. [Google Scholar] [CrossRef]
- Gooch, J.L. Spatial genetic patterns in relation to regional history and structure: Gammarus minus (Amphipoda) in Appalachian watersheds. Am. Midl. Nat. 1990, 124, 93–104. [Google Scholar] [CrossRef]
- Gooch, J.L.; Glazier, D.S. Temporal and spatial patterns in mid-Appalachian springs. Mem. Entomol. Soc. Can. 1991, 155, 29–49. [Google Scholar] [CrossRef]
- Carlini, D.B.; Manning, J.; Sullivan, P.G.; Fong, D.W. Molecular genetic variation and population structure in morphologically differentiated cave and surface populations of the freshwater amphipod Gammarus minus. Mol. Ecol. 2009, 18, 1932–1945. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S.; Sparks, B.L. Energetics of amphipods in ion-poor waters: Stress resistance is not invariably linked to low metabolic rates. Funct. Ecol. 1997, 11, 126–128. [Google Scholar] [CrossRef]
- Glazier, D.S. Amphipoda. In Reference Module in Earth Systems and Environmental Sciences; Elias, S.A., Ed.; Elsevier: Waltham, MA, USA, 2014; pp. 1–49. [Google Scholar]
- Glazier, D.S. Is fatter fitter? Body storage and reproduction in ten populations of the freshwater amphipod Gammarus minus. Oecologia 2000, 122, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Marchant, R.; Hynes, H.B.N. Field estimates of feeding rate for Gammarus pseudolimnaeus (Crustacea: Amphipoda) in the Credit River, Ontario. Freshwater Biol. 1981, 11, 27–36. [Google Scholar] [CrossRef]
- Naylor, C.; Maltby, L.; Calow, P. Scope for growth in Gammarus pulex, a freshwater detritivore. Hydrobiologia 1989, 188-189, 517–523. [Google Scholar] [CrossRef]
- Graça, M.A.S.; Maltby, L.; Calow, P. Importance of fungi in the diet of Gammarus pulex and Asellus aquaticus. II. Effects on growth, reproduction and physiology. Oecologia 1993, 96, 304–309. [Google Scholar] [CrossRef]
- Kostalos, M.S.; Seymour, R.L. Role of microbial enriched detritus in the nutrition of Gammarus minus (Amphipoda). Oikos 1976, 27, 512–516. [Google Scholar] [CrossRef]
- Suberkropp, K. Interactions with invertebrates. In The Fungal Community; Carroll, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 729–747. [Google Scholar]
- Graça, M.A.S.; Maltby, L.; Calow, P. Importance of fungi in the diet of Gammarus pulex and Asellus aquaticus. I. Feeding strategies. Oecologia 1993, 93, 139–144. [Google Scholar]
- Bärlocher, F. The role of fungi in the nutrition of stream invertebrates. Bot. J. Linn. Soc. 1985, 91, 83–94. [Google Scholar] [CrossRef]
- Bärlocher, F.; Kendrick, B. Assimilation efficiency of Gammarus pseudolimnaeus (Amphipoda) feeding on fungal mycelium or autumn-shed leaves. Oikos 1985, 26, 55–59. [Google Scholar] [CrossRef]
- Åbjörnsson, K.; Dahl, J.; Nyström, P.; Brönmark, C. Influence of predator and dietary chemical cues on the behaviour and shredding efficiency of Gammarus pulex. Aquat. Ecol. 2000, 34, 379–387. [Google Scholar] [CrossRef]
- Konishi, M.; Nakano, S.; Iwata, T. Trophic cascading effects of predatory fish on leaf litter processing in a Japanese stream. Ecol. Res. 2001, 16, 415–422. [Google Scholar] [CrossRef]
- Sangiorgio, F.; Glazier, D.S.; Mancinelli, G.; Basset, A. How can habitat size influence leaf litter decomposition in five mid-Appalachian springs (USA)? The importance of the structure of the detritivorous guild. Hydrobiologia 2010, 654, 227–236. [Google Scholar] [CrossRef]
- Beermann, J.; Boos, K.; Gutow, L.; Boersma, M.; Peralta, A.C. Combined effects of predator cues and competition define habitat choice and food consumption of amphipod mesograzers. Oecologia 2018, 186, 645–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gee, J.H.R. Population dynamics and morphometrics of Gammarus pulex L.: Evidence of seasonal food limitation in a freshwater detritivore. Freshwater Biol. 1988, 19, 333–343. [Google Scholar] [CrossRef]
- Elliott, J.M.; Davison, W. Energy equivalents of oxygen consumption in animal energetics. Oecologia 1975, 19, 195–201. [Google Scholar] [CrossRef]
- Kooijman, S.A.L.M. Dynamic Energy and Mass Budgets in Biological Systems; Cambridge University Press: Cambridge, UK, 2000; pp. 1–424. [Google Scholar]
- Van der Meer, J. Metabolic theories in ecology. Trends Ecol. Evol. 2006, 21, 136–140. [Google Scholar] [CrossRef]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for ontogenetic growth. Nature 2001, 413, 628–631. [Google Scholar] [CrossRef]
- Smith, R.J. Use and misuse of the reduced major axis for line-fitting. Am. J. Phys. Anthropol. 2009, 140, 476–486. [Google Scholar] [CrossRef]
- Glazier, D.S. A unifying explanation for diverse metabolic scaling in animals and plants. Biol. Rev. 2010, 85, 111–138. [Google Scholar] [CrossRef] [PubMed]
- White, C.R. Allometric estimation of metabolic rates in animals. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2011, 158, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Kerkhoff, A.J.; Enquist, B. Multiplicative by nature: Why logarithmic transformation is necessary in allometry. J. Theor. Biol. 2009, 257, 519–521. [Google Scholar] [CrossRef]
- Glazier, D.S. Log-transformation is useful for examining proportional relationships in allometric scaling. J. Theor. Biol. 2009, 334, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Cumming, G. Inference by eye: Reading the overlap of independent confidence intervals. Stat. Med. 2008, 28, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Rubner, M. Über den Einfluss der Körpergrösse auf Stoff- und Kraftwechsel. Z. Biol. 1883, 19, 535–562. [Google Scholar]
- Kleiber, M. The Fire of Life; Wiley: New York, NY, USA, 1961; pp. 1–454. [Google Scholar]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef]
- Savage, V.M.; Gillooly, J.F.; Woodruff, W.H.; West, G.B.; Allen, A.P.; Enquist, B.J.; Brown, J.H. The predominance of quarter-power scaling in biology. Funct. Ecol. 2004, 18, 257–282. [Google Scholar] [CrossRef]
- Gould, S.J. Allometry and size in ontogeny and phylogeny. Biol. Rev. 1966, 41, 587–638. [Google Scholar] [CrossRef]
- Sebens, K.P. The limits to indeterminate growth: An optimal size model applied to passive suspension feeders. Ecology 1982, 63, 209–222. [Google Scholar] [CrossRef]
- Reiss, M.J. The allometry of reproduction: Why larger species invest relatively less in their offspring. J. Theor. Biol. 1985, 113, 529–544. [Google Scholar] [CrossRef]
- Banavar, J.R.; Moses, M.E.; Brown, J.H.; Damuth, J.; Rinaldo, A.; Sibly, R.M.; Maritan, A. A general basis for quarter-power scaling in animals. Proc. Natl. Acad. Sci. USA 2010, 107, 15816–15820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobling, M. Fish Bioenergetics; Chapman & Hall: London, UK, 1994; pp. 1–310. [Google Scholar]
- Essington, T.E.; Kitchell, J.F.; Walters, C.J. The von Bertalanffy growth function, bioenergetics, and the consumption rates of fish. Can. J. Fish. Aquat. Sci. 2001, 58, 2129–2138. [Google Scholar] [CrossRef]
- Basset, A.; Cozzoli, F.; Paparella, F. A unifying approach to allometric scaling of resource ingestion rates under limiting conditions. Ecosphere 2012, 3, 1–14. [Google Scholar] [CrossRef]
- Brown, J.S.; Kotler, B.P.; Smith, R.J.; Wirtz, W.O. The effects of owl predation on the foraging behavior of heteromyid rodents. Oecologia 1988, 76, 408–415. [Google Scholar] [CrossRef]
- Wooster, D.; Sih, A. A review of the drift and activity responses of stream prey to predator presence. Oikos 1995, 73, 3–8. [Google Scholar] [CrossRef]
- Verdolin, J.L. Meta-analysis of foraging and predation risk trade-offs in terrestrial systems. Behav. Ecol. Sociobiol. 2006, 60, 457–464. [Google Scholar] [CrossRef]
- Kolar, C.S.; Rahel, F.J. Interaction of a biotic factor (predator presence) and an abiotic factor (low oxygen) as an influence on benthic invertebrate communities. Oecologia 1993, 95, 210–219. [Google Scholar] [CrossRef]
- Szokoli, F.; Winkelmann, C.; Berendonk, T.U.; Worischka, S. The effects of fish kairomones and food availability on the predator avoidance behaviour of Gammarus pulex. Arch. Hydrobiol. 2015, 186, 249–258. [Google Scholar] [CrossRef]
- Jermacz, Ł.; Kobak, J. Keep calm and don’t stop growing: Non-consumptive effects of a sympatric predator on two invasive Ponto-Caspian gammarids Dikerogammarus villosus and Pontogammarus robustoides. PLoS ONE 2017, 12, e0182481. [Google Scholar] [CrossRef] [Green Version]
- Friberg, N.; Andersen, T.H.; Hansen, H.O.; Iversen, T.M.; Jacobsen, D.; Krøjgaard, L.; Larsen, S.E. The effect of brown trout (Salmo trutta L.) on stream invertebrate drift, with special reference to Gammarus pulex L. Hydrobiologia 1994, 294, 105–110. [Google Scholar] [CrossRef]
- MacNeil, C.; Dick, J.T.; Elwood, R.W. The trophic ecology of freshwater Gammarus spp. (Crustacea: Amphipoda): Problems and perspectives concerning the functional feeding group concept. Biol. Rev. 1997, 72, 349–364. [Google Scholar] [CrossRef]
- Weibel, E.R.; Taylor, C.R.; Hoppeler, H. The concept of symmorphosis: A testable hypothesis of structure-function relationship. Proc. Natl. Acad. Sci. USA. 1991, 88, 10357–10361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weibel, E.R. Symmorphosis: On Form and Function in Shaping Life; Harvard University Press: Cambridge, MA, USA, 2000; pp. 1–263. [Google Scholar]
- Charlesworth, B. Evolution in Age-Structured Populations; Cambridge University Press: Cambridge, UK, 1994; pp. 1–306. [Google Scholar]
- Sibly, R.M.; Calow, P. Physiological Ecology of Animals: An Evolutionary Approach; Blackwell Scientific Publications: Oxford, UK, 1986; pp. 1–179. [Google Scholar]
- Roff, D.A. The Evolution of Life Histories: Theory and Analysis; Chapman & Hall: New York, NY, USA, 1992; pp. 1–535. [Google Scholar]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992; pp. 1–249. [Google Scholar]
- Wellborn, G.A. The mechanistic basis of body size differences between two Hyalella (Amphipoda) species. J. Freshwater Ecol. 1994, 9, 159–168. [Google Scholar] [CrossRef]
- Sears, K.E.; Kerkhoff, A.J.; Messerman, A.; Itagaki, H. Ontogenetic scaling of metabolism, growth, and assimilation: Testing metabolic scaling theory with Manduca sexta larvae. Physiol. Biochem. Zool. 2012, 85, 159–173. [Google Scholar] [CrossRef]
- Glazier, D.S. Beyond the “3/4-power law”: Variation in the intra- and interspecific scaling of metabolic rate in animals. Biol. Rev. 2005, 80, 611–662. [Google Scholar] [CrossRef]
- Glazier, D.S. Activity affects intraspecific body-size scaling of metabolic rate in ectothermic animals. J. Comp. Physiol. B 2009, 179, 821–828. [Google Scholar] [CrossRef]
- Nakaoka, M. Nonlethal effects of predators on prey populations: Predator-mediated change in bivalve growth. Ecology 2000, 81, 1031–1045. [Google Scholar] [CrossRef]
- Guo, J.; Martín, P.R.; Zhang, C.; Zhang, J.E. Predation risk affects growth and reproduction of an invasive snail and its lethal effect depends on prey size. PLoS ONE 2017, 12, e0187747. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.D.; Moore, K.A. The effect of environmental factors on the activity of Gammarus pseudolimnaeus (Amphipoda). Hydrobiologia 1982, 96, 137–147. [Google Scholar] [CrossRef]
- Andersson, K.G.; Brönmark, C.; Herrmann, J.; Malmqvist, B.; Otto, C.; Sjörström, P. Presence of sculpins (Cottus gobio) reduces drift and activity of Gammarus pulex (Amphipoda). Hydrobiologia 1986, 133, 209–215. [Google Scholar] [CrossRef]
- Holomuski, J.R.; Hoyle, J.D. Effect of predatory fish presence on habitat use and diel movement of the stream amphipod, Gammarus minus. Freshwater Biol. 1990, 24, 509–517. [Google Scholar] [CrossRef]
- Wooster, D.E. Amphipod (Gammarus minus) responses to predators and predator impact on amphipod density. Oecologia 1998, 115, 253–259. [Google Scholar] [CrossRef]
- James, W.R.; McClintock, J.B. Anti-predator responses of amphipods are more effective in the presence of conspecific chemical cues. Hydrobiologia 2017, 797, 277–288. [Google Scholar] [CrossRef]
- Glazier, D. Body-mass scaling of metabolic rate: What are the relative roles of cellular versus systemic effects? Biology 2015, 4, 187–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirst, A.G.; Glazier, D.S.; Atkinson, D. Body shape shifting during growth permits tests that distinguish between competing geometric theories of metabolic scaling. Ecol. Lett. 2014, 17, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S.; Hirst, A.G.; Atkinson, D. Shape shifting predicts ontogenetic changes in metabolic scaling in diverse aquatic invertebrates. Proc. R Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D. Rediscovering and reviving old observations and explanations of metabolic scaling in living systems. Systems 2018, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Pütter, A. Studien über physiologische Ähnlichkeit VI. Wachstumähnlichkeiten. Pflügers. Arch Gesamte Physiol. Menschen. Tiere. 1920, 180, 298–340. [Google Scholar] [CrossRef]
- Von Bertalanffy, L. Quantitative laws in metabolism and growth. Quart. Rev. Biol. 1957, 32, 217–231. [Google Scholar] [CrossRef]
- Hou, C.; Zuo, W.; Moses, M.E.; Woodruff, W.H.; Brown, J.H.; West, G.B. Energy uptake and allocation during ontogeny. Science 2008, 322, 736–739. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A. Energy flow in growth and production. Trends Ecol. Evol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Resource allocation patterns. In Resource Allocation Theory Applied to Farm Animal Production; Rauw, W.M., Ed.; CAB International: Wallingford, UK, 2009; pp. 22–43. [Google Scholar]
- Killen, S.S.; Gamperl, A.K.; Brown, J.A. Ontogeny of predator-sensitive foraging and routine metabolism in larval shorthorn sculpin, Myoxocephalus scorpius. Mar. Biol. 2007, 152, 1249–1261. [Google Scholar] [CrossRef]
- Kortet, R.; Laakkonen, M.V.; Tikkanen, J.; Vainikka, A.; Hirvonen, H. Size-dependent stress response in juvenile Arctic charr (Salvelinus alpinus) under prolonged predator conditioning. Aquacult. Res. 2019, 50, 1482–1490. [Google Scholar] [CrossRef]
- Glazier, D.S. The 3/4-power law is not universal: Evolution of isometric, ontogenetic metabolic scaling in pelagic animals. BioScience 2006, 56, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S. Scaling of metabolic scaling within physical limits. Systems 2014, 2, 425–450. [Google Scholar] [CrossRef] [Green Version]
- Killen, S.S.; Atkinson, D.; Glazier, D.S. The intraspecific scaling of metabolic rate with body mass in fishes depends on lifestyle and temperature. Ecol. Lett. 2010, 13, 184–193. [Google Scholar] [CrossRef]
- White, C.R.; Kearney, M.R. Metabolic scaling in animals: Methods, empirical results, and theoretical explanations. Compr. Physiol. 2014, 4, 231–256. [Google Scholar]
- Pequeno, P.A.; Baccaro, F.B.; Souza, J.L.; Franklin, E. Ecology shapes metabolic and life history scalings in termites. Ecol. Eng. 2017, 42, 115–124. [Google Scholar] [CrossRef]
- Fossen, E.I.; Pélabon, C.; Einum, S. Genetic and environmental effects on the scaling of metabolic rate with body size. J. Exp. Biol. 2019, 222, jeb193243. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, M.I.; Pennell, M.; Altermatt, F.; Matthews, B.; Melian, C.; Gonzalez, A. Principles of ecology revisited: Integrating information and ecological theories for a more unified science. Front. Ecol. Evol. 2019, 7, 219. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.I. Sexual selection, natural selection, and body size in Gammarus pulex (Amphipoda). Am. Nat. 1988, 131, 348–359. [Google Scholar] [CrossRef]
- Jormalainen, V. Precopulatory mate guarding in crustaceans: Male competitive strategy and intersexual conflict. Quart. Rev. Biol. 1998, 73, 275–304. [Google Scholar] [CrossRef]
- Bollache, L.; Cézilly, F. Sexual selection on male body size and assortative pairing in Gammarus pulex (Crustacea: Amphipoda): Field surveys and laboratory experiments. J. Zool. 2004, 264, 135–141. [Google Scholar] [CrossRef]
- Cothran, R.D. The mechanistic basis of a large male mating advantage in two freshwater amphipod species. Ethology 2008, 114, 1145–1153. [Google Scholar] [CrossRef]
- Steiner, U.K. Linking antipredator behaviour, ingestion, gut evacuation and costs of predator-induced responses in tadpoles. Anim. Behav. 2007, 74, 1473–1479. [Google Scholar] [CrossRef] [Green Version]
- Killen, S.S.; Marras, S.; McKenzie, D.J. Fuel, fasting, fear: Routine metabolic rate and food deprivation exert synergistic effects on risk-taking in individual juvenile European sea bass. J. Anim. Ecol. 2011, 80, 1024–1033. [Google Scholar] [CrossRef]
- Weetman, D.; Atkinson, D.; Chubb, J.C. Effects of temperature on anti-predator behaviour in the guppy, Poecilia reticulata. Anim. Behav. 1998, 55, 1361–1372. [Google Scholar] [CrossRef] [Green Version]
- Benesh, D.P.; Kitchen, J.; Pulkkinen, K.; Hakala, I.; Valtonen, E.T. The effect of Echinorhynchus borealis (Acanthocephala) infection on the anti-predator behavior of a benthic amphipod. J. Parasitol. 2008, 94, 542–546. [Google Scholar] [CrossRef]
- Dalesman, S.; Rundle, S.D. Influence of rearing and experimental temperatures on predator avoidance behaviour in a freshwater pulmonate snail. Freshwater Biol. 2010, 55, 2107–2113. [Google Scholar] [CrossRef]
- Välimäki, K.; Herczeg, G. Ontogenetic and evolutionary effects of predation and competition on nine-spined stickleback (Pungitius pungitius) body size. J. Anim. Ecol. 2012, 81, 859–867. [Google Scholar] [CrossRef]
- Miller, L.P.; Matassa, C.M.; Trussell, G.C. Climate change enhances the negative effects of predation risk on an intermediate consumer. Global Change Biol. 2014, 20, 3834–3844. [Google Scholar] [CrossRef] [PubMed]
- Janssens, L.; Van Dievel, M.; Stoks, R. Warming reinforces nonconsumptive predator effects on prey growth, physiology, and body stoichiometry. Ecology 2015, 96, 3270–3280. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, O.J.; Rosenblatt, A.E.; Smylie, M. Temperature dependence of predation stress and the nutritional ecology of a generalist herbivore. Ecology 2016, 97, 3119–3130. [Google Scholar] [CrossRef] [PubMed]
- Bakker, T.C.; Frommen, J.G.; Thünken, T. Adaptive parasitic manipulation as exemplified by acanthocephalans. Ethology 2017, 123, 779–784. [Google Scholar] [CrossRef]
- Laws, A.N. Climate change effects on predator–prey interactions. Curr. Opin. Insect Sci. 2017, 23, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, O.J.; Rosenblatt, A.E. The temperature dependence of predation stress and prey nutritional stoichiometry. Front. Ecol. Evol. 2017, 5, 73. [Google Scholar] [CrossRef] [Green Version]
- Dalton, C.M.; Tracy, K.E.; Hairston, N.G.; Flecker, A.S. Fasting or fear: Disentangling the roles of predation risk and food deprivation in the nitrogen metabolism of consumers. Ecology 2018, 99, 681–689. [Google Scholar] [CrossRef]
- Luhring, T.M.; Vavra, J.M.; Cressler, C.E.; DeLong, J.P. Phenotypically plastic responses to predation risk are temperature dependent. Oecologia 2019, 191, 709–719. [Google Scholar] [CrossRef] [Green Version]
- Marangon, E.; Goldenberg, S.U.; Nagelkerken, I. Ocean warming increases availability of crustacean prey via riskier behavior. Behav. Ecol. 2019. [Google Scholar] [CrossRef]
- Åbjörnsson, K.; Hansson, L.A.; Brönmark, C. Responses of prey from habitats with different predator regimes: Local adaptation and heritability. Ecology 2004, 85, 1859–1866. [Google Scholar] [CrossRef] [Green Version]
- Välimäki, K.; Herczeg, G.; Merilä, J. Morphological anti-predator defences in the nine-spined stickleback: Constitutive, induced or both? Biol. J. Linn. Soc. 2012, 107, 854–866. [Google Scholar] [CrossRef] [Green Version]
- Reger, J.; Lind, M.I.; Robinson, M.R.; Beckerman, A.P. Predation drives local adaptation of phenotypic plasticity. Nature Ecol. Evol. 2018, 2, 100–107. [Google Scholar]
- Kaushik, N.K.; Hynes, H.B.N. The fate of dead leaves that fall into streams. Arch. Hydrobiol. 1971, 68, 465–515. [Google Scholar]
- Janssens, L.; Stoks, R. Predation risk causes oxidative damage in prey. Biol. Lett. 2013, 9, 20130350. [Google Scholar] [CrossRef] [PubMed]
- Helfman, G.S. Threat-sensitive predator avoidance in damselfish-trumpetfish interactions. Behav. Ecol. Sociobiol. 1989, 24, 47–58. [Google Scholar] [CrossRef]
- Monclús, R.; Palomares, F.; Tablado, Z.; Martínez-Fontúrbel, A.; Palme, R. Testing the threat-sensitive predator avoidance hypothesis: Physiological responses and predator pressure in wild rabbits. Oecologia 2009, 158, 615–623. [Google Scholar] [CrossRef]
- Kent, M.I.; Herbert-Read, J.E.; McDonald, G.; Wood, A.J.; Ward, A.J. Fine-scale behavioural adjustments of prey on a continuum of risk. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190448. [Google Scholar] [CrossRef] [Green Version]
- Hébert, M.; Versace, E.; Vallortigara, G. Inexperienced preys know when to flee or to freeze in front of a threat. Proc. Natl. Acad. Sci. USA 2019, 116, 22918–22920. [Google Scholar] [CrossRef]
- Chivers, D.P.; Mirza, R.S.; Bryer, P.J.; Kiesecker, J.M. Threat-sensitive predator avoidance by slimy sculpins: Understanding the importance of visual versus chemical information. Can. J. Zool. 2001, 79, 867–873. [Google Scholar] [CrossRef]










| Trait | Scaling Exponent for Fish Spring Populations | Scaling Exponent for Non-Fish Spring Populations |
|---|---|---|
| Somatic investment | ||
| Lab assimilation rate 3 | 0.671 (±0.117; 109) | 0.682 (±0.239; 105) |
| Fat mass (juveniles & adult males) 4 | 0.952 (±0.091; 76) | 1.114 (±0.059; 75) |
| Fat mass (juveniles & adult non-brooding females) 4 | 1.146 (±0.131; 77) | 1.337 (±0.072; 75) |
| Field assimilation rate 5 | 0.623 | 0.796 |
| Growth rate | 0.661 (±0.031; 178) | 0.798 (±0.030; 228) |
| Resting metabolic rate | 0.591 (±0.060; 336) | 0.760 (±0.080; 190) |
| Gill surface area | 0.620 (±0.084; 100) | 0.743 (±0.046; 85) |
| Reproductive investment | ||
| Total mass of eggs per brood | 1.184 (±0.211; 83) | 0.860 (±0.232; 114) |
| Egg number per brood | 1.034 (±0.217; 83) | 0.758 (±0.220; 114) |
| Individual egg mass | 0.151 (±0.096; 83) | 0.100 (±0.105; 114) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glazier, D.S.; Borrelli, J.J.; Hoffman, C.L. Effects of Fish Predators on the Mass-Related Energetics of a Keystone Freshwater Crustacean. Biology 2020, 9, 40. https://doi.org/10.3390/biology9030040
Glazier DS, Borrelli JJ, Hoffman CL. Effects of Fish Predators on the Mass-Related Energetics of a Keystone Freshwater Crustacean. Biology. 2020; 9(3):40. https://doi.org/10.3390/biology9030040
Chicago/Turabian StyleGlazier, Douglas S., Jonathan J. Borrelli, and Casandra L. Hoffman. 2020. "Effects of Fish Predators on the Mass-Related Energetics of a Keystone Freshwater Crustacean" Biology 9, no. 3: 40. https://doi.org/10.3390/biology9030040
APA StyleGlazier, D. S., Borrelli, J. J., & Hoffman, C. L. (2020). Effects of Fish Predators on the Mass-Related Energetics of a Keystone Freshwater Crustacean. Biology, 9(3), 40. https://doi.org/10.3390/biology9030040