A Perspective on Body Size and Abundance Relationships across Ecological Communities



Abstract
:1. Introduction
2. Case Studies
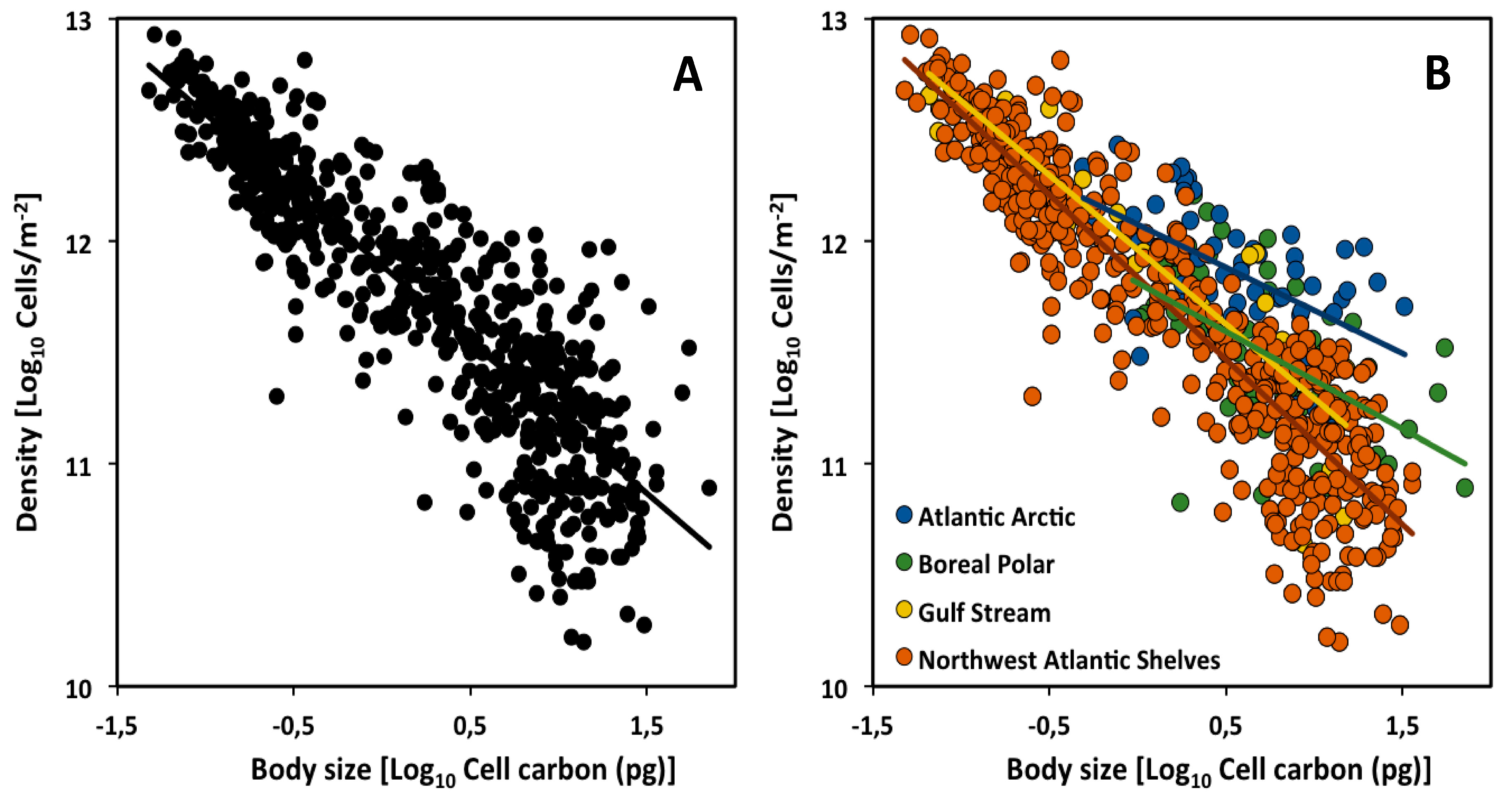
2.1. Across Climate Zones (Biogeographic Regions)
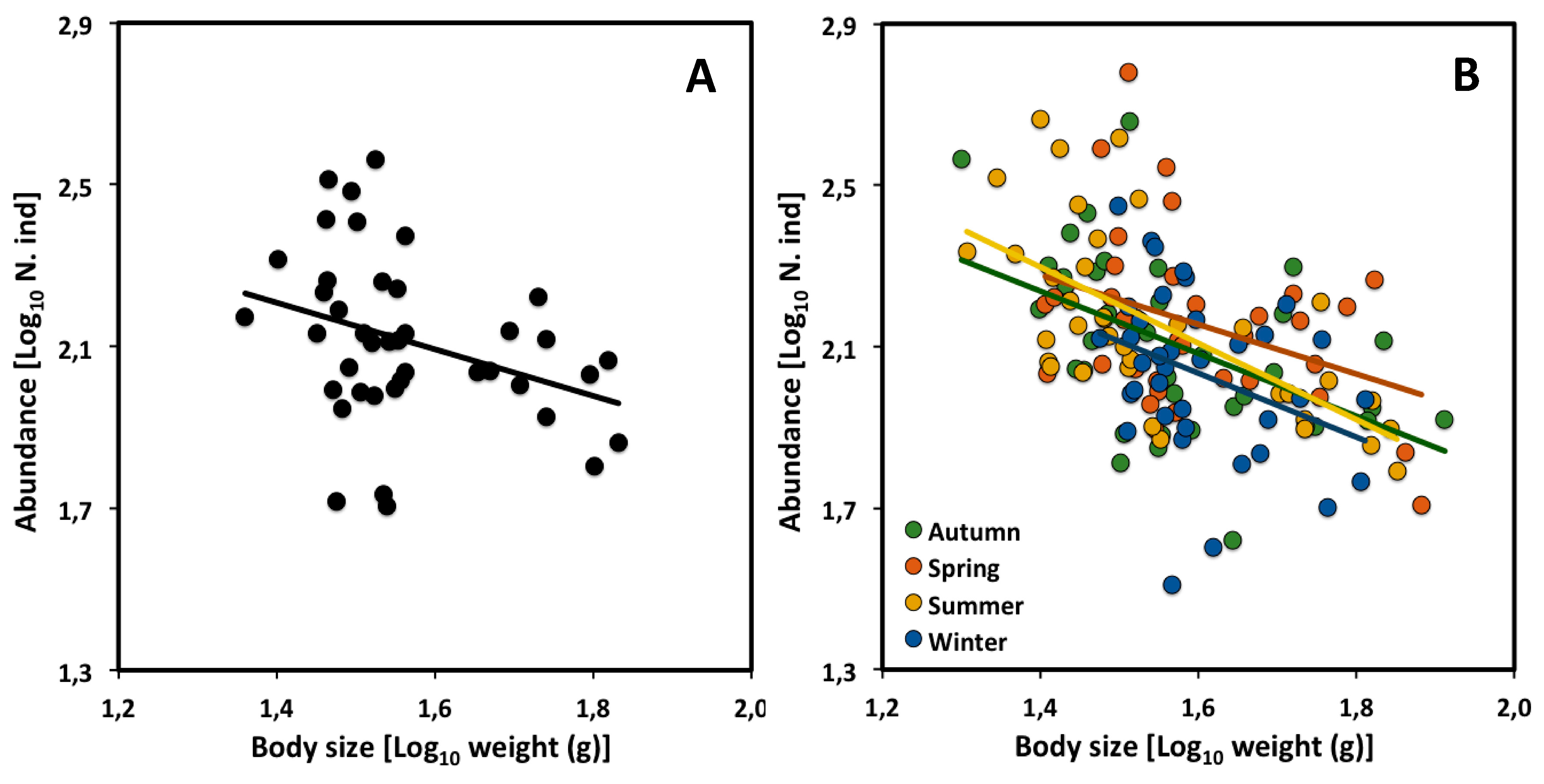
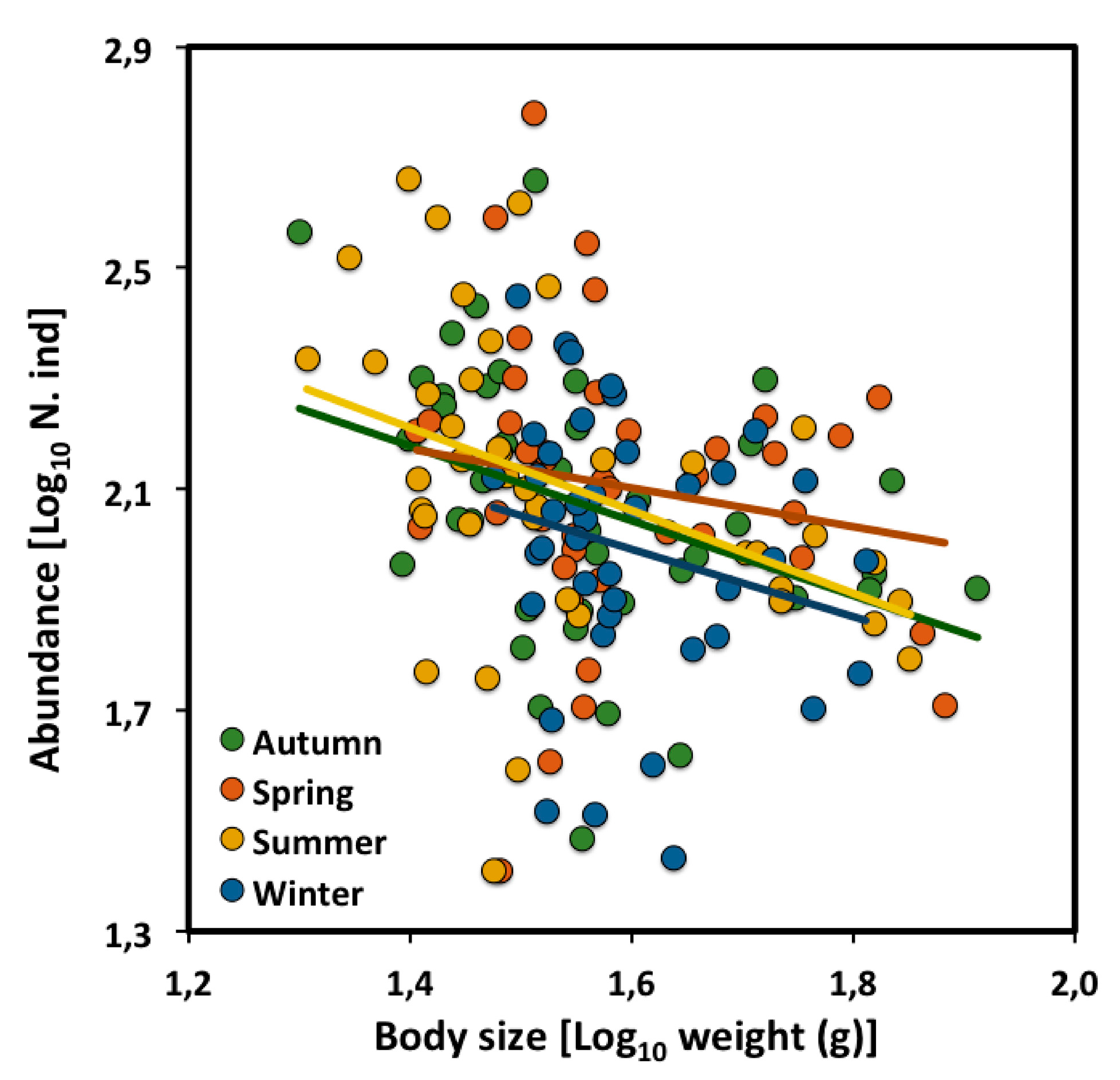
2.2. Across Seasons
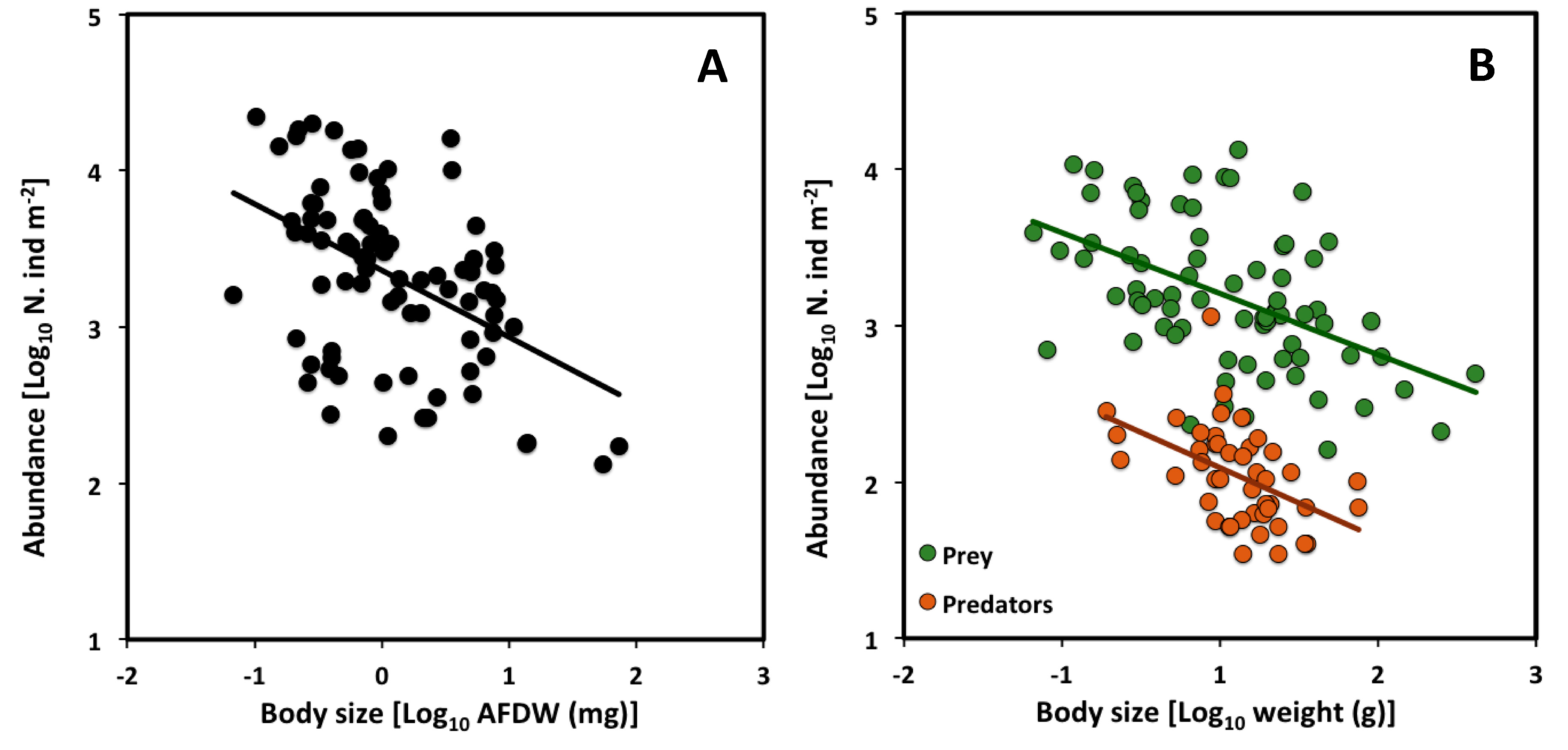
2.3. Across Trophic Levels
3. Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CCSR | Slope | Intercept | 95% CI | n | r2 | p |
|---|---|---|---|---|---|---|
| Autumn | −0.71 | 3.18 | −1.27 to −0.15 | 41 | 0.15 | * |
| Spring | −0.35 | 2.67 | −1.05 to −0.34 | 41 | 0.03 | ns |
| Summer | −0.75 | 3.26 | −1.28 to −0.22 | 41 | 0.17 | ** |
| Winter | −0.61 | 2.97 | −1.47 to −0.25 | 41 | 0.05 | ns |
| Seasons | p Value for Slope a | p Value for Intercept b | ||||||
|---|---|---|---|---|---|---|---|---|
| AU | SP | SU | WI | AU | SP | SU | WI | |
| Autumn (AU) | - | ns | ns | ns | - | ns | ns | ns |
| Spring (SP) | ns | - | ns | ns | ns | - | ns | ns |
| Summer (SU) | ns | ns | - | ns | ns | ns | - | ns |
| Winter (WI) | ns | ns | ns | - | ns | ns | ns | - |
References
- Li, B.L.; Wu, H.I.; Zou, G. Self-thinning rule: A causal interpretation from ecological field theory. Ecol. Model. 2000, 132, 167–173. [Google Scholar] [CrossRef]
- Yoda, K. Self-thinning in overcrowded pure stands under cultivated and natural conditions (Intraspecific competition among higher plants. XI). J. Inst. Polytech. Osaka City Univ. 1963, 14, 107–129. [Google Scholar]
- Westoby, M. The place of the self-thinning rule in population dynamics. Am. Nat. 1981, 118, 581–587. [Google Scholar] [CrossRef]
- Belgrano, A.; Allen, A.P.; Enquist, B.J.; Gillooly, J.F. Allometric scaling of maximum population density: A common rule for marine phytoplankton and terrestrial plants. Ecol. Lett. 2002, 5, 611–613. [Google Scholar] [CrossRef]
- Begon, M.; Firbank, L.; Wall, R. Is there a self-thinning rule for animal populations? Oikos 1986, 46, 122–124. [Google Scholar] [CrossRef]
- Latto, J. Evidence for a self-thinning rule in animals. Oikos 1994, 69, 531–534. [Google Scholar] [CrossRef]
- Fréchette, M.; Lefaivre, D. On self-thinning in animals. Oikos 1995, 73, 425–428. [Google Scholar] [CrossRef]
- Branch, G.M. Mechanisms reducing intraspecific competition in Patella spp.: Migration, differentiation and territorial behaviour. J. Anim. Ecol. 1975, 44, 575–600. [Google Scholar] [CrossRef]
- Hogarth, P.J. Population Density, Mean Weight, and the Nature of the “Thinning Line” in Semibalanus balanoides (L.) (Cirripedia Thoracica). Crustaceana 1985, 49, 215–218. [Google Scholar] [CrossRef]
- Hughes, R.N.; Griffiths, C.L. Self-thinning in barnacles and mussels: The geometry of packing. Am. Nat. 1988, 132, 484–491. [Google Scholar] [CrossRef]
- Frechette, M.; Lefaivre, D. Discriminating between food and space limitation in benthic suspension feeders using self-thinning relationships. Mar. Ecol. Prog. Ser. 1990, 65, 15–23. [Google Scholar] [CrossRef]
- Petraitis, P.S. The role of growth in maintaining spatial dominance by mussels (Mytilus edulis). Ecology 1995, 76, 1337–1346. [Google Scholar] [CrossRef]
- Guinez, R.; Castilla, J.C. A tridimensional self-thinning model for multilayered intertidal mussels. Am. Nat. 1999, 154, 341–357. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.W.; Kramer, D.L. Territory size as a predictor of the upper limit to population density of juvenile salmonids in streams. Can. J. Fish. Aquat. Sci. 1990, 47, 1724–1737. [Google Scholar] [CrossRef]
- Elliott, J.M. The self-thinning rule applied to juvenile sea trout, Salmo trutta. J. Anim. Ecol. 1993, 62, 371–379. [Google Scholar] [CrossRef]
- Bohlin, T.; Dellefors, C.; Faremo, U.; Johlander, A. The energetic equivalence hypothesis and the relation between population density and body size in stream-living salmonids. Am. Nat. 1994, 143, 478–493. [Google Scholar] [CrossRef]
- Armstrong, J.D.; Herbert, N.A. Homing movements of displaced stream-dwelling brown trout. J. Fish Biol. 1997, 50, 445–449. [Google Scholar] [CrossRef]
- Dunham, J.B.; Vinyard, G.L. Relationships between body mass, population density, and the self-thinning rule in stream-living salmonids. Can. J. Fish. Aquat. Sci. 1997, 54, 1025–1030. [Google Scholar] [CrossRef]
- White, E.P.; Ernest, S.M.; Kerkhoff, A.J.; Enquist, B.J. Relationships between body size and abundance in ecology. Trends Ecol. Evol. 2007, 22, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Damuth, J.D. Population density and body size in mammals. Nature 1981, 290, 699–700. [Google Scholar] [CrossRef]
- Damuth, J.D. Of size and abundance. Nature 1991, 351, 268–269. [Google Scholar] [CrossRef]
- Damuth, J.D. Population ecology: Common rules for animals and plants. Nature 1998, 395, 115–116. [Google Scholar] [CrossRef]
- Nee, S.; Read, A.F.; Greenwood, J.J.; Harvey, P.H. The relationship between abundance and body size in British birds. Nature 1991, 351, 312–313. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Enquist, B.J.; Brown, J.H.; West, G.B. Allometric scaling of plant energetics and population density. Nature 1998, 395, 163–165. [Google Scholar] [CrossRef]
- Li, W.K.W. Macroecological patterns of phytoplankton in the northwestern North Atlantic Ocean. Nature 2002, 419, 154–157. [Google Scholar] [CrossRef]
- Long, Z.T.; Morin, P.J. Effects of organism size and community composition on ecosystem functioning. Ecol. Lett. 2005, 8, 1271–1282. [Google Scholar] [CrossRef]
- Gjoni, V.; Cozzoli, F.; Rosati, I.; Basset, A. Size-density relationships: A cross-community approach to benthic macroinvertebrates in Mediterranean and Black Sea lagoons. Estuar. Coast. 2017, 40, 1142–1158. [Google Scholar] [CrossRef]
- Gjoni, V.; Basset, A. A cross-community approach to energy pathways across lagoon macroinvertebrate guilds. Estuar. Coast. 2018, 41, 2433–2446. [Google Scholar] [CrossRef]
- Gjoni, V.; Ghinis, S.; Pinna, M.; Mazzotta, L.; Marini, G.; Ciotti, M.; Rosati, I.; Vignes, F.; Arima, S.; Basset, A. Patterns of functional diversity of macroinvertebrates across three aquatic ecosystem types, NE Mediterranean. Mediterr. Mar. Sci. 2019, 20, 703–717. [Google Scholar] [CrossRef] [Green Version]
- Arim, M.; Berazategui, M.; Barreneche, J.M.; Ziegler, L.; Zarucki, M.; Abades, S.R. Determinants of density–body size scaling within food webs and tools for their detection. Adv. Ecol. Res. 2011, 45, 1–39. [Google Scholar]
- Meehan, T.D.; Jetz, W.; Brown, J.H. Energetic determinants of abundance in winter landbird communities. Ecol. Lett. 2004, 7, 532–537. [Google Scholar] [CrossRef]
- White, E.P.; Ernest, S.M.; Thibault, K.M. Trade-offs in community properties through time in a desert rodent community. Am. Nat. 2004, 164, 670–676. [Google Scholar] [CrossRef]
- Glazier, D.S. Beyond the ‘3/4−power law’: Variation in the intra−and interspecific scaling of metabolic rate in animals. Biol. Rev. 2005, 80, 611–662. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. A unifying explanation for diverse metabolic scaling in animals and plants. Biol. Rev. 2010, 85, 111–138. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. Metabolic scaling in complex living systems. Systems 2014, 2, 451–540. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S. Rediscovering and reviving old observations and explanations of metabolic scaling in living systems. Systems 2018, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Griffen, B.D.; Cannizzo, Z.J.; Gül, M.R. Ecological and evolutionary implications of allometric growth in stomach size of brachyuran crabs. PLoS ONE 2018, 13, e0207416. [Google Scholar] [CrossRef] [Green Version]
- Malerba, M.E.; Marshall, D.J. Size-abundance rules? Evolution changes scaling relationships between size, metabolism and demography. Ecol. Lett. 2019, 22, 1407–1416. [Google Scholar] [CrossRef]
- Agusti, S.; Kalff, J. The influence of growth conditions on the size dependence of maximal algal density and biomass. Limnol. Oceanogr. 1989, 34, 1104–1108. [Google Scholar] [CrossRef]
- Rodríguez, J. Some comments on the size-based structural analysis of the pelagic ecosystem. Sci. Mar. 1994, 58, 1–10. [Google Scholar]
- Li, W.K.W.; Glen Harrison, W.; Head, E.J. Coherent assembly of phytoplankton communities in diverse temperate ocean ecosystems. Proc. R. Soc. B−Biol. Sci. 2006, 273, 1953–1960. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, D. Effects of temperature on the size of aquatic ectotherms: Exceptions to the general rule. J. Therm. Biol. 1995, 20, 61–74. [Google Scholar] [CrossRef]
- Atkinson, D.; Ciotti, B.J.; Montagnes, D.J.X. Protists decrease in size linearly with temperature: Ca. 2.5% C− 1. Proc. R. Soc. B−Biol. Sci. 2003, 270, 2605–2611. [Google Scholar] [CrossRef] [Green Version]
- Morán, X.A.G.; López-Urrutia, Á.; Calvo-Díaz, A.; Li, W.K.W. Increasing importance of small phytoplankton in a warmer ocean. Glob. Chang. Biol. 2010, 16, 1137–1144. [Google Scholar] [CrossRef]
- Kenagy, G.J.; Bartholomew, G.A. Seasonal reproductive patterns in five coexisting California desert rodent species. Ecol. Monogr. 1985, 55, 371–397. [Google Scholar] [CrossRef]
- Zeng, Z.; Brown, J.H. Population ecology of a desert rodent: Dipodomys merriami in the Chihuahuan desert. Ecology 1987, 68, 656–665. [Google Scholar] [CrossRef]
- Waser, P.M.; Jones, W.T. Survival and reproductive effort in banner-tailed kangaroo rats. Ecology 1991, 72, 771–777. [Google Scholar] [CrossRef]
- Lindeman, R.L. The trophic-dynamic aspect of ecology. Ecology 1942, 23, 399–417. [Google Scholar] [CrossRef]
- Odum, E.P. Energy flow in ecosystems: A historical review. Am. Zool. 1968, 8, 11–18. [Google Scholar] [CrossRef]
- Merritt, R.W.; Cummins, K.W. Trophic Relationships of Macroinvertebrates. In Methods in Stream Ecology, 3rd ed.; Hauer, F.R., Lamberti, G.A., Eds.; Academic Press: London, UK, 2007; Volume 1, pp. 413–433. [Google Scholar]
- Cummins, K.W.; Merritt, R.W.; Andrade, P.C. The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in south Brazil. Stud. Neotrop. Fauna Environ. 2005, 40, 69–89. [Google Scholar] [CrossRef]
- Carlier, A.; Riera, P.; Amouroux, J.M.; Bodiou, J.Y.; Escoubeyrou, K.; Desmalades, M.; Caparros, J.; Grémare, A. A seasonal survey of the food web in the Lapalme lagoon (northwestern Mediterranean) assessed by carbon and nitrogen stable isotope analysis. Estuar. Coast. Mar. Sci. 2007, 73, 299–315. [Google Scholar] [CrossRef]
- Cohen, J.E.; Jonsson, T.; Carpenter, S.R. Ecological community description using the food web, species abundance, and body size. Proc. Natl. Acad. Sci. USA 2003, 100, 1781–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virnstein, R.W. The importance of predation by crabs and fishes on benthic infauna in Chesapeake Bay. Ecology 1977, 58, 1199–1217. [Google Scholar] [CrossRef]
- Peterson, C.H. Predation, Competitive Exclusion, and Diversity in the Soft Sediment Communities of Estuaries and Lagoon. In Ecological Processes in Coastal and Marine Systems; Livingston, R.J., Ed.; Plenum Publishing Co.: New York, NY, USA, 1979; pp. 233–264. [Google Scholar]
- Holland, A.F.; Mountford, N.K.; Hiegel, M.H.; Kaumeyer, K.R.; Mihursky, K.A. Influence of predation on infaunal abundance in upper Chesapeake Bay, USA. Mar. Biol. 1980, 57, 221–235. [Google Scholar] [CrossRef]
- Eggleston, D.B.; Lipcius, R.N.; Hines, A.H. Density-dependent predation by blue crabs upon infaunal clam species with contrasting distribution and abundance patterns. Mar. Ecol. Prog. Ser. 1992, 85, 55–68. [Google Scholar] [CrossRef]
- Glazier, D.S. Scaling of metabolic scaling within physical limits. Systems 2014, 2, 425–450. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J.; Hammond, S.T. On the interpretation of the normalization constant in the scaling equation. Front. Ecol. Evol. 2018, 6, 212. [Google Scholar] [CrossRef] [Green Version]



| CCSR | Slope | 95% CI | Intercept | n | r2 | p |
|---|---|---|---|---|---|---|
| All assemblages | −0.68 | −0.71 to −0.65 | 11.89 | 635 | 0.77 | *** |
| Atlantic Arctic | −0.39 | −0.53 to −0.24 | 12.07 | 59 | 0.33 | *** |
| Boreal Polar | −0.44 | −0.60 to −0.27 | 11.81 | 124 | 0.31 | *** |
| Gulf Stream | −0.67 | −0.77 to −0.71 | 11.96 | 31 | 0.77 | *** |
| NW Atlantic Shelves | −0.74 | −0.78 to −0.56 | 11.84 | 479 | 0.85 | *** |
| Climate Zone | p Value for Slope a | p Value for Intercept b | ||||||
|---|---|---|---|---|---|---|---|---|
| AA | BP | GS | NWAS | AA | BP | GS | NWAS | |
| Atlantic Arctic (AA) | - | ns | *** | *** | - | *** | - | - |
| Boreal Polar (BP) | ns | - | *** | *** | *** | - | - | - |
| Gulf Stream (GS) | *** | *** | - | ns | - | - | - | ** |
| NW Atlantic Shelves (NWAS) | *** | *** | ns | - | - | - | ** | - |
| CCSR | Slope | 95% CI | Intercept | n | r2 | p |
|---|---|---|---|---|---|---|
| All assemblages | −0.55 | −1.06 to −0.03 | 2.96 | 41 | 0.11 | * |
| Autumn | −0.77 | −1.24 to −0.31 | 3.32 | 38 | 0.25 | ** |
| Spring | −0.61 | −1.16 to −0.07 | 3.14 | 38 | 0.13 | * |
| Summer | −0.94 | −1.28 to −0.22 | 3.62 | 38 | 0.42 | *** |
| Winter | −0.79 | −1.47 to −0.25 | 3.30 | 38 | 0.12 | * |
| Seasons | p Value for Slope a | p Value for Intercept 1 b | ||||||
|---|---|---|---|---|---|---|---|---|
| AU | SP | SU | WI | AU | SP | SU | WI | |
| Autumn (AU) | - | ns | ns | ns | - | ns | ns | ns |
| Spring (SP) | ns | - | ns | ns | ns | - | ns | ** |
| Summer (SU) | ns | ns | - | ns | ns | ns | - | ns |
| Winter (WI) | ns | ns | ns | - | ns | ** | ns | - |
| CCSR | Slope | 95% CI | Intercept | n | r2 | p |
|---|---|---|---|---|---|---|
| All assemblages | −0.43 | −0.60 to −0.25 | 3.36 | 85 | 0.22 | *** |
| Prey | −0.39 | −0.53 to −0.24 | 3.40 | 75 | 0.25 | *** |
| Predators | −0.45 | −0.78 to −0.24 | 2.32 | 45 | 0.23 | *** |
| Trophic Level | p Value for Slope a | p Value for Intercept b | ||
|---|---|---|---|---|
| Prey | Predators | Prey | Predators | |
| Prey | - | ns | - | *** |
| Predators | ns | - | *** | - |
| Assemblages | N | Slope | 95% CI | Deviation from −3/4 | Reference |
|---|---|---|---|---|---|
| Phytoplankton | 656 | −0.78 | −0.74 to −0.811 | = | [1] |
| Algae, bacteria & protozoa | 20 | −0.35 | −0.01 to −0.71 1 | > | [27] |
| 20 | 0.36 | 0.00 to −0.72 2 | > | ||
| 20 | −1.15 | −0.96 to −1.34 3 | < | ||
| 20 | −1.34 | −1.01 to−1.67 4 | < | ||
| 20 | −1.05 | −1.21 to −0.89 5 | < | ||
| 20 | −1.02 | −1.16 to −0.88 6 | < | ||
| Macroinvertebrates | 158 | −0.27 | −0.41 to −0.131 | > | [28] |
| Macroinvertebrates | 75 | −0.35 | −0.55 to −0.23 7 | > | [29] |
| 68 | −0.35 | −0.62 to −0.08 8 | > | ||
| 65 | −0.58 | −0.78 to −0.37 9 | > | ||
| 64 | −0.44 | −0.58 to 0.31 10 | > | ||
| 45 | −0.45 | −0.67 to−0.24 11 | > | ||
| Macroinvertebrates | 32 | −0.36 | −0.67 to −0.06 | > | [30] |
| Amphibians, fishes | 18 | −0.64 | −1.00 to −0.28 | = | [31] |
| & macroinvertebrates | |||||
| Winter land birds | 285 | −1.00 | −1.43 to −0.57 | < | [32] |
| Desert rodents | 25 | −0.57 | −0.97 to −0.18 | = | [33] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gjoni, V.; Glazier, D.S. A Perspective on Body Size and Abundance Relationships across Ecological Communities. Biology 2020, 9, 42. https://doi.org/10.3390/biology9030042
Gjoni V, Glazier DS. A Perspective on Body Size and Abundance Relationships across Ecological Communities. Biology. 2020; 9(3):42. https://doi.org/10.3390/biology9030042
Chicago/Turabian StyleGjoni, Vojsava, and Douglas Stewart Glazier. 2020. "A Perspective on Body Size and Abundance Relationships across Ecological Communities" Biology 9, no. 3: 42. https://doi.org/10.3390/biology9030042
APA StyleGjoni, V., & Glazier, D. S. (2020). A Perspective on Body Size and Abundance Relationships across Ecological Communities. Biology, 9(3), 42. https://doi.org/10.3390/biology9030042