


Solid-State Fermentation for the Recovery of Phenolic Compounds from Agro-Wastes

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
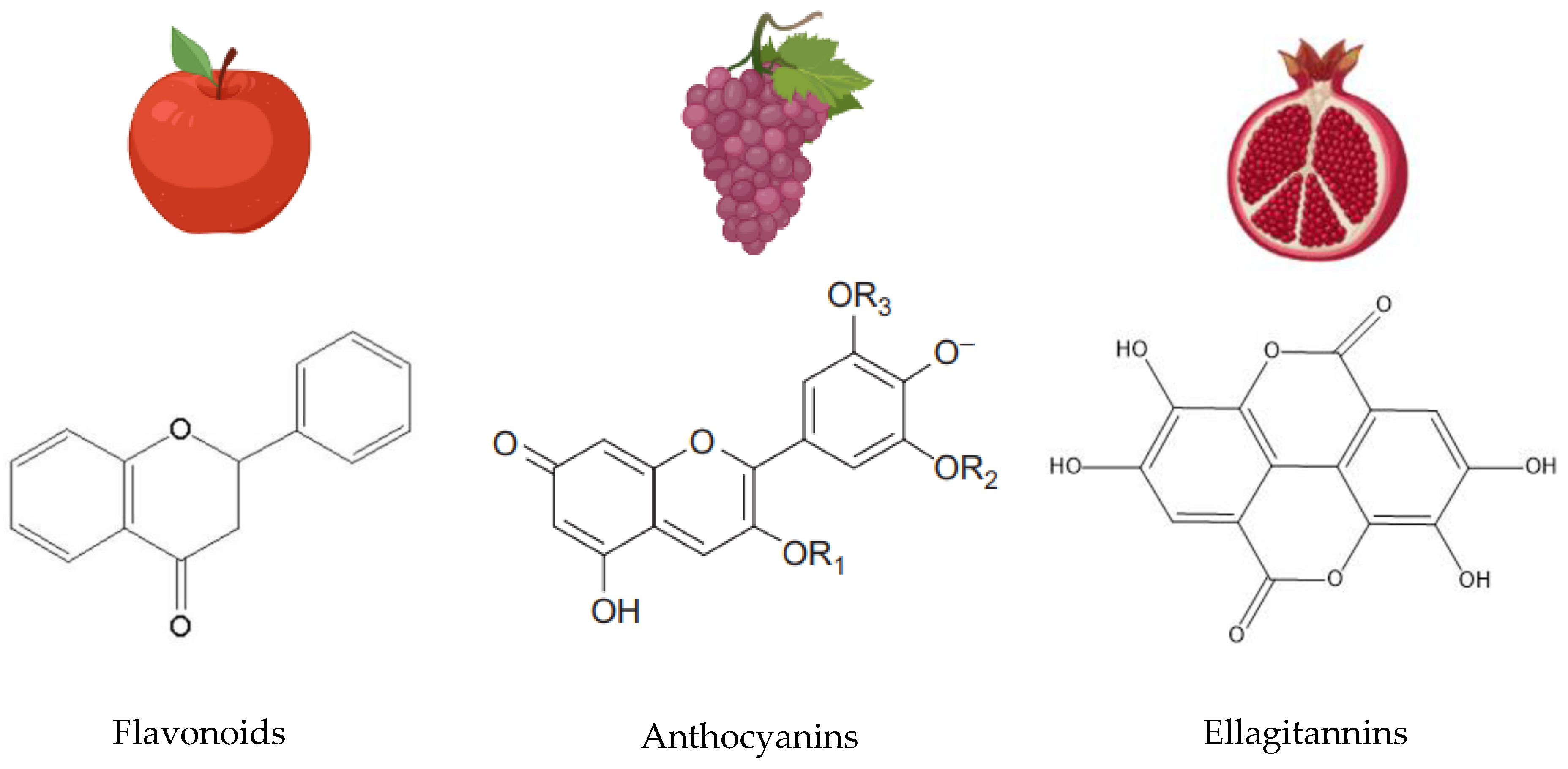
2. Polyphenol Chemistry
Hydrolysable and Condensed Polyphenols
3. Biological Activities of Phenolic Compounds
3.1. Polyphenols Activity against SARS-CoV-2
3.2. Antioxidant Activity of Phenolic Compounds
3.3. Antimicrobial Activity of Phenolic Compounds
3.4. Antiproliferative and Anticarcinogenic Activities of Phenolic Compounds
4. Solid-State Fermentation Extraction Effects on Phenolic Contents
4.1. Factors That Affect a SSF
4.1.1. Bioreactors
4.1.2. Temperature
4.1.3. Inoculum and Microorganism
Yeast
Filamentous Fungi
Bacteria
4.1.4. Moisture and Water Activity
4.1.5. pH
4.1.6. Substrate
4.1.7. Generalities of Natural Agro-Industrial Wastes Substrates Used in SSF
Castilla Rose
Mango
Grapes
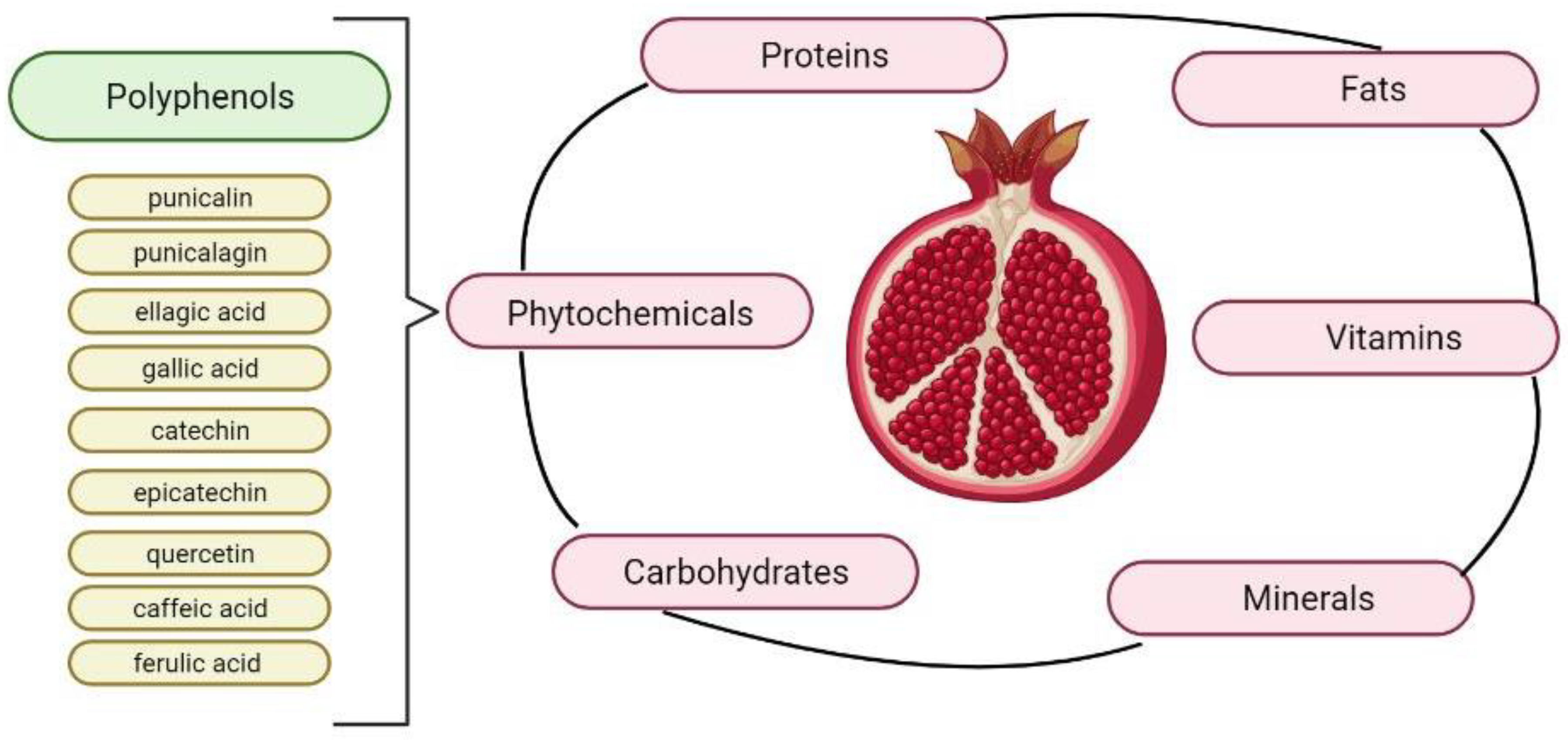
Pomegranate
Rambutan
5. Synthesis of Phenolic Compounds Recovered by SSF Assistant Extraction
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, J.; Gong, X.; Yang, M.; Zhang, C.; Li, M. Biosynthesis, Chemistry, and Pharmacology of Polyphenols from Chinese Salvia Species: A Review. Molecules 2019, 24, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepúlveda, L.; Wong-Paz, J.E.; Buenrostro-Figueroa, J.; Ascacio-Valdés, J.A.; Aguilera-Carbó, A.; Aguilar, C.N. Solid State Fermentation of Pomegranate Husk: Recovery of Ellagic Acid by SEC and Identification of Ellagitannins by HPLC/ESI/MS. Food Biosci. 2018, 22, 99–104. [Google Scholar] [CrossRef]
- Burton-Freeman, B.; Sandhu, A.; Edirisinghe, I. Anthocyanins. In Nutraceuticals; Gupta, R.C., Ed.; Elsevier: San Diego, CA, USA, 2016; pp. 489–500. [Google Scholar] [CrossRef]
- Torres-León, C.; Ramírez-Guzmán, N.; Ascacio-Valdés, J.; Serna-Cock, L.; dos Santos Correia, M.T.; Contreras-Esquivel, J.C.; Aguilar, C.N. Solid-State Fermentation with Aspergillus niger to Enhance the Phenolic Contents and Antioxidative Activity of Mexican Mango Seed: A Promising Source of Natural Antioxidants. Lwt 2019, 112, 108236. [Google Scholar] [CrossRef]
- Soccol, C.R.; da Costa, E.S.F.; Letti, L.A.J.; Karp, S.G.; Woiciechowski, A.L.; de Souza Vandenberghe, L.P. Recent Developments and Innovations in Solid State Fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- Sepúlveda, L.; Laredo-Alcalá, E.; Buenrostro-Figueroa, J.J.; Ascacio-Valdés, J.A.; Genisheva, Z.; Aguilar, C.; Teixeira, J. Ellagic Acid Production Using Polyphenols from Orange Peel Waste by Submerged Fermentation. Electron. J. Biotechnol. 2020, 43, 1–7. [Google Scholar] [CrossRef]
- Bentrad, N.; Hamida-Ferhat, A. Date Palm Fruit (Phoenix dactylifera): Nutritional Values and Potential Benefits on Health. In The Mediterranean Diet; Preedy, V.R., Watson, R.R., Eds.; Elsevier: San Diego, CA, USA, 2020; pp. 239–255. [Google Scholar] [CrossRef]
- Amarowicz, R.; Janiak, M. Hydrolysable Tannins. In Encyclopedia of Food Chemistry; Elsevier: Olsztyn, Poland, 2019; pp. 337–343. [Google Scholar] [CrossRef]
- Karas, D.; Ulrichová, J.; Valentová, K. Galloylation of Polyphenols Alters Their Biological Activity. Food Chem. Toxicol. 2017, 105, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Ascacio-Valdés, J.A.; Buenrostro, J.J.; De la Cruz, R.; Sepúlveda, L.; Aguilera, A.F.; Prado, A.; Contreras, J.C.; Rodríguez, R.; Aguilar, C.N. Fungal Biodegradation of Pomegranate Ellagitannins. J. Basic Microbiol. 2014, 54, 28–34. [Google Scholar] [CrossRef]
- Aguilar-Zarate, P.; Wong-Paz, J.E.; Buenrostro-Figueroa, J.J.; Ascacio, J.A.; Contreras-Esquivel, J.C.; Aguilar, C.N. Ellagitannins: Bioavailability, Purification and Biotechnological Degradation. Mini-Rev. Med. Chem. 2018, 18, 1244–1252. [Google Scholar] [CrossRef]
- Chojnacka, K.; Witek-Krowiak, A.; Skrzypczak, D.; Mikula, K.; Młynarz, P. Phytochemicals Containing Biologically Active Polyphenols as an Effective Agent against COVID-19-Inducing Coronavirus. J. Funct. Foods 2020, 73, 104146. [Google Scholar] [CrossRef]
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and Coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.K.; Osswald, H.L.; Prato, G. Recent Progress in the Development of HIV-1 Protease Inhibitors for the Treatment of HIV/AIDS. J. Med. Chem. 2016, 59, 5172–5208. [Google Scholar] [CrossRef] [Green Version]
- de Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent Insights into Emerging Coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Silva, R.F.M.; Pogačnik, L. Polyphenols from Food and Natural Products: Neuroprotection and Safety. Antioxidants 2020, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Quero, J.; Mármol, I.; Cerrada, E.; Rodríguez-Yoldi, M.J. Insight into the Potential Application of Polyphenol-Rich Dietary Intervention in Degenerative Disease Management. Food Funct. 2020, 11, 2805–2825. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; González-Paramás, A.M.; Oludemi, T.; Ayuda-Durán, B.; González-Manzano, S. Plant Phenolics as Functional Food Ingredients. In Advances in Food and Nutrition Research; Ferreira, I., Barros, L., Eds.; Elsevier: San Diego, CA, USA, 2019; pp. 183–257. [Google Scholar] [CrossRef]
- Paraiso, I.L.; Revel, J.S.; Stevens, J.F. Potential Use of Polyphenols in the Battle against COVID-19. Curr. Opin. Food Sci. 2020, 32, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sk, M.F.; Sonawane, A.; Kar, P.; Sadhukhan, S. Plant-Derived Natural Polyphenols as Potential Antiviral Drugs against SARS-CoV-2 via RNA-dependent RNA Polymerase (RdRp) Inhibition: An in-Silico Analysis. J. Biomol. Struct. Dyn. 2021, 39, 6249–6264. [Google Scholar] [CrossRef] [PubMed]
- Schettig, R.; Sears, T.; Klein, M.; Tan-Lim, R.; Matthias, R., Jr.; Aussems, C.; Hummel, M.; Sears, R.; Poteet, Z.; Warren, D.; et al. COVID-19 Patient with Multifocal Pneumonia and Respiratory Difficulty Resolved Quickly: Possible Antiviral and Anti-Inflammatory Benefits of Quercinex (Nebulized Quercetin-NAC) as Adjuvant. Adv. Infect. Dis. 2020, 10, 45–55. [Google Scholar] [CrossRef]
- Torres-León, C.; Aguirre-Joya, J.A.; Czaja, A.; Aguillón-Gutiérrez, D.R. In Silico Screening Bioaktiver Verbindungen Aus Mexikanischen Wüstenpflanzen zur Vorhersage Potenzieller Inhibitoren von SARSCoronavirus 2 (SARS-CoV-2). J. Med. Spice Plants 2020, 24, 153–156. [Google Scholar]
- Khalifa, I.; Zhu, W.; Mohammed, H.H.H.; Dutta, K.; Li, C. Tannins Inhibit SARS-CoV-2 through Binding with Catalytic Dyad Residues of 3CL pro: An in Silico Approach with 19 Structural Different Hydrolysable Tannins. J. Food Biochem. 2020, 44, e13432. [Google Scholar] [CrossRef] [PubMed]
- Guclu, G.; Kelebek, H.; Selli, S. Antioxidant Activity in Olive Oils. In Olives and Olive Oil in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 313–325. [Google Scholar] [CrossRef]
- Ghendov-Mosanu, A.; Cojocari, D.; Balan, G.; Patras, A.; Lung, I.; Soran, M.-L.; Opriş, O.; Cristea, E.; Sturza, R. Chemometric Optimization of Biologically Active Compounds Extraction from Grape Marc: Composition and Antimicrobial Activity. Molecules 2022, 27, 1610. [Google Scholar] [CrossRef] [PubMed]
- Fia, G.; Bucalossi, G.; Gori, C.; Borghini, F.; Zanoni, B. Recovery of Bioactive Compounds from Unripe Red Grapes (Cv. Sangiovese) through a Green Extraction. Foods 2020, 9, 566. [Google Scholar] [CrossRef] [PubMed]
- Grases, F.; Prieto, R.M.; Fernández-Cabot, R.A.; Costa-Bauzá, A.; Sánchez, A.M.; Prodanov, M. Effect of Consuming a Grape Seed Supplement with Abundant Phenolic Compounds on the Oxidative Status of Healthy Human Volunteers. Nutr. J. 2015, 14, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, E.; He, X.; Li, H.; Wu, S.; Li, S.; Deng, G. Biological Activities of Polyphenols from Grapes. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Elsevier: London, UK, 2014; pp. 47–58. [Google Scholar] [CrossRef] [Green Version]
- Ye Elmogahzy. Finished Fibrous Assemblies. In Engineering Textiles; Elsevier: Amsterdam, The Netherlands, 2020; pp. 275–298. [Google Scholar]
- Fei, P.; Ali, M.A.; Gong, S.; Sun, Q.; Bi, X.; Liu, S.; Guo, L. Antimicrobial Activity and Mechanism of Action of Olive Oil Polyphenols Extract against Cronobacter Sakazakii. Food Control 2018, 94, 289–294. [Google Scholar] [CrossRef]
- Torres-León, C.; Ventura-Sobrevilla, J.; Serna-Cock, L.; Ascacio-Valdés, J.A.; Contreras-Esquivel, J.; Aguilar, C.N. Pentagalloylglucose (PGG): A Valuable Phenolic Compound with Functional Properties. J. Funct. Foods 2017, 37, 176–189. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- WHO. Deaths by Cause, Age, Sex, by Country and by Region. Available online: https://www.who.int/data/global-health-estimates (accessed on 1 January 2023).
- Cui, Y.; He, J.; Yang, K.-L.; Zhou, K. Aerobic Acetone-Butanol-Isopropanol (ABI) Fermentation through a Co-Culture of Clostridium Beijerinckii G117 and Recombinant Bacillus Subtilis 1A1. Metab. Eng. Commun. 2020, 11, e00137. [Google Scholar] [CrossRef]
- Rady, I.; Mohamed, H.; Rady, M.; Siddiqui, I.A.; Mukhtar, H. Cancer Preventive and Therapeutic Effects of EGCG, the Major Polyphenol in Green Tea. Egypt. J. Basic Appl. Sci. 2018, 5, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Maya-Cano, D.A.; Arango-Varela, S.; Santa-Gonzalez, G.A. Phenolic Compounds of Blueberries (Vaccinium spp.) as a Protective Strategy against Skin Cell Damage Induced by ROS: A Review of Antioxidant Potential and Antiproliferative Capacity. Heliyon 2021, 7, e06297. [Google Scholar] [CrossRef]
- Srivastava, N.; Srivastava, M.; Ramteke, P.W.; Mishra, P.K. Solid-State Fermentation Strategy for Microbial Metabolites Production: An Overview. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 345–354. [Google Scholar] [CrossRef]
- Costa, J.A.V.; Treichel, H.; Kumar, V.; Pandey, A. Advances in Solid-State Fermentation. In Current Developments in Biotechnology and Bioengineering; Pandey, A., Larroche, C., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–17. [Google Scholar] [CrossRef]
- de Oliveira, J.; Rodrigues, C.; Vandenberghe, L.P.S.; Câmara, M.C.; Libardi, N.; Soccol, C.R. Gibberellic Acid Production by Different Fermentation Systems Using Citric Pulp as Substrate/Support. Biomed. Res. Int. 2017, 2017, 5191046. [Google Scholar] [CrossRef] [Green Version]
- Cerda, A.; Artola, A.; Barrena, R.; Font, X.; Gea, T.; Sánchez, A. Innovative production of bioproducts from organic waste through solid-state fermentation. Front. Sustain. Food Syst. 2019, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Mekoue Nguela, J.; Vernhet, A.; Julien-Ortiz, A.; Sieczkowski, N.; Mouret, J.-R. Effect of Grape Must Polyphenols on Yeast Metabolism during Alcoholic Fermentation. Food Res. Int. 2019, 121, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, F.; Carpena, M.; Fraga-Corral, M.; Echave, J.; Riaz Rajoka, M.S.; Barba, F.J.; Cao, H.; Xiao, J.; Prieto, M.A.; Simal-Gandara, J. Valorization of Kiwi Agricultural Waste and Industry By-Products by Recovering Bioactive Compounds and Applications as Food Additives: A Circular Economy Model. Food Chem. 2022, 370, 131315. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive Phenolic Compounds: Production and Extraction by Solid-State Fermentation. A Review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buenrostro-Figueroa, J.; Gutierrez-Sánchez, G.; Prado-Barragán, L.A.; Rodríguez-Herrera, R.; Aguilar-Zárate, P.; Sepúlveda, L.; Ascacio-Valdés, J.A.; Tafolla-Arellano, J.C.; Aguilar, C.N. Influence of Culture Conditions on Ellagitannase Expression and Fungal Ellagitannin Degradation. Bioresour. Technol. 2021, 337, 125462. [Google Scholar] [CrossRef] [PubMed]
- Yepes-Betancur, D.P.; Márquez-Cardozo, C.J.; Cadena-Chamorro, E.M.; Martinez-Saldarriaga, J.; Torres-León, C.; Ascacio-Valdes, A.; Aguilar, C.N. Solid-State Fermentation—Assisted Extraction of Bioactive Compounds from Hass Avocado Seeds. Food Bioprod. Process. 2021, 126, 155–163. [Google Scholar] [CrossRef]
- Jaibiba, P.; Naga Vignesh, S.; Hariharan, S. Working Principle of Typical Bioreactors. In Bioreactors; Singh, L., Yousuf, A., Mahapatra, D.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 145–173. [Google Scholar] [CrossRef]
- Singh, J.; Kaushik, N.; Biswas, S. Bioreactors–Technology & Design Analysis. Scitech J. 2014, 1, 27–36. [Google Scholar]
- Ghosh, B.; Bhattacharya, D.; Mukhopadway, M. Use of Fermentation Technology for Value-Added Industrial Research. In Principles and Applications of Fermentation Technology; Kuila, A., Sharma, V., Eds.; Wiley: Hoboken, NJ, USA, 2018; pp. 141–161. [Google Scholar]
- Finkler, A.T.J.; de Lima Luz, L.F.; Krieger, N.; Mitchell, D.A.; Jorge, L.M. A Model-Based Strategy for Scaling-up Traditional Packed-Bed Bioreactors for Solid-State Fermentation Based on Measurement of O2 Uptake Rates. Biochem. Eng. J. 2021, 166, 107854. [Google Scholar] [CrossRef]
- Arora, S.; Rani, R.; Ghosh, S. Bioreactors in Solid State Fermentation Technology: Design, Applications and Engineering Aspects. J. Biotechnol. 2018, 269, 16–34. [Google Scholar] [CrossRef]
- Pitol, L.O.; Finkler, A.T.J.; Dias, G.S.; Machado, A.S.; Zanin, G.M.; Mitchell, D.A.; Krieger, N. Optimization Studies to Develop a Low-Cost Medium for Production of the Lipases of Rhizopus Microsporus by Solid-State Fermentation and Scale-up of the Process to a Pilot Packed-Bed Bioreactor. Process Biochem. 2017, 62, 37–47. [Google Scholar] [CrossRef]
- Finkler, A.T.J.; Weber, M.Z.; Fuchs, G.A.; Scholz, L.A.; de Lima Luz, L.F., Jr.; Krieger, N.; Mitchell, D.A.; de Matos Jorge, L.M. Estimation of Heat and Mass Transfer Coefficients in a Pilot Packed-Bed Solid-State Fermentation Bioreactor. Chem. Eng. J. 2021, 408, 127246. [Google Scholar] [CrossRef]
- Mitchell, D.A.; Pitol, L.O.; Biz, A.; Finkler, A.T.J.; de Lima Luz, L.F.; Krieger, N. Design and Operation of a Pilot-Scale Packed-Bed Bioreactor for the Production of Enzymes by Solid-State Fermentation. In Solid State Fermentation. Advances in Biochemical; Steudler, S., Werner, A., Cheng, J., Eds.; Springer: Basel, Switzerland, 2019; pp. 27–50. [Google Scholar] [CrossRef]
- Finkler, A.T.J.; Biz, A.; Pitol, L.O.; Medina, B.S.; Luithardt, H.; Luz, L.F.D.L.; Krieger, N.; Mitchell, D.A. Intermittent Agitation Contributes to Uniformity across the Bed during Pectinase Production by Aspergillus niger Grown in Solid-State Fermentation in a Pilot-Scale Packed-Bed Bioreactor. Biochem. Eng. J. 2017, 121, 1–12. [Google Scholar] [CrossRef]
- Kucharczyk, K.; Tuszyński, T. The Effect of Temperature on Fermentation and Beer Volatiles at an Industrial Scale. J. Inst. Brew. 2018, 124, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Alenyorege, E.A.; Ouyang, N.; Zhou, A.; Ma, H. Simulated Natural and High Temperature Solid-State Fermentation of Soybean Meal: A Comparative Study Regarding Microorganisms, Functional Properties and Structural Characteristics. LWT 2022, 159, 113125. [Google Scholar] [CrossRef]
- Sood, S.; Singhal, R.; Bhat, S.; Kumar, A. Inoculum Preparation. In Comprehensive Biotechnology; Moo-Young, M., Ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 230–243. [Google Scholar] [CrossRef]
- Kondo, M.; Mulianda, R.; Matamura, M.; Shibata, T.; Mishima, T.; Jayanegara, A.; Isono, N. Validation of a Phenol-sulfuric Acid Method in a Microplate Format for the Quantification of Soluble Sugars in Ruminant Feeds. Anim. Sci. J. 2021, 92, e13530. [Google Scholar] [CrossRef] [PubMed]
- Dias, E.; Ebdon, J.; Taylor, H. Estimating the Concentration of Viral Pathogens and Indicator Organisms in the Final Effluent of Wastewater Treatment Processes Using Stochastic Modelling. Microb. Risk Anal. 2019, 11, 47–56. [Google Scholar] [CrossRef]
- Rawat, S. Food Spoilage: Microorganisms and Their Prevention. Asian J. Plant Sci. 2015, 5, 47–56. [Google Scholar]
- Perricone, M.; Gallo, M.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Yeasts. In The Microbiological Quality of Food; Becilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing: Duxford, UK, 2017; pp. 121–131. [Google Scholar] [CrossRef]
- Deak, T. (Ed.) Handbook of Food Spoilage Yeasts, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar] [CrossRef]
- Deak, T. (Ed.) Preservation: Inhibition and Inactivation of Yeasts. In Handbook of Food Spoilage Yeasts; Taylor & Francis: Boca Raton, FL, USA, 2007; p. 30. [Google Scholar] [CrossRef]
- Kumar, A.; Chadha, S.; Rath, D. CRISPR-Cas9 System for Functional Genomics of Filamentous Fungi: Applications and Challenges. In Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology; Sharma, V.K., Shah, M.P., Parmar, S., Kumar, A., Eds.; Elsevier: London, UK, 2021; pp. 541–576. [Google Scholar] [CrossRef]
- José Carlos, D.L.-M.; Leonardo, S.; Jesús, M.-C.; Paola, M.-R.; Alejandro, Z.-C.; Juan, A.-V.; Cristóbal Noé, A. Solid-State Fermentation with Aspergillus niger GH1 to Enhance Polyphenolic Content and Antioxidative Activity of Castilla Rose (Purshia plicata). Plants 2020, 9, 1518. [Google Scholar] [CrossRef] [PubMed]
- Meini, M.-R.; Cabezudo, I.; Galetto, C.S.; Romanini, D. Production of Grape Pomace Extracts with Enhanced Antioxidant and Prebiotic Activities through Solid-State Fermentation by Aspergillus niger and Aspergillus oryzae. Food Biosci. 2021, 42, 101168. [Google Scholar] [CrossRef]
- Akita, L.G.; Laudien, J.; Biney, C.; Akrong, M.O. A Baseline Study of Spatial Variability of Bacteria (Total Coliform, E. Coli, and Enterococcus spp.) as Biomarkers of Pollution in Ten Tropical Atlantic Beaches: Concern for Environmental and Public Health. Environ. Sci. Pollut. Res. 2021, 28, 50941–50965. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Bashar, M.A.E.; Rady, I.; El-Naggar, H.A.; Abd El-Maoula, L.M.; Mehany, A.B.M. Antiproliferative Activity, Proapoptotic Effect, and Cell Cycle Arrest in Human Cancer Cells of Some Marine Natural Product Extract. Oxid. Med. Cell. Longev. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Ong, A.; Lee, C.-L.K. Cooperative Metabolism in Mixed Culture Solid-State Fermentation. LWT 2021, 152, 112300. [Google Scholar] [CrossRef]
- Wang, J.; Barba, F.J.; Sørensen, J.C.; Frandsen, H.B.; Sørensen, S.; Olsen, K.; Orlien, V. The Role of Water in the Impact of High Pressure on the Myrosinase Activity and Glucosinolate Content in Seedlings from Brussels Sprouts. Innov. Food Sci. Emerg. Technol. 2019, 58, 102208. [Google Scholar] [CrossRef]
- Jin, G.; Zhu, Y.; Rinzema, A.; Wijffels, R.H.; Ge, X.; Xu, Y. Water Dynamics during Solid-State Fermentation by Aspergillus oryzae YH6. Bioresour. Technol. 2019, 277, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.A.; Arnaud, T.; Lu-Chau, T.A.; Fdz-Polanco, M.; Moreira, M.T.; Rivero, J.A.C. Review of Solid State Fermentation for Lignocellulolytic Enzyme Production: Challenges for Environmental Applications. Rev. Environ. Sci. Bio/Technol. 2016, 15, 31–46. [Google Scholar] [CrossRef]
- Sala, A.; Artola, A.; Sánchez, A.; Barrena, R. Rice Husk as a Source for Fungal Biopesticide Production by Solid-State Fermentation Using B. Bassiana and T. Harzianum. Bioresour. Technol. 2020, 296, 122322. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Kumar, D.; Varadwaj, P.; Tiwari, A. Water Activity and Biomass Estimation Using Digital Image Processing in Solid-State Fermentation. Bioresour. Technol. 2020, 308, 123277. [Google Scholar] [CrossRef]
- Johnson, G.J. Encyclopedia of Analytical Science (2nd Edition). Ref. Rev. 2005, 19, 38–39. [Google Scholar] [CrossRef]
- Mohd-Zaki, Z.; Bastidas-Oyanedel, J.; Lu, Y.; Hoelzle, R.; Pratt, S.; Slater, F.; Batstone, D. Influence of PH Regulation Mode in Glucose Fermentation on Product Selection and Process Stability. Microorganisms 2016, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temudo, M.F.; Kleerebezem, R.; van Loosdrecht, M. Influence of the PH on (Open) Mixed Culture Fermentation of Glucose: A Chemostat Study. Biotechnol. Bioeng. 2007, 98, 69–79. [Google Scholar] [CrossRef]
- Buenrostro-Figueroa, J.; Ascacio-Valdés, A.; Sepúlveda, L.; Prado-Barragán, A.; Aguilar-González, M.Á.; Aguilar, C.N. Ellagic Acid Production by Solid-State Fermentation Influenced by the Inert Solid Supports. Emirates J. Food Agric. 2018, 30, 750–757. [Google Scholar] [CrossRef]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-Industrial Wastes and Their Utilization Using Solid State Fermentation: A Review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Teles, A.S.C.; Chávez, D.W.H.; Oliveira, R.A.; Bon, E.P.S.; Terzi, S.C.; Souza, E.F.; Gottschalk, L.M.F.; Tonon, R.V. Use of Grape Pomace for the Production of Hydrolytic Enzymes by Solid-State Fermentation and Recovery of Its Bioactive Compounds. Food Res. Int. 2019, 120, 441–448. [Google Scholar] [CrossRef]
- Cerda-Cejudo, N.D.; Buenrostro-Figueroa, J.J.; Sepúlveda, L.; Torres-Leon, C.; Chávez-González, M.L.; Ascacio-Valdés, J.A.; Aguilar, C.N. Recovery of Ellagic Acid from Mexican Rambutan Peel by Solid-State Fermentation-Assisted Extraction. Food Bioprod. Process. 2022, 134, 86–94. [Google Scholar] [CrossRef]
- Mountain States Wholesale Nursey. Purshia plicata (syn. Cowania plicata) Antelope Bush. Available online: https://mswn.com/wp-content/uploads/info-sheets/Purshia-plicata.pdf (accessed on 9 September 2020).
- Ueno, H.; Shimada, A.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Matsumoto, Y.; Okamoto, M.; Fujiwara, Y.; Ishihara, T. Anti-Stress Effects of the Hydroalcoholic Extract of Rosa Gallica Officinalis in Mice. Heliyon 2019, 5, e01945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oak, P.; Deshpande, A.; Giri, A.; Gupta, V. Metabolomic Dynamics Reveals Oxidative Stress in Spongy Tissue Disorder During Ripening of Mangifera Indica L. Fruit. Metabolites 2019, 9, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall-Medrano, A.; Olivas-Aguirre, F.J.; Velderrain-Rodriguez, G.R.; González-Aguilar, A.; de la Rosa, L.A.; López-Díaz, J.A.; Álvarez-Parrilla, E. El Mango: Aspectos Agroindustriales, Valor Nutricional/Funcional y Efectos En La Salud. Nutricion 2015, 31, 67–75. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, L.; He, T.; Ou, Z.; Hu, Z.; Wang, K. Effects of Mango Peel Powder on Starch Digestion and Quality Characteristics of Bread. Int. J. Biol. Macromol. 2019, 140, 647–652. [Google Scholar] [CrossRef]
- Venkitasamy, C.; Zhao, L.; Zhang, R.; Pan, Z. Grapes. In Integrated Processing Technologies for Food and Agricultural By-Products; Pan, Z., Zhang, R., Zicari, S., Eds.; Elsevier: London, UK, 2019; pp. 133–163. [Google Scholar] [CrossRef]
- García-Lomillo, J.; González-SanJosé, M.L. Applications of Wine Pomace in the Food Industry: Approaches and Functions. Compr. Rev. Food Sci. Food Saf. 2017, 16, 3–22. [Google Scholar] [CrossRef]
- Dhineshkumar, V.; Ramasamy, D. Pomegranate Processing and Value Addition: A Review. Indian Hortic. J. 2016, 6, 1–12. [Google Scholar]
- Venkitasamy, C.; Zhao, L.; Zhang, R.; Pan, Z. Pomegranate. In Integrated Processing Technologies for Food and Agricultural By-Products; Pan, Z., Zhang, R., Zicari, S., Eds.; Elsevier: London, UK, 2019; pp. 181–216. [Google Scholar] [CrossRef]
- Bhinge, S.D.; Bhutkar, M.A.; Randive, D.S.; Wadkar, G.H.; Todkar, S.S.; Savali, A.S.; Chittapurkar, H.R. Screening of Hair Growth Promoting Activity of Punica granatum L. (Pomegranate) Leaves Extracts and Its Potential to Exhibit Antidandruff and Anti-Lice Effect. Heliyon 2021, 7, e06903. [Google Scholar] [CrossRef] [PubMed]
- Uzuner, S. Pomegranate. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Elsevier: Cambridge, MA, USA, 2020; pp. 549–563. [Google Scholar]
- USDA. National Nutrient Database for Standard Reference. Available online: https://fdc.nal.usda.gov (accessed on 28 April 2014).
- Li, W.; Zeng, J.; Shao, Y. Rambutãn—Nephelium lappaceum. In Exotic Fruits; Rodrigues, S., Silva, E.d.O., de Brito, E.S., Eds.; Elsevier: London, UK, 2018; pp. 369–375. [Google Scholar] [CrossRef]
- Akhtar, M.T.; Ismail, S.N.; Shaari, K. Rambutan (Nephelium lappaceum L.). In Fruit and Vegetable Phytochemicals; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 1227–1234. [Google Scholar] [CrossRef]
- Olmo, A. Estadísticas de Producción de Rambután en México. Available online: https://blogagricultura.com/estadisticas-rambutan-mexico/#:~:text=En2020laproducciónde,porhectárea(-2.5%25) (accessed on 1 January 2023).
- Sekar, M. Rambutan Fruits Extract in Aging Skin. In Aging; Preedy, V.R., Patel, V.B., Eds.; Elsevier: London, UK, 2020; pp. 303–307. [Google Scholar] [CrossRef]
- Lourith, N.; Kanlayavattanakul, M.; Chaikul, P.; Chansriniyom, C.; Bunwatcharaphansakun, P. In Vitro and Cellular Activities of the Selected Fruits Residues for Skin Aging Treatment. An. Acad. Bras. Cienc. 2017, 89 (Suppl. S1), 577–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, C.; Ascacio-Valdés, J.; De la Garza, H.; Wong-Paz, J.; Aguilar, C.N.; Martínez-Ávila, G.C.; Castro-López, C.; Aguilera-Carbó, A. Polyphenolic Content, in Vitro Antioxidant Activity and Chemical Composition of Extract from Nephelium lappaceum L. (Mexican Rambutan) Husk. Asian Pac. J. Trop. Med. 2017, 10, 1201–1205. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Ma, Q.; Guo, Y.; Sun, L. Protective Effects of Rambutan (Nephelium lappaceum) Peel Phenolics on H 2 O 2 -Induced Oxidative Damages in HepG2 Cells and d -Galactose-Induced Aging Mice. Food Chem. Toxicol. 2017, 108, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Flores, A.; Saenz-Galindo, A.; Ascacio-Valdés, J. Aprovechamiento de La Cáscara de Rambután (Nephelium lappaceum) Como Fuente de Compuestos Antioxidantes. In Colección Memorias de los Congresos de la Sociedad Química de México; Química de Productos Naturales(QPNT): Puerto Vallarta, Mexico, 2017; pp. 6–9. [Google Scholar]
- Perera, A.; Appleton, D.; Ying, L.H.; Elendran, S.; Palanisamy, U.D. Large Scale Purification of Geraniin from Nephelium lappaceum Rind Waste Using Reverse-Phase Chromatography. Sep. Purif. Technol. 2012, 98, 145–149. [Google Scholar] [CrossRef]
- Thitilertdecha, N.; Teerawutgulrag, A.; Kilburn, J.D.; Rakariyatham, N. Identification of Major Phenolic Compounds from Nephelium lappaceum L. and Their Antioxidant Activities. Molecules 2010, 15, 1453–1465. [Google Scholar] [CrossRef] [Green Version]
- Monrroy, M.; Araúz, O.; García, J.R. Active Compound Identification in Extracts of N. Lappaceum Peel and Evaluation of Antioxidant Capacity. J. Chem. 2020, 2020, 4301891. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Zhang, H.; Zhuang, Y. Preparation of Free, Soluble Conjugate, and Insoluble-Bound Phenolic Compounds from Peels of Rambutan (Nephelium lappaceum) and Evaluation of Antioxidant Activities in Vitro. J. Food Sci. 2012, 77, 198–204. [Google Scholar] [CrossRef]
- Mingshu, L.; Kai, Y.; Qiang, H.; Dongying, J. Biodegradation of Gallotannins and Ellagitannins. J. Basic Microbiol. 2006, 46, 68–84. [Google Scholar] [CrossRef]
- Huang, W.; Niu, H.; Gong, G.H.; Lu, Y.R.; Li, Z.S.; Li, H. Individual and Combined Effects of Physicochemical Parameters on Ellagitannin Acyl Hydrolase and Ellagic Acid Production from Ellagitannin by Aspergillus oryzae. Bioprocess Biosyst. Eng. 2007, 30, 281–288. [Google Scholar] [CrossRef]
- de la Cruz, R.; Ascacio, J.A.; Buenrostro, J.; Sepúlveda, L.; Rodríguez, R.; Prado-Barragán, A.; Contreras, J.C.; Aguilera, A.; Aguilar, C.N. Optimization of Ellagitannase Production by Aspergillus niger GH1 by Solid-State Fermentation. Prep. Biochem. Biotechnol. 2015, 45, 617–631. [Google Scholar] [CrossRef]
- Vilhelmova-Ilieva, N.; Galabov, A.S.; Mileva, M. Tannins as Antiviral Agents. In Tannins—Structural Properties, Biological Properties and Current Knowledge; IntechOpen: London, UK, 2020; pp. 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sepúlveda, L.; Aguilera-Carbó, A.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R.; Martínez-Hernández, J.L.; Aguilar, C.N. Optimization of Ellagic Acid Accumulation by Aspergillus niger GH1 in Solid State Culture Using Pomegranate Shell Powder as a Support. Process Biochem. 2012, 47, 2199–2203. [Google Scholar] [CrossRef]
- Chávez-González, M.L.; Guyot, S.; Rodríguez-Herrera, R.; Prado-Barragán, A.; Aguilar, C.N. Production Profiles of Phenolics from Fungal Tannic Acid Biodegradation in Submerged and Solid-State Fermentation. Process Biochem. 2014, 49, 541–546. [Google Scholar] [CrossRef]
- Ascacio-Valdés, J.A.; Aguilera-Carbó, A.F.; Buenrostro, J.J.; Prado-Barragán, A.; Rodríguez-Herrera, R.; Aguilar, C.N. The Complete Biodegradation Pathway of Ellagitannins by Aspergillus niger in Solid-State Fermentation. J. Basic Microbiol. 2016, 56, 329–336. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Substrate | Microorganism | Reactors | Conditions of Fermentation | Polyphenolic Compounds Recovered | References |
|---|---|---|---|---|---|
| Pomegranate peel | Aspergillus niger PSH | Tray reactor (40 × 30 × 6 cm) | 2 × 107 spores/g at 30 °C for 18 h | Pullicalagin, punicalin, ellagic acid | [7] |
| Mango Ataulfo seed | Aspergillus niger GH1 | Petri dishes | 2 × 107 spores/g at 30 °C for | Gallic acid, ellagic acid | [5] |
| Grape pomace and wheat bran | Aspergillus niger 3T5B8 | Erlenmeyer flasks (125 mL) | 107 spores at 37 °C kinetic until 96 h | Ellagitannins, anthocyanins, proanthocyanidins | [81] |
| Rambutan peel | Aspergillus niger GH1 | Polypropylene flask (five cubic centimeters) | 2 × 107 spores/g at 25 °C for 24 h | Ellagic acid | [82] |
| Castilla Rose | Aspergillus niger GH1 | Erlenmeyer flasks (250 mL) | 2 × 106 esp/g at 25 °C for 24 h | Ellagic acid, Catechin, Epicatequin, Kaempferol 3,7-O-diglucoside, | [66] |
| Group | Structure | Compounds | Molecular Weight (g/mol) | References |
|---|---|---|---|---|
| Ellagitannin |  | Ellagic acid | 302 | [100,101,102] |
| Ellagitannin |  | Gerannin | 952 | [100,101,103,104] |
| Ellagitannin |  | Corilagin | 634 | [100,101,102,104,105] |
| Flavonoids |  | Catechin | 289 | [101,106] |
| Hydroxybenzoic acid |  | Caffeic acid | 180 | [106] |
| Hydroxybenzoic acid |  | Syringic acid | 197 | [101,106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerda-Cejudo, N.D.; Buenrostro-Figueroa, J.J.; Sepúlveda-Torre, L.; Torres-León, C.; Chávez-González, M.L.; Ascacio-Valdés, J.A.; Aguilar, C.N. Solid-State Fermentation for the Recovery of Phenolic Compounds from Agro-Wastes. Resources 2023, 12, 36. https://doi.org/10.3390/resources12030036
Cerda-Cejudo ND, Buenrostro-Figueroa JJ, Sepúlveda-Torre L, Torres-León C, Chávez-González ML, Ascacio-Valdés JA, Aguilar CN. Solid-State Fermentation for the Recovery of Phenolic Compounds from Agro-Wastes. Resources. 2023; 12(3):36. https://doi.org/10.3390/resources12030036
Chicago/Turabian StyleCerda-Cejudo, Nadia D., José J. Buenrostro-Figueroa, Leonardo Sepúlveda-Torre, Cristian Torres-León, Mónica L. Chávez-González, Juan A. Ascacio-Valdés, and Cristóbal N. Aguilar. 2023. "Solid-State Fermentation for the Recovery of Phenolic Compounds from Agro-Wastes" Resources 12, no. 3: 36. https://doi.org/10.3390/resources12030036
APA StyleCerda-Cejudo, N. D., Buenrostro-Figueroa, J. J., Sepúlveda-Torre, L., Torres-León, C., Chávez-González, M. L., Ascacio-Valdés, J. A., & Aguilar, C. N. (2023). Solid-State Fermentation for the Recovery of Phenolic Compounds from Agro-Wastes. Resources, 12(3), 36. https://doi.org/10.3390/resources12030036