
Aboveground Biomass and Carbon Storage in Mangrove Forests in Southeastern Mexico

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Area
2.2. Sampling Desing
2.3. Determination of Tree Density
2.4. Determination of Aboveground Biomass
2.5. Determination of Carbon and CO2 Equivalent
3. Results
3.1. Tree Density
3.2. Woodland Diameter
3.3. Aboveground Biomass
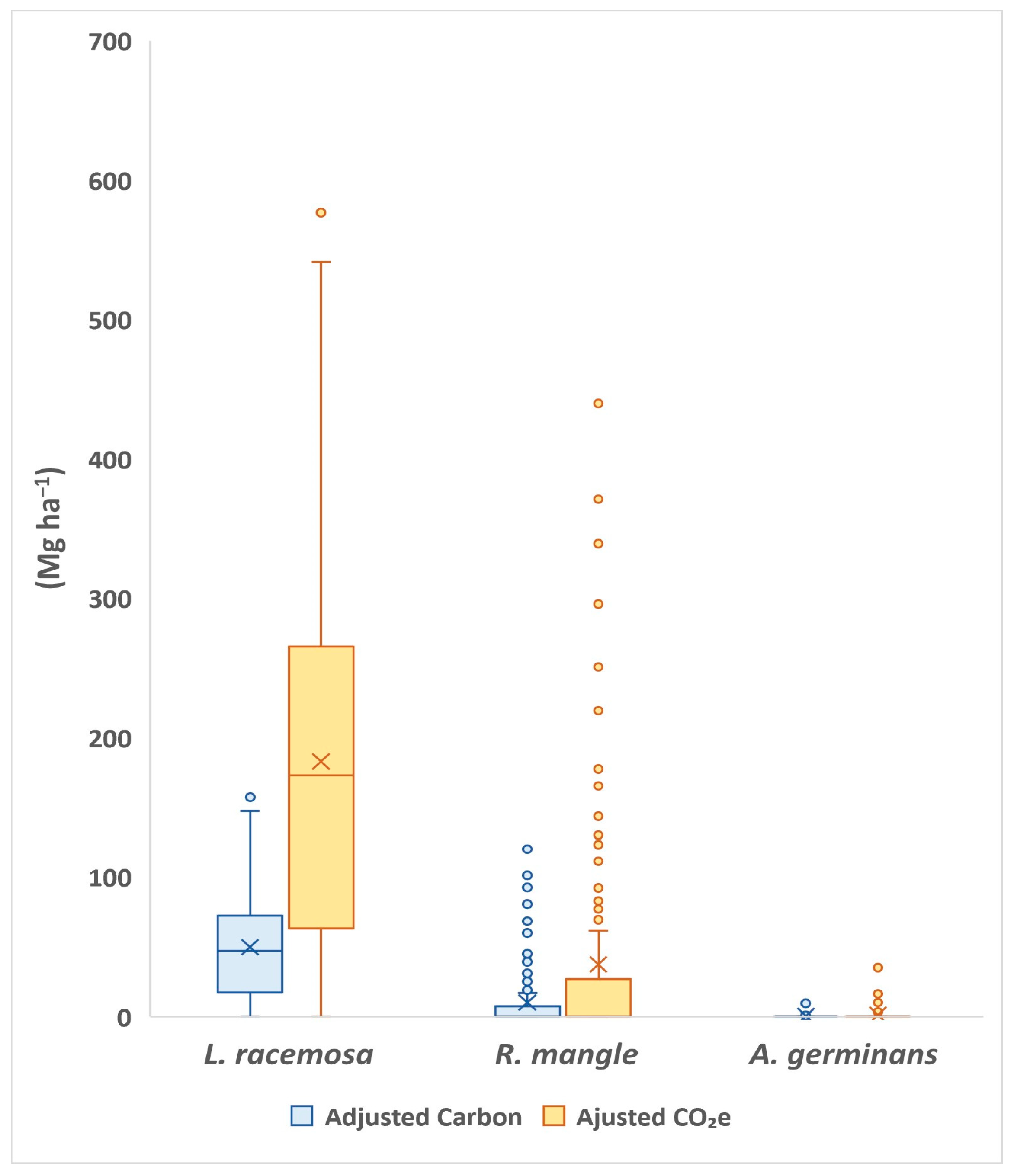
3.4. Aboveground Carbon
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bautista-Olivas, A.L.; Mendoza-Cariño, M.; Rodríguez, J.C.; Colado-Amador, C.E.; Robles-Zazueta, C.A.; Meling-López, A.E. Above-ground biomass and carbon sequestration in mangroves in the arid area of the northwest of Mexico: Bahía del Tóbari and Estero El Sargento, Sonora. Rev. Chapingo Ser. Cienc. For. Ambient. 2018, 24, 387–403. [Google Scholar] [CrossRef]
- Comisión para la Cooperación Ambiental. Carbono Azul en América del Norte: Evaluación de la Distribución de los Lechos de Pasto Marino, Marismas y Manglares, y su Papel Como Sumideros de Carbono; Comisión para la Cooperación Ambiental: Montreal, QC, Canadá, 2016; ISBN 978-2-89700-141-4. [Google Scholar]
- Jennerjahn, T.C. Relevance and magnitude of ‘Blue Carbon’ storage in mangrove sediments: Carbon accumulation rates vs. stocks, sources vs. Sinks. Estuar. Coast. Shelf Sci. 2021, 248, 107027. [Google Scholar] [CrossRef]
- Inoue, T. Carbon Sequestration in Mangroves. In Blue Carbon in Shallow Coastal Ecosystems; Kuwae, T., Hori, M., Eds.; Springer: Singapore, 2019; pp. 73–99. [Google Scholar] [CrossRef]
- Twilley, R.R.; Castañeda-Moya, E.; Rivera-Monroy, V.H.; Rovai, A. Productivity and Carbon Dynamics in Mangrove Wetlands. In Mangrove Ecosystems: A Global Biogeographic Perspective; Rivera-Monroy, V., Lee, S., Kristensen, E., Twilley, R., Eds.; Springer: Cham, Switzerland, 2017; pp. 113–162. [Google Scholar] [CrossRef]
- Thorhaug, A.; Gallagher, J.B.; Kiswara, W.; Prathep, A.; Huang, X.; Yap, T.K.; Dorward, S.; Berlyn, G. Coastal and estuarine blue carbon stocks in the greater Southeast Asia region: Seagrasses and mangroves per nation and sum of total. Mar. Pollut. Bull. 2020, 160, 111168. [Google Scholar] [CrossRef]
- Thorhaug, A.L.; Poulos, H.M.; López-Portillo, J.; Barr, J.; Lara-Domínguez, A.L.; Ku, T.C.; Berlyn, G.P. Gulf of Mexico estuarine blue carbon stock, extent and flux: Mangroves, marshes, and seagrasses: A North American hotspot. Sci. Total Environ. 2019, 653, 1253–1261. [Google Scholar] [CrossRef]
- Velázquez-Salazar, S.; Rodríguez-Zúñiga, M.T.; Alcántara-Maya, J.A.; Villeda-Chávez, E.; Valderrama-Landeros, L.; Troche-Souza, C.; Vázquez-Balderas, B.; Pérez-Espinosa, I.; Cruz-López, M.I.; Ressl, R.; et al. Manglares de México. Actualización y Análisis de los Datos 2020; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, México, 2021; ISBN 978-607-8570-50-8. [Google Scholar]
- Kauffman, J.B.; Donato, D.; Adame, M.F. Protocolo Para la Medición, Monitoreo y Reporte de la Estructura, Biomasa y Reservas de Carbono de los Manglares. Documento de Trabajo 117; Cifor: Bogor, Indonesia, 2013. [Google Scholar] [CrossRef]
- Velázquez-Pérez, C.; Tovilla-Hernández, C.; Romero-Berny, E.I.; De Jesús-Navarrete, A. Estructura del manglar y su influencia en el almacén de carbono en la Reserva La Encrucijada, Chiapas, México. Madera Bosques 2019, 25, e2531885. [Google Scholar] [CrossRef]
- Ajonina, G.N.; Kairo, J.; Grimsditch, G.; Sembres, T.; Chuyong, G.; Diyouke, E. Assessment of Mangrove Carbon Stocks in Cameroon, Gabon, the Republic of Congo (RoC) and the Democratic Republic of Congo (DRC) Including their Potential for Reducing Emissions from Deforestation and Forest Degradation (REDD+). In The Land/Ocean Interactions in the Coastal Zone of West and Central Africa. Estuaries of the World; Diop, S., Barusseau, J.P., Descamps, C., Eds.; Springer: Cham, Switzerland, 2014; pp. 177–189. [Google Scholar] [CrossRef]
- Harishma, K.M.; Sandeep, S.; Sreekumar, V.B. Biomass and carbon stocks in mangrove ecosystems of Kerala, southwest coast of India. Ecol. Process. 2020, 9, 31. [Google Scholar] [CrossRef]
- da Motta Portillo, J.T.; Londe, V.; Moreira, F.W.A. Aboveground biomass and carbon stock are related with soil humidity in a mangrove at the Piraquê-Açu River, Southeastern Brazil. J. Coast. Conserv. 2017, 21, 139–144. [Google Scholar] [CrossRef]
- Sjögersten, S.; Barreda-Bautista, B.; Brown, C.; Boyd, D.; López-Rosas, H.; Hernández, E.; Monroy, R.; Rincón, M.; Vane, C.; Moss-Hayes, V.; et al. Coastal wetland ecosystems deliver large carbon stocks in tropical Mexico. Geoderma 2021, 403, 115173. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; Chan-Keb, C.A.; Chávez-Barrera, J.; Osti-Sáenz, J.; Expósito-Díaz, G.; Alonso-Campos, V.A.; Muñiz-Salazar, R.; Ruiz-Fernández, A.C.; Pérez-Bernal, L.H.; Sánchez-Cabeza, J.A.; et al. Reserva de carbono en un ecosistema de manglar al norte de México: Cambios ambientales durante 35 años. Rev. Mex. Biodivers. 2020, 91, e912910. [Google Scholar] [CrossRef]
- Hutchison, J.; Manica, A.; Swetnam, R.; Balmfold, A.; Spalding, M. Predicting global patterns in mangrove forest biomass. Conserv. Lett. 2014, 7, 233–240. [Google Scholar] [CrossRef]
- Medeiros, T.C.C.; Sampaio, E.V.S.B. Allometry of aboveground biomasses in mangrove species in Itamaracá, Pernambuco, Brazil. Wetl. Ecol. Manag. 2008, 16, 323–330. [Google Scholar] [CrossRef]
- Fu, W.; Wu, Y. Estimation of aboveground biomass of different mangrove trees based on canopy diameter and tree height. Procedia Environ. Sci. 2011, 10, 2189–2194. [Google Scholar] [CrossRef]
- Rozainah, M.Z.; Sofawi, A.B.; Joharee, N.A.; Ahmad, Z.P. Stand structure and biomass estimation in the Klang Islands Mangrove Forest, Peninsular Malaysia. Environ. Earth Sci. 2018, 77, 486. [Google Scholar] [CrossRef]
- Cooray, P.L.I.G.M.; Kodikara, K.A.S.; Kumara, M.P.; Jayasinghe, U.I.; Madarasinghe, S.K.; Dahdouh-Guebas, F.; Gorman, D.; Huxham, M.; Jayatissa, L.P. Climate and intertidal zonation drive variability in the carbon stocks of Sri Lankan mangrove forests. Geoderma 2021, 389, 114929. [Google Scholar] [CrossRef]
- De la Peña, A.; Rojas, C.A.; De la Peña, M. Valoración económica del manglar por el almacenamiento de carbono, Ciénaga Grande de Santa Marta. Clío América 2010, 4, 133–150. [Google Scholar]
- Tang, W.; Feng, W.; Jia, M.; Shi, J.; Zuo, H.; Trettin, C.C. The assessment of mangrove biomass and carbon in West Africa: A spatially explicit analytical framework. Wetl. Ecol. Manag. 2016, 24, 153–171. [Google Scholar] [CrossRef]
- Estrada, G.C.D.; Soares, M.L.G.; Santos, D.M.C.; Fernandez, V.; de Almeida, P.M.M.; Estevam, M.R.M.; Machado, M.R.O. Allometric models for aboveground biomass estimation of the mangrove Avicennia schaueriana. Hydrobiologia 2014, 734, 171–185. [Google Scholar] [CrossRef]
- Jakovac, C.C.; Latawiec, A.E.; Lacerda, E.; Lucas, I.L.; Korys, K.A.; Iribarrem, A.; Malaguti, G.A.; Turner, R.K.; Luisetti, T.; Strassburg, B.B.N. Costs and Carbon Benefits of Mangrove Conservation and Restoration: A Global Analysis. Ecol. Econ. 2020, 176, 106758. [Google Scholar] [CrossRef]
- Komiyama, A.; Poungparn, S.; Kato, S. Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Yuen, J.Q.; Fung, T.; Ziegler, A.D. Review of allometric equations for major land covers in SE Asia: Uncertainty and implications for above- and below-ground carbon estimates. For. Ecol. Manag. 2016, 360, 323–340. [Google Scholar] [CrossRef]
- Henry, M.; Picard, N.; Trotta, C.; Manlay, R.J.; Valentini, R.; Bernoux, M.; Saint-André, L. Estimating tree biomass of sub-Saharan African forests: A review of available allometric equations. Silva Fenn. 2011, 45, 477–569. [Google Scholar] [CrossRef]
- Rodríguez-Zúñiga, M.T.; Villeda-Chávez, E.; Vázquez-Lule, A.D.; Bejarano, M.; Cruz-López, M.I.; Olguín, M.; Villela Gaytán, S.A.; Flores, R. Métodos Para la Caracterización de los Manglares Mexicanos: Un Enfoque Espacial Multiescala; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, México, 2018; ISBN 978-607-8570-03-4. [Google Scholar]
- Martins, N.E.; dos Santos, L.M.; Konig Brun, F.G.; Brun, E.J.; Krefta, S.M.; Grisi Macedo, R.L. Condiciones de los árboles urbanos: Un estudio de revisión. RECyT 2018, 20, 56–61. [Google Scholar]
- Protocolo Forestal para México Versión 2.0 [Internet]. Climate Action Reserve. Available online: https://www.climateactionreserve.org/how/protocols/mexico-forest/ (accessed on 27 January 2022).
- Herrera Silveira, J.A.; Camacho, R.A.; Pech, E.; Pech, M.; Ramírez, R.J.; Teutli, H.C. Dinámica del carbono (almacenes y flujos) en manglares de México. Terra Latinoam 2016, 34, 61–72. [Google Scholar]
- Lugo, A.E.; Snedaker, S.C. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Schaeffer-Novelli, Y.; Cintrón-Molero, G.; Soares, M.L.G.; De-Rosa, T. Brazilian mangroves. Aquat. Ecosyst. Health Manag. 2000, 3, 561–570. [Google Scholar] [CrossRef]
- Mora-Carvajal, M.J.; Bustamante-González, A.; Cajuste-Bontemps, L.; Vargas-López, S.; Cruz Bello, G.M.; Ramírez-Juárez, J. Pago por servicios ambientales hidrológicos y dinámica de la cobertura arbórea en la región Iztaccíhuatl-Popocatépetl, Puebla, México. Acta Agron 2019, 68, 84–91. [Google Scholar] [CrossRef]
- Rontard, B.; Reyes, H.; Aguilar, M. Pagos por captura de carbono en el mercado voluntario en México: Diversidad y complejidad de su aplicación en Chiapas y Oaxaca. Soc. Ambient. 2020, 22, 212–236. [Google Scholar] [CrossRef]
- Tovilla-Hernández, C.; Infante-Mata, D.M.; Ovalle-Estrada, F.; De la Presa-Pérez, J.C.; García-Alfaro, J.R.; De la Cruz-Montes, G. Informe: Inventario del Manglar y Avance de la Intrusión Salina en el Ejido Úrsulo Galván, Municipio de Jalpa de Méndez, Tabasco, México; Fondo Institucional de Fomento Regional para el Desarrollo Científico, Tecnológico y de Innovación, El Colegio de la Frontera Sur, Consejo Nacional de Ciencia y Tecnología: Tapachula, México, 2013. [Google Scholar]
- Palma-López, D.J.; Jiménez-Ramírez, R.; Zavala-Cruz, J.; Bautista-Zúñiga, F.; Gavi-Reyes, F.; Palma-Cancino, D.Y. Actualización de la clasificación de suelos de Tabasco, México. Agroproductividad 2017, 10, 29–35. [Google Scholar]
- Domínguez-Domínguez, M.; Zavala-Cruz, J.; Rincón-Ramírez, J.A.; Nartínez-Zurimendi, P. Management Strategies for the Conservation, Restoration and Utilization of Mangroves in Southeastern Mexico. Wetlands 2019, 39, 907–919. [Google Scholar] [CrossRef]
- Domínguez-Domínguez, M.; Zavala-Cruz, J.; Martínez-Zurimendi, P. Manejo Forestal Sustentable de los Manglares de Tabasco; Secretaría de Recursos Naturales y Protección Ambiental. Colegio de Postgraduados: Villahermosa, México, 2011. [Google Scholar]
- Censo de Población y Vivienda 2020. México: Instituto Nacional de Estadística y Geografía. Available online: https://www.inegi.org.mx/programas/ccpv/2020/default.html#Datos_abiertos (accessed on 28 March 2023).
- Datos Geográficos Perimetrales de los Núcleos Agrarios Certificados, por Estado—Formato SHAPE. México: Gobierno de México. Available online: https://datos.gob.mx/busca/dataset/datos-geograficos-perimetrales-de-los-nucleos-agrarios-certificados-por-estado--formato-shape (accessed on 28 November 2021).
- Day, J.W., Jr.; Conner, W.H.; Ley-Lou, F.; Day, R.H.; Machado-Navarro, A. The productivity and composition of mangrove forests, Laguna de Términos, México. Aquat. Bot. 1987, 27, 267–284. [Google Scholar] [CrossRef]
- Smith, T.J.; Whelan, K.R.T. Development of allometric relations for three mangrove species in South Florida for use in the Greater Everglades Ecosystem restoration. Wetl. Ecol. Manag. 2006, 14, 409–419. [Google Scholar] [CrossRef]
- Rodríguez, R.; Jiménez, J.; Meza, J.; Aguirre, O.; Razo, R. Carbono contenido en un bosque tropical subcaducifolio en la reserva de la biosfera el cielo, Tamaulipas, México. Rev. Latinoam. Recur. Nat. 2008, 4, 215–222. [Google Scholar]
- Hughes, F.; Kauffman, B.; Jaramillo, V. Biomass, Carbon, and Nutrient Dynamics of Secondary Forests in a humid Tropical Region of Mexico. Ecology 1999, 80, 1892–1907. [Google Scholar] [CrossRef]
- Rodríguez, R.; Jiménez, J.; Aguirre, O.; Treviño, E.; Treviño, E. Estimación de carbono almacenado de niebla en Tamaulipas, México. Ciencia UANL 2006, 9, 179–188. [Google Scholar]
- Hung, D.N.; Son, N.V.; Hung, N.P. Tree allometric equation development for estimation of forest above-ground biomass in Viet Nam—Evergreen broadleaf forests in Quang Binh Province. In Tree Allometric Equation Development for Estimation of Forest Above-Ground Biomass in Viet Nam; Inoguchi, A., Henry, M., Birigazzi, L., Sola, G., Eds.; UN-REDD Programme: Hanoi, Vietnam, 2012. [Google Scholar]
- Krisnawati, H.; Adinugroho, W.C.; Imanuddin, R. Monograph Allometric Models for Estimating Tree Biomass at Various Forest Ecosystem Types in Indonesia; Research and Development Center for Conservation and Rehabilitation, Forestry Research and Development Agency, Ministry of Forestry: Bogor, Indonesia, 2012; ISBN 978-979-3145-93-8. [Google Scholar]
- Návar, J. Allometric equations for tree species and carbon stocks for forests of northwestern Mexico. For. Ecol. Manag. 2009, 257, 427–434. [Google Scholar] [CrossRef]
- Coronado-Molina, C.; Álvarez-Guillén, H.; Day, J.W.; Reyes, E.; Pérez, B.C.; Vera-Herrera, F.; Twilley, R. Litterfall dynamics in carbonate and deltaic mangrove ecosystems in the Gulf of Mexico. Wetl. Ecol. Manag. 2012, 20, 123–136. [Google Scholar] [CrossRef]
- Hernández-Nava, J.; Pascual-Barrera, A.E.; Zaldívar-Jiménez, A.; Pérez-Ceballos, R. Estructura y secuestro de carbono en manglares urbanos, fundamentos para su conservación en Isla del Carmen, Campeche, México. Bot. Sci. 2022, 100, 899–911. [Google Scholar] [CrossRef]
- Torres, J.R.; Infante-Mata, D.; Sánchez, A.J.; Espinoza-Tenorio, A.; Barba, E. Atributos estructurales, productividad (hojarasca) y fenología del manglar en la Laguna Mecoacán, Golfo de México. Rev. Biol. Trop. 2017, 65, 1592–1608. [Google Scholar] [CrossRef]
- Torres, J.R.; Barba, E.; Choix, F.J. Mangrove Productivity and Phenology in Relation to Hydroperiod and Physical–Chemistry Properties of Water and Sediment in Biosphere Reserve, Centla Wetland, Mexico. Trop. Conserv. Sci. 2018, 11, 14. [Google Scholar] [CrossRef]
- Carmona-Díaz, G.; Morales-Mávil, J.E.; Rodríguez-Luna, E. 2004. Plan de manejo para el manglar de Sontecomapan, Catemaco, Veracruz, México: Una estrategia para la conservación de sus recursos naturales. Madera Bosques 2004, 10, 5–23. [Google Scholar] [CrossRef]
- Moreno-Casasola, P.; Hernández, M.E.; Campos, C.A. Hydrology, Soil Carbon Sequestration and Water Retention along a Coastal Wetland Gradient in the Alvarado Lagoon System, Veracruz, Mexico. J. Coast. Res. 2017, 77 (Suppl. S1), 104–115. [Google Scholar] [CrossRef]
- Téllez-García, C.P.; Valdez-Hernández, J.I. Caracterización estructural del manglar en el Estero Palo Verde, laguna de Cuyutlán, Colima. Rev. Chapingo Ser. Cienc. For. Ambient. 2012, 18, 395–408. [Google Scholar] [CrossRef]
- Torres-Fernández del Campo, J.; Olvera-Vargas, M.; Figueroa-Rangel, B.L.; Cuevas-Guzmán, R.; Iñiguez-Dávalos, L.I. Patterns of Spatial Diversity and Structure of Mangrove Vegetation in Pacific West-Central Mexico. Wetlands 2018, 38, 919–931. [Google Scholar] [CrossRef]
- Romero-Berny, E.I.; Tovilla-Hernández, C.; Torrescano-Valle, N.; Schmook, B. Heterogeneidad estructural del manglar como respuesta a factores ambientales y antrópicos en el Soconusco, Chiapas, México. Polibotánica 2019, 47, 39–58. [Google Scholar] [CrossRef]
- Infante-Mata, D.; Moreno-Casasola, P.; Madero-Vega, C. Pachira aquatica, un indicador del límite del manglar? Rev. Mex. Biodivers. 2014, 85, 143–160. [Google Scholar] [CrossRef]
- Adame, M.F.; Santini, N.S.; Tovilla, C.; Vázquez-Lule, A.; Castro, L.; Guevara, M. Carbon stocks and soil sequestration rates of tropical riverine wetlands. Biogeosciences 2015, 12, 3805–3818. [Google Scholar] [CrossRef]
- Sreelekshmi, S.; Preethy, C.M.; Varghese, R.; Joseph, P.; Asha, C.V.; Nandan, S.B.; Radhakrishnan, C.K. Diversity, stand structure, and zonation pattern of mangroves in southwest coast of India. J. Asia-Pac. Biodivers. 2018, 11, 573–582. [Google Scholar] [CrossRef]
- Grinson, G.; Krishnan, P.; Mini, K.G.; Salim, S.S.; Ragavan, P.; Tenjing, S.Y.; Muruganandam, R.; Dubey, S.K.; Gopalakrishnan, A.; Purvaja, R.; et al. Structure and regeneration status of mangrove patches along the estuarine and coastal stretches of Kerala, India. J. For. Res. 2019, 30, 507–518. [Google Scholar] [CrossRef]
- Shah, K.; Kamal, A.H.M.; Rosli, Z.; Hakeem, R.K.; Hoque, M.M. Composition and diversity of plants in Sibuti mangrove forest, Sarawak, Malaysia. For. Sci. Technol. 2016, 12, 70–76. [Google Scholar] [CrossRef]
- Rijal, S.S.; Pham, T.D.; Noer’Aulia, S.; Putera, M.I.; Saintilan, N. Mapping Mangrove Above-Ground Carbon Using Multi-Source Remote Sensing Data and Machine Learning Approach in Loh Buaya, Komodo National Park, Indonesia. Forests 2023, 14, 94. [Google Scholar] [CrossRef]
- Iimura, Y.; Kinjo, K.; Kondo, M.; Ohtsuka, T. Soil carbon stocks and their primary origin at mature mangrove ecosystems in the estuary of Fukido River, Ishigaki Island, southwestern Japan. Soil Sci. Plant Nutr. 2019, 65, 435–443. [Google Scholar] [CrossRef]
- Jiménez, J.A.; Lugo, A.E.; Cintron, G. Tree Mortality in Mangrove Forests. Biotropica 1985, 17, 177–185. [Google Scholar] [CrossRef]
- Gross, J.; Flores, E.E.; Schwendenmann, L. Stand Structure and Aboveground Biomass of a Pelliciera rhizophorae Mangrove Forest, Gulf of Monitjo Ramsar Site, Pacific Coast, Panama. Wetlands 2014, 34, 55–65. [Google Scholar] [CrossRef]
- Ragavan, P.; Kumar, S.; Kathiresan, K.; Mohan, P.M.; Jayaraj, R.S.C.; Ravichandaran, K.; Rana, T.S. Biomass and vegetation carbon stock in mangrove forests of the Andaman Islands, India. Hydrobiologia 2021, 848, 4673–4693. [Google Scholar] [CrossRef]
- Uddin, M.M.; Hossain, M.M.; Aziz, A.A.; Lovelock, C.E. Ecological development of mangrove plantations in the Bangladesh Delta. For. Ecol. Manag. 2022, 517, 120269. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Bernardino, A.F.; Ferreira, T.O.; Giovannoni, L.R.; de, O. Gomes, L.E.; Romero, D.J.; Jimenez, L.C.Z.; Ruiz, F. Carbon stocks of mangroves and salt marshes of the Amazon region, Brazil. Biol. Lett. 2018, 14, 20180208. [Google Scholar] [CrossRef]
- Kasawani, I.; Kamaruzaman, J.; Nurun-nadhirah, M.I. A Study of Forest Structure, Diversity Index and Above-ground Biomass at Tok Bali Mangrove Forest, Kelantan, Malaysia. In Proceedings of the 5th WSEAS International Conference on Environment, Ecosystems and Development, Tenerife, Spain, 14–16 December 2007; pp. 269–276. [Google Scholar]
- Shaltout, K.H.; Ahmed, M.T.; Alrumman, S.A.; Ahmed, D.A.; Eid, E.M. Standing Crop Biomass and Carbon Content of Mangrove Avicennia marina (Forssk.) Vierh. along the Red Sea Coast of Saudi Arabia. Sustainability 2021, 13, 13996. [Google Scholar] [CrossRef]
- Phan, S.M.; Thi, N.H.T.; Nguyen, T.K.; Lovelock, C. Modelling above ground biomass accumulation of mangrove plantations in Vietnam. For. Ecol. Manag. 2019, 432, 376–386. [Google Scholar] [CrossRef]
- Vorster, A.G.; Evangelista, P.H.; Stovall, A.E.L.; Ex, S. Variability and uncertainty in forest biomass estimates from the tree to landscape scale: The role of allometric equations. Carbon Balance Manag. 2020, 15, 8. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Lule, A.; Colditz, R.; Herrera-Silveira, J.; Guevara, M.; Rodríguez-Zúñiga, M.T.; Cruz, I.; Ressl, R.; Vargas, R. Greenness trends and carbon stocks of mangroves across Mexico. Environ. Res. Leton. 2019, 14, 075010. [Google Scholar] [CrossRef]
- Vinod, K.; Asokan, P.K.; Zacharia, P.U.; Ansar, C.P.; Vijayan, G.; Anasukoya, A.; Kunhi Koya, V.A.; Nikhiljith, M.K. Assessment of biomass and carbon stocks in mangroves of Thalassery estuarine wetland of Kerala, south-west coast of India. J. Coast. Res. 2019, 86 (Suppl. S1), 209–217. [Google Scholar] [CrossRef]
- de Jong Cleyndert, G.; Cuni-Sanchez, A.; Seki, H.A.; Shirima, D.D.; Munishi, P.K.T.; Burgess, N.; Calders, K.; Marchant, R. The effects of seaward distance on above and below ground carbon stocks in estuarine mangrove ecosystems. Carbon Balance Manag. 2020, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Gómez, J.G.; Lluch-Cota, S.E.; Rivera-Monroy, V.H.; Lluch-Cota, D.B.; Troyo-Diéguez, E.; Oechel, W.; Serviere-Zaragoza, E. Mangrove wetland productivity and carbon stocks in an arid zone of the Gulf of California (La Paz Bay, Mexico). For. Ecol. Manag. 2019, 442, 135–147. [Google Scholar] [CrossRef]
- Wang, G.; Guan, D.; Xiao, L.; Peart, M.R. Ecosystem carbon storage affected by intertidal locations and climatic factors in three estuarine mangrove forests of South China. Reg. Environ. Chang. 2019, 19, 1701–1712. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species Group | Aboveground Biomass Equation | Source |
|---|---|---|
| R. mangle | Day et al. [42] | |
| A. germinans | Smith and Whelan [43] | |
| L. racemosa | Day et al. [42] | |
| Pachira aquatica | Rodríguez et al. [44] | |
| Lonchocarpus luteomaculatus | Hughes et al. [45] | |
| Ficus maxima | Rodríguez et al. [46] | |
| Annona glabra | Hung et al. [47] | |
| Cocos nucifera | Krisnawati et al. [48] | |
| Pithecellobium dulce | Návar [49] |
| Code | Description | Decay Adjustment (%) | Biomass Considered (%) |
|---|---|---|---|
| 1 | Very healthy/dominant: The crown is full on all sides. The crown of the tree is generally above those of other trees around it and has minimal competition. | 0 | 100 |
| 2 | Healthy/codominant: The crown is not full on all sides due to competition with annex trees but has a proportion of the crown that receives full sunlight. | 0 | 100 |
| 3 | Suppressed: The crown is usually under those of other trees and has live but decaying foliage. | 0 | 100 |
| 4 | Dead with some deterioration: The tree has no foliage. The branches and the upper part are intact, and the bark is retained to the tree. | 25 | 75 |
| 5 | Dead with advanced deterioration: The tree may not have the upper part, it has fungal bodies, and the bark is not present. The tree no longer has foliage in the crown. | 50 | 50 |
| Tree Section | Standardized Portion of Biomass in Each Section of the Tree (%) | Current Portion of Defect in Each Section of the Tree (%) |
|---|---|---|
| Upper 1/3 | 10 | 0–100 |
| Middle 1/3 | 30 | 0–100 |
| Lower 1/3 | 60 | 0–100 |
| Species | Individuals ha−1 | Number of Plots with the Species |
|---|---|---|
| L. racemosa | 3178.1 | 108 |
| R. mangle | 258.4 | 51 |
| Pachira aquatica | 29.2 | 17 |
| A. germinans | 15.4 | 10 |
| Lonchocarpus luteomaculatus | 15.4 | 7 |
| Cocos nucifera | 12.3 | 3 |
| Annona glabra | 3.8 | 2 |
| Ficus maxima | 2.3 | 3 |
| Pithecellobium dulce | 1.5 | 1 |
| Species | 5.0–10 | 10.1–15 | 15.1–20 | 20.1–25 | 25.1–30 | 30.1–35 | 35.1–40 | >40 |
|---|---|---|---|---|---|---|---|---|
| cm | ||||||||
| L. racemosa | 2853 | 1063 | 169 | 28 | 18 | 8 | 4 | 6 |
| R. mangle | 220 | 95 | 13 | 4 | 1 | 0 | 1 | 1 |
| P. aquatica | 28 | 10 | 0 | 0 | 0 | 0 | 0 | 0 |
| L. luteomaculatus | 18 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| A. germinans | 12 | 4 | 2 | 0 | 0 | 2 | 0 | 0 |
| C. nucifera | 0 | 0 | 3 | 8 | 4 | 1 | 0 | 0 |
| A. glabra | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| F. maxima | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| P. dulce | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Plots with the diameter class | 117 | 108 | 70 | 21 | 10 | 8 | 3 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ávila-Acosta, C.R.; Domínguez-Domínguez, M.; Vázquez-Navarrete, C.J.; Acosta-Pech, R.G.; Martínez-Zurimendi, P. Aboveground Biomass and Carbon Storage in Mangrove Forests in Southeastern Mexico. Resources 2024, 13, 41. https://doi.org/10.3390/resources13030041
Ávila-Acosta CR, Domínguez-Domínguez M, Vázquez-Navarrete CJ, Acosta-Pech RG, Martínez-Zurimendi P. Aboveground Biomass and Carbon Storage in Mangrove Forests in Southeastern Mexico. Resources. 2024; 13(3):41. https://doi.org/10.3390/resources13030041
Chicago/Turabian StyleÁvila-Acosta, Carlos Roberto, Marivel Domínguez-Domínguez, César Jesús Vázquez-Navarrete, Rocío Guadalupe Acosta-Pech, and Pablo Martínez-Zurimendi. 2024. "Aboveground Biomass and Carbon Storage in Mangrove Forests in Southeastern Mexico" Resources 13, no. 3: 41. https://doi.org/10.3390/resources13030041
APA StyleÁvila-Acosta, C. R., Domínguez-Domínguez, M., Vázquez-Navarrete, C. J., Acosta-Pech, R. G., & Martínez-Zurimendi, P. (2024). Aboveground Biomass and Carbon Storage in Mangrove Forests in Southeastern Mexico. Resources, 13(3), 41. https://doi.org/10.3390/resources13030041