Suitability of Wild Phragmites australis as Bio-Resource: Tissue Quality and Morphology of Populations from Three Continents




Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sampling Sites
2.2. Morphological Measurements
2.3. Relative Leaf Water Content
2.4. Shoot Ion, Carbon, and Nitrogen Concentrations
2.5. Shoot Ash Concentration and Loss on Ignition
2.6. Statistical Data Treatment
3. Results
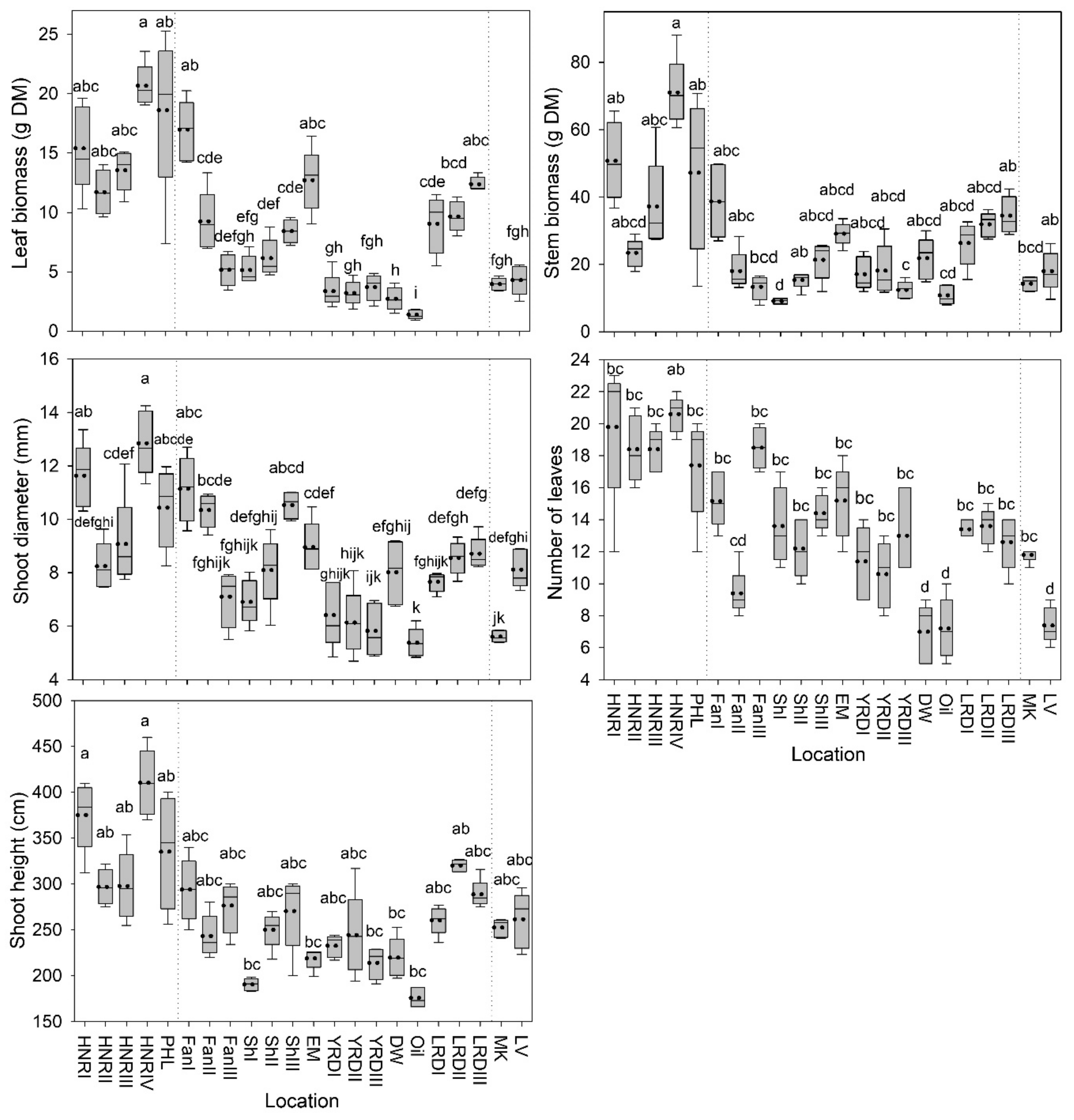
3.1. Shoot Morphology and Biomass
3.2. Shoot Ash Concentration and Biomass Water Content
3.3. Shoot Ion, Carbon, and Nitrogen Concentrations
3.4. Correlation between Parameters and Hierarchical Clustering
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, P.; Zhang, L.; Chang, Y.; Xu, M.; Hao, Y.; Liang, S.; Liu, G.; Yang, Z.; Wang, C. Food-energy-water (FEW) nexus for urban sustainability: A comprehensive review. Resour. Conserv. Recycl. 2019, 142, 215–224. [Google Scholar] [CrossRef]
- Pandiyan, K.; Singh, A.; Singh, S.; Saxena, A.K.; Nain, L. Technological interventions for utilization of crop residues and weedy biomass for second generation bio-ethanol production. Renew. Energy 2019, 132, 723–741. [Google Scholar] [CrossRef]
- Mayer, F.; Gerin, P.A.; Noo, A.; Lemaigre, S.; Stilmant, D.; Schmit, T.; Leclech, N.; Ruelle, L.; Gennen, J.; von Francken-Welz, H.; et al. Assessment of energy crops alternative to maize for biogas production in the Greater Region. Bioresour. Technol. 2014, 166, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, B.M.; Baxter, L.L.; Miles, T.R.; Miles, T.R. Combustion properties of biomass. Fuel Process. Technol. 1998, 54, 17–46. [Google Scholar] [CrossRef]
- Marschner, H. 13—Nutrient Availability in Soils. In Mineral Nutrition of Higher Plants, 2nd ed.; Marschner, H., Ed.; Academic Press: London, UK, 1995; pp. 483–507. ISBN 978-0-12-473542-2. [Google Scholar]
- Ren, L.; Eller, F.; Lambertini, C.; Guo, W.Y.; Brix, H.; Sorrell, B.K. Assessing nutrient responses and biomass quality for selection of appropriate paludiculture crops. Sci. Total Environ. 2019, 664, 1150–1161. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Matias, J.C.O.; Catalão, J.P.S. Biomass combustion systems: A review on the physical and chemical properties of the ashes. Renew. Sustain. Energy Rev. 2016, 53, 235–242. [Google Scholar] [CrossRef]
- Lessmann, J.M.; Brix, H.; Bauer, V.; Clevering, O.A.; Comin, F.A. Effect of climatic gradients on the photosynthetic responses of four Phragmites australis populations. Aquat. Bot. 2001, 69, 109–126. [Google Scholar] [CrossRef]
- Eller, F.; Skálová, H.; Caplan, J.S.; Bhattarai, G.P.; Burger, M.K.; Cronin, J.T.; Guo, W.-Y.; Guo, X.; Hazelton, E.L.G.; Kettenring, K.M.; et al. Cosmopolitan species as models for ecophysiological responses to global change: The common reed Phragmites australis. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Dragoni, F.; Giannini, V.; Ragaglini, G.; Bonari, E.; Silvestri, N. Effect of Harvest Time and Frequency on Biomass Quality and Biomethane Potential of Common Reed (Phragmites australis) Under Paludiculture Conditions. BioEnergy Res. 2017, 10, 1066–1078. [Google Scholar] [CrossRef]
- Geurts, J.J.M.; van Duinen, G.-J.A.; van Belle, J.; Wichmann, S.; Wichtmann, W.; Fritz, C. Recognize the high potential of paludiculture on rewetted peat soils to mitigate climate change. Landbauforschung 2019, 69, 5–8. [Google Scholar] [CrossRef]
- Wichtmann, W.; Couwenberg, J. Reed as a Renewable Resource and Other Aspects of Paludiculture Foreword. Mires Peat 2013, 13, 1–2. [Google Scholar]
- Wichmann, S. Commercial viability of paludiculture: A comparison of harvesting reeds for biogas production, direct combustion, and thatching. Ecol. Eng. 2017. [Google Scholar] [CrossRef]
- Brix, H. Genetic diversity, ecophysiology and growth dynamics of reed (Phragmites australis)—Introduction. Aquat. Bot. 1999, 64, 179–184. [Google Scholar]
- Koppitz, H. Analysis of genetic diversity among selected populations of Phragmites australis world-wide. Aquat. Bot. 1999, 64, 209–221. [Google Scholar] [CrossRef]
- Hansen, D.L.; Lambertini, C.; Jampeetong, A.; Brix, H. Clone-specific differences in Phragmites australis: Effects of ploidy level and geographic origin. Aquat. Bot. 2007, 86, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Eller, F.; Lambertini, C.; Nguyen, L.X.; Achenbach, L.; Brix, H. Interactive effects of elevated temperature and CO2 on two phylogeographically distinct clones of common reed (Phragmites australis). AoB Plants 2013, 5, 1–13. [Google Scholar] [CrossRef]
- Lambertini, C.; Gustafsson, M.H.G.; Frydenberg, J.; Lissner, J.; Speranza, M.; Brix, H. A phylogeographic study of the cosmopolitan genus Phragmites (Poaceae) based on AFLPs. Plant Syst. Evol. 2006, 258, 161–182. [Google Scholar] [CrossRef] [Green Version]
- Klimeš, L. Phragmites australis at an extreme altitude: Rhizome architecture and its modelling. Folia Geobot. 2000, 35, 403–417. [Google Scholar] [CrossRef]
- Bellavance, M.E.; Brisson, J. Spatial dynamics and morphological plasticity of common reed (Phragmites australis) and cattails (Typha sp.) in freshwater marshes and roadside ditches. Aquat. Bot. 2010, 93, 129–134. [Google Scholar] [CrossRef]
- Saltonstall, K. Genetic variation among north American Populations of Phragmites australis: Implications for management. Estuaries 2003, 26, 444–451. [Google Scholar] [CrossRef]
- Hauber, D.P.; Saltonstall, K.; White, D.A.; Hood, C.S. Genetic Variation in the Common Reed, Phragmites australis, in the Mississippi River Delta Marshes: Evidence for Multiple Introductions. Estuaries Coasts 2011, 34, 851–862. [Google Scholar] [CrossRef]
- Lambertini, C.; Mendelssohn, I.A.; Gustafsson, M.G.H.; Olesen, B.; Riis, T.; Sorrell, B.K.; Brix, H. Tracing the origin of Gulf Coast Phragmites (Poaceae)—A story of long distance dispersal and hybridization. Am. J. Bot. 2012, 99, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, C.; Sorrell, B.K.; Riis, T.; Olesen, B.; Brix, H. Exploring the borders of European Phragmites within a cosmopolitan genus. AoB Plants 2012, pls020. [Google Scholar] [CrossRef]
- Nguyen, L.X.; Lambertini, C.; Sorrell, B.K.; Eller, F.; Achenbach, L.; Brix, H. Photosynthesis of co-existing Phragmites haplotypes in their non-native range: Are characteristics determined by adaptations derived from their native origin? AoB Plants 2013, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Achenbach, L.; Eller, F.; Nguyen, L.X.; Brix, H. Differences in salinity tolerance of genetically distinct Phragmites australis clones. AoB Plants 2013, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Kiviat, E. Ecosystem services of Phragmites in North America with emphasis on habitat functions. AoB Plants 2013, 5. [Google Scholar] [CrossRef]
- Haslam, S.M. Early decay of Phragmites thatch: An outline of the problem. Aquat. Bot. 1989, 35, 129–132. [Google Scholar] [CrossRef]
- Boar, R.R.; Kirby, J.J.H.; Leeming, D.J. Variations in the quality of the thatching reed Phragmites australis from wetlands in East Anglia, England. In Floodplains: Interdisciplinary Approaches; Marriott, S., Alexander, J., Eds.; Geological Soc. Publishing House: Avon, UK, 1999; Volume 163, pp. 145–151. [Google Scholar]
- Nuamah, L.A.; Li, Y.; Pu, Y.; Nwankwegu, A.S.; Haikuo, Z.; Norgbey, E.; Banahene, P.; Bofah-Buoh, R. Constructed wetlands, status, progress, and challenges. The need for critical operational reassessment for a cleaner productive ecosystem. J. Clean. Prod. 2020, 269, 122340. [Google Scholar] [CrossRef]
- Brix, H.; Ye, S.; Laws, E.A.; Sun, D.; Li, G.; Ding, X.; Yuan, H.; Zhao, G.; Wang, J.; Pei, S. Large-scale management of common reed, Phragmites australis, for paper production: A case study from the Liaohe Delta, China. Ecol. Eng. 2014, 73, 760–769. [Google Scholar] [CrossRef]
- Ren, L.; Eller, F.; Lambertini, C.; Guo, W.Y.; Sorrell, B.K.; Brix, H. Minimum Fe requirement and toxic tissue concentration of Fe in Phragmites australis: A tool for alleviating Fe-deficiency in constructed wetlands. Ecol. Eng. 2018, 118, 152–160. [Google Scholar] [CrossRef]
- Ren, L.; Guo, X.; Liu, S.; Yu, T.; Guo, W.; Wang, R.; Ye, S.; Lambertini, C.; Brix, H.; Eller, F. Intraspecific variation in Phragmites australis: Clinal adaption of functional traits and phenotypic plasticity vary with latitude of origin. J. Ecol. 2020. [Google Scholar] [CrossRef]
- Hazelton, E.L.G.; Mozdzer, T.J.; Burdick, D.M.; Kettenring, K.M.; Whigham, D.F. SPECIAL ISSUE: Phragmites australis in North America and Europe Phragmites australis management in the United States: 40 years of methods and outcomes. AoB Plants 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Lambertini, C.; Guo, W.Y.; Ye, S.; Eller, F.; Guo, X.; Li, X.Z.; Sorrell, B.K.; Speranza, M.; Brix, H. Phylogenetic diversity shapes salt tolerance in Phragmites australis estuarine populations in East China. Sci. Rep. 2020, 10, 17645. [Google Scholar] [CrossRef]
- Han, G.; Chu, X.; Xing, Q.; Li, D.; Yu, J.; Luo, Y.; Wang, G.; Mao, P.; Rafique, R. Effects of episodic flooding on the net ecosystem CO2 exchange of a supratidal wetland in the Yellow River Delta. J. Geophys. Res. Biogeosci. 2015, 120, 1506–1520. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Krauss, K.W.; Brix, H.; Wei, M.; Olsson, L.; Yu, X.; Ma, X.; Wang, J.; Yuan, H.; Zhao, G.; et al. Inter-Annual Variability of Area-Scaled Gaseous Carbon Emissions from Wetland Soils in the Liaohe Delta, China. PLoS ONE 2016, 11, e0160612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swearingen, J.; Saltonstall, K. Phragmites Field Guide: Distinguishing Native and Exotic Forms of Common Reed (Phragmites australis) in the United States. Available online: http://www.nps.gov/plants/alien/pubs/index.htm (accessed on 28 September 2020).
- Saltonstall, K.; Peterson, P.M.; Soreng, R.J. Recognition of Phragmites australis subsp. americanus (Poaceae: Arundinoideae) in North America: Evidence from morphological and genetic analyses. SIDA 2004, 21, 683–692. [Google Scholar]
- Barrs, H.D.; Weatherley, P.E. A Re-Examination of the Relative Turgidity Technique for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.; James, R.; Munns, R.; Condon, T.; Passioura, J. Osmotic adjustment leads to anomalously low estimates of relative water content in wheat and barley. Funct. Plant Biol. 2008, 35, 1172–1182. [Google Scholar] [CrossRef]
- Granato, D.; Santos, J.S.; Escher, G.B.; Ferreira, B.L.; Maggio, R.M. Use of principal component analysis (PCA) and hierarchical cluster analysis (HCA) for multivariate association between bioactive compounds and functional properties in foods: A critical perspective. Trends Food Sci. Technol. 2018, 72, 83–90. [Google Scholar] [CrossRef]
- Guo, W.-Y.; Lambertini, C.; Nguyen, L.X.; Li, X.-Z.; Brix, H. Preadaptation and post-introduction evolution facilitate the invasion of Phragmites australis in North America. Ecol. Evol. 2014, 4, 4567–4577. [Google Scholar] [CrossRef]
- Back, C.L.; Holomuzki, J.R.; Klarer, D.M.; Whyte, R.S. Herbiciding invasive reed: Indirect effects on habitat conditions and snail-algal assemblages one year post-application. Wetl. Ecol. Manag. 2012, 20, 419–431. [Google Scholar] [CrossRef]
- Rohal, C.B.; Cranney, C.; Hazelton, E.L.G.; Kettenring, K.M. Invasive Phragmites australis management outcomes and native plant recovery are context dependent. Ecol. Evol. 2019, 9, 13835–13849. [Google Scholar] [CrossRef] [Green Version]
- Vaicekonyte, R.; Kiviat, E.; Nsenga, F.; Ostfeld, A. An exploration of common reed (Phragmites australis) bioenergy potential in North America. Mires Peat 2013, 13, 1–9. [Google Scholar]
- Eller, F.; Ehde, P.M.; Oehmke, C.; Ren, L.; Brix, H.; Sorrell, B.K.; Weisner, S.E.B. Biomethane Yield from Different European Phragmites australis Genotypes, Compared with Other Herbaceous Wetland Species Grown at Different Fertilization Regimes. Resources 2020, 9, 57. [Google Scholar] [CrossRef]
- Woehler-Geske, A.; Moschner, C.R.; Gellerich, A.; Militz, H.; Greef, J.-M.; Hartung, E. Provenances and properties of thatching reed (Phragmites australis). Landbauforschung 2016, 66, 1–10. [Google Scholar] [CrossRef]
- De Araujo, V.K.R.; da Silva, G.B.; de Lima Araújo, E.; de Mendonça Pimentel, R.M.; da Silva, K.A. Spatio-temporal variation in leaf morphofunctional attributes and relation to growth and survival of young woody plants. Braz. J. Bot. 2019, 42, 1–11. [Google Scholar] [CrossRef]
- Moore, G.E.; Burdick, D.M.; Peter, C.R.; Keirstead, D.R. Belowground biomass of Phragmites australis in coastal marshes. Northeast. Nat. 2012, 19, 611–626. [Google Scholar] [CrossRef]
- Haslam, S.M. Stem types of Phragmites communis Trin. Ann. Bot. 1969, 33, 127–131. [Google Scholar] [CrossRef]
- Engloner, A.I. Structure, growth dynamics and biomass of reed (Phragmites australis)—A review. Flora 2009, 204, 331–346. [Google Scholar] [CrossRef]
- Bates, T.E. Factors affecting critical nutrient concentrations in plants and their evaluation: A review. Soil Sci. 1971, 12, 116–130. [Google Scholar] [CrossRef]
- Lu, M.; Caplan, J.S.; Bakker, J.D.; Langley, J.A.; Mozdzer, T.J.; Drake, B.G.; Megonigal, J.P. Allometry data and equations for coastal marsh plants. Ecology 2016, 97, 3554. [Google Scholar] [CrossRef] [Green Version]
- Geurts, J.J.M.; Oehmke, C.; Lambertini, C.; Eller, F.; Sorrell, B.K.; Mandiola, S.R.; Grootjans, A.P.; Brix, H.; Wichtmann, W.; Lamers, L.; et al. Nutrient removal potential and biomass production by Phragmites australis and Typha latifolia on European rewetted peat and mineral soils. Stoten 2020. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, I.; Kauter, D. The influence of nitrogen fertilizer on the yield and combustion quality of whole grain crops for solid fuel use. Ind. Crops Prod. 2003, 17, 103–117. [Google Scholar] [CrossRef]
- Carson, B.D.; Lishawa, S.C.; Tuchman, N.C.; Monks, A.M.; Lawrence, B.A.; Albert, D.A. Harvesting invasive plants to reduce nutrient loads and produce bioenergy: An assessment of Great Lakes coastal wetlands. Ecosphere 2018, 9, e02320. [Google Scholar] [CrossRef]
- Kaltschmitt, M.; Hartmann, H.; Hofbauer, H. Energie aus Biomasse: Grundlagen, Techniken und Verfahren; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 9783540850946. [Google Scholar]
- Stitt, M.; Krapp, A. The interaction between elevated carbon dioxide and nitrogen nutrition: The physiological and molecular background. Plant Cell Environ. 1999, 22, 553–621. [Google Scholar] [CrossRef]
- Mason, C.F.; Bryant, R.J. Production, nutrient content and decomposition of Phragmites communis Trin. and Typha angustifolia L. J. Ecol. 1975, 63, 71–95. [Google Scholar] [CrossRef]
- Kandel, T.P.; Sutaryo, S.; Møller, H.B.; Jørgensen, U.; Lærke, P.E. Chemical composition and methane yield of reed canary grass as influenced by harvesting time and harvest frequency. Bioresour. Technol. 2013, 130, 659–666. [Google Scholar] [CrossRef]
- Risén, E.; Gregeby, E.; Tatarchenko, O.; Blidberg, E.; Malmström, M.E.; Welander, U.; Gröndahl, F. Assessment of biomethane production from maritime common reed. J. Clean. Prod. 2013, 53, 186–194. [Google Scholar] [CrossRef]
- De Groot, C.C.; Marcelis, L.F.M.; van den Boogaard, R.; Kaiser, W.M.; Lambers, H. Interaction of nitrogen and phosphorus nutrition in determining growth. Plant Soil 2003, 248, 257–268. [Google Scholar] [CrossRef]
- Ksenofontova, T. Morphology, production and mineral contents in Phragmites australis in different waterbodies of the Estonian SSR. Folia Geobot. Phytotaxon. 1988, 23, 17–43. [Google Scholar] [CrossRef]
- Zehra, A.; Gul, B.; Ansari, R.; Khan, M.A. Role of calcium in alleviating effect of salinity on germination of Phragmites karka seeds. S. Afr. J. Bot. 2012, 78, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Sheng, C.D.; Azevedo, J.L.T. Estimating the higher heating value of biomass fuels from basic analysis data. Biomass Bioenergy 2005, 28, 499–507. [Google Scholar] [CrossRef]
- Madakadze, I.C.; Stewart, K.; Peterson, P.R.; Coulman, B.E.; Smith, D.L. Switchgrass Biomass and Chemical Composition for Biofuel in Eastern Canada. Agron. J. 1999, 91, 696–701. [Google Scholar] [CrossRef]
- Delucchi, M.A. Impacts of biofuels on climate change, water use, and land use. Ann. N. Y. Acad. Sci. 2010, 1195, 28–45. [Google Scholar] [CrossRef]
- Alvarez, M.G.; Tron, F.; Mauchamp, A. Sexual versus asexual colonization by Phragmites australis: 25-year reed dynamics in a mediterranean marsh, Southern France. Wetlands 2005, 25, 639–647. [Google Scholar] [CrossRef]
- Haslam, S.M. The Development and Emergence of Buds in Phragmites communis Trin. Ann. Bot. 1969, 33, 289–301. [Google Scholar] [CrossRef]
- Melts, I.; Ivask, M.; Geetha, M.; Takeuchi, K.; Heinsoo, K. Combining bioenergy and nature conservation: An example in wetlands. Renew. Sustain. Energy Rev. 2019, 111, 293–302. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Location | GPS Coordinates | Altitude (m) | Sampling Date | Shoot Density (Number of Shoots m−2) | |
|---|---|---|---|---|---|---|
| MK | DK, Magisterkogen | 54°54′30″ N | 8°48′45.0″ E | −1 | August 2016 | 112 |
| LV | DK, Lille Vildmose | 56°53′31″ N | 10°11′17.4″ E | 4 | September 2016 | 110 |
| FanI | CN, Fanggan | 36°25′10.6″ N | 117°26′34.9″ E | 544 | August 2016 | 20 |
| FanII | CN, Fanggan field | 36°25′16.8″ N | 117°26′24″ E | 540 | August 2016 | 34 |
| FanIII | CN, Fanggan mountain | 36°25′25.8″ N | 117°27′27.8″ E | 486 | August 2016 | 40 |
| ShI | CN, Shandong stream | 36°27′17.6″ N | 117°29′15.5″ E | 266 | August 2016 | 70 |
| ShII | CN, Shandong shore | 36°27′13.9″ N | 117°30′34.7″ E | 246 | August 2016 | 80 |
| ShIII | CN, Shandong floodplain | 36°27′13.3″ N | 117°30′30.6″ E | 246 | August 2016 | 80 |
| EM | CN, Elkmarsh | 33°3′0.7″ N | 120°48′59″ E | 10 | September 2016 | 48 |
| YRDI | CN, YRD Reserve | 37°45′49″ N | 119°2′5.9″ E | 9 | September 2016 | 90 |
| YRDII | CN, YRD Reserve Tidal creek | 37°46′38.4″ N | 119°5′13.1″ E | 9 | September 2016 | 136 |
| YRDIII | CN, YRD Reserve border | 37°50′1.5″ N | 119°2′17.4″ E | 5 | September 2016 | 106 |
| DW | CN, Dongwang | 37°47′34.4″ N | 118°26′18.8″ E | 18 | September 2016 | 82 |
| Oil | CN, YRD oil field | 38°2′44.8″ N | 118°40′39.7″ E | −0.5 | September 2016 | 56 |
| LRDI | CN, LRD boardwalks | 40°57′39.2″ N | 121°48′10.8″ E | −2 | August 2017 | 40 |
| LRDII | CN, LRD road | 40°59′51.9″ N | 121°50′7.4″ E | −2 | August 2017 | 50 |
| LRDIII | CN, LRD road north | 41°0′16.4″ N | 121°50′41.4″ E | −3 | August 2017 | 62 |
| HNRI | US, HNR Darby Township | 39°53′19.4″ N | 75°15′41.4″ W | 7 | July 2017 | 52 |
| HNRII | US, HNR fishing platform | 39°52′49.6″ N | 75°15′55″ W | 2 | July 2017 | 46 |
| HNRIII | US, HNR Tinicum Township | 39°52′37.8″ N | 75°16′40.8″ W | 2 | July 2017 | 50 |
| HNRIV | US, HNR Darby Creek | 39°52′28.3″ N | 75°17′43″ W | 2 | July 2017 | 48 |
| PHL | US, ruderal wetland | 39°52′20.2″ N | 75°16′30.5″ W | 1 | August 2017 | 70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eller, F.; Guo, X.; Ye, S.; Mozdzer, T.J.; Brix, H. Suitability of Wild Phragmites australis as Bio-Resource: Tissue Quality and Morphology of Populations from Three Continents. Resources 2020, 9, 143. https://doi.org/10.3390/resources9120143
Eller F, Guo X, Ye S, Mozdzer TJ, Brix H. Suitability of Wild Phragmites australis as Bio-Resource: Tissue Quality and Morphology of Populations from Three Continents. Resources. 2020; 9(12):143. https://doi.org/10.3390/resources9120143
Chicago/Turabian StyleEller, Franziska, Xiao Guo, Siyuan Ye, Thomas J. Mozdzer, and Hans Brix. 2020. "Suitability of Wild Phragmites australis as Bio-Resource: Tissue Quality and Morphology of Populations from Three Continents" Resources 9, no. 12: 143. https://doi.org/10.3390/resources9120143
APA StyleEller, F., Guo, X., Ye, S., Mozdzer, T. J., & Brix, H. (2020). Suitability of Wild Phragmites australis as Bio-Resource: Tissue Quality and Morphology of Populations from Three Continents. Resources, 9(12), 143. https://doi.org/10.3390/resources9120143