1. Introduction
Microalgae and cyanobacteria have received increasing attention for their application in cosmetic products [
1,
2,
3,
4,
5]. Several components of these microorganisms have the characteristics to become cosmetic ingredients. Polysaccharides may be used as thickeners [
6] and moisturizing agents [
1,
5] and might be endowed with antioxidant, anti-inflammatory and anti-ageing activity [
4,
7]. Pigments have a dual nature of colourants and antioxidant compounds [
7]. Other molecules of interest are phenolics, fatty acids, sunscreens such as scytonemin and mycosporine-like amino acids, and biologically active peptides [
7,
8].
Spas are increasingly becoming wellness centres proposing cosmetic treatments and programmes to improve lifestyle (e.g., weight loss, fitness, relaxation programmes), often disjunct from the presence of site-specific thermal waters, unless traditional health treatments (e.g., for respiratory, dermatological and bone diseases) are run in parallel [
9]. In traditional spas, the health-promoting action of treatments is connected to the specific characteristics of the waters springing on-site, and an important role is played by mud therapy or pelotherapy [
10,
11]. Beneficial effects (medical and cosmetic) of thermal muds are partly due to physical properties (heat) and chemical components (e.g., minerals); nevertheless, additional functions (e.g., antioxidant and anti-inflammatory) may originate from the maturation process carried out in the presence of microorganisms, which usually include an important phototrophic component [
12].
The complex microbial communities developing around hot springs have long been named in different ways, among which are
bioglée or
bioglea [
12,
13].
Bioglea was described by Laporte et al. [
13] as a microbial community associated with a mineral component assuming a slimy consistency. In more recent definitions, it is considered as an organic peloid composed of algae and bacteria with a liquid component, generally mineral water bearing reduced sulphur; in other terms, it is a biofilm consisting of a gelatinous pellicle, which may assume different colours (yellowish, greenish, greyish or reddish), occurring at the surface or dispersed in thermal or non-thermal sulphur-bearing spring waters [
12].
Bioglea, as mentioned above, is a fundamental component in the process of thermal mud maturation [
14,
15,
16,
17]; moreover, it has been used in spas for direct treatments [
14,
18,
19] or to extract bioactives to enrich spa own-branded cosmetic products (e.g., Saturnia Bioplancton
® from Terme di Saturnia; brands from Dax, Acquadax™; and the patented Algodax).
In Dax (France), a process to obtain a mature peloid for spa treatments has been patented [
20], and, as part of this process, an interesting
bioglea cultivation step is performed. In brief, the virgin clay is mixed with thermal water, the biomass of a
Clostridium strain is isolated from the mud and cultivated in fermenters, and the
bioglea is grown under a greenhouse on inclined planes sprinkled with thermal water to allow the development of a thick mat dominated by cyanobacteria. The mix is left to mature for a pre-determined period and then is packed and distributed to spas for pelotherapy and cosmetic applications [
20]. Production processes such as this, which represent an upscale compared to traditional methods, are of great interest for the development of the sector, as they transform
bioglea from a “gift of nature” collected as it is, into an actual industrial product that may be obtained following more standardisable protocols. This process permits obtaining a higher amount of product with a higher quality (avoiding contamination with exogenous chemicals) and partially modulating production to favour a stable community with the required characteristics in terms of composition and/or bioactivity.
Another strategy to improve standardisation of the product is to isolate and separately cultivate
bioglea components, which would improve the reproducibility of the product and the optimisation of culture conditions to increase the target molecule production in a scaling-up perspective. This approach is applied in Iceland, where
Cyanobacterium aponium isolated from the Blue Lagoon, is cultivated indoors in tubular photobioreactors using geothermal water added with nutrients [
21]. On the other hand, the cultivation of isolated strains leads to the loss of the interactions among the microbial community components, which could play a role in triggering the production of bioactive molecules.
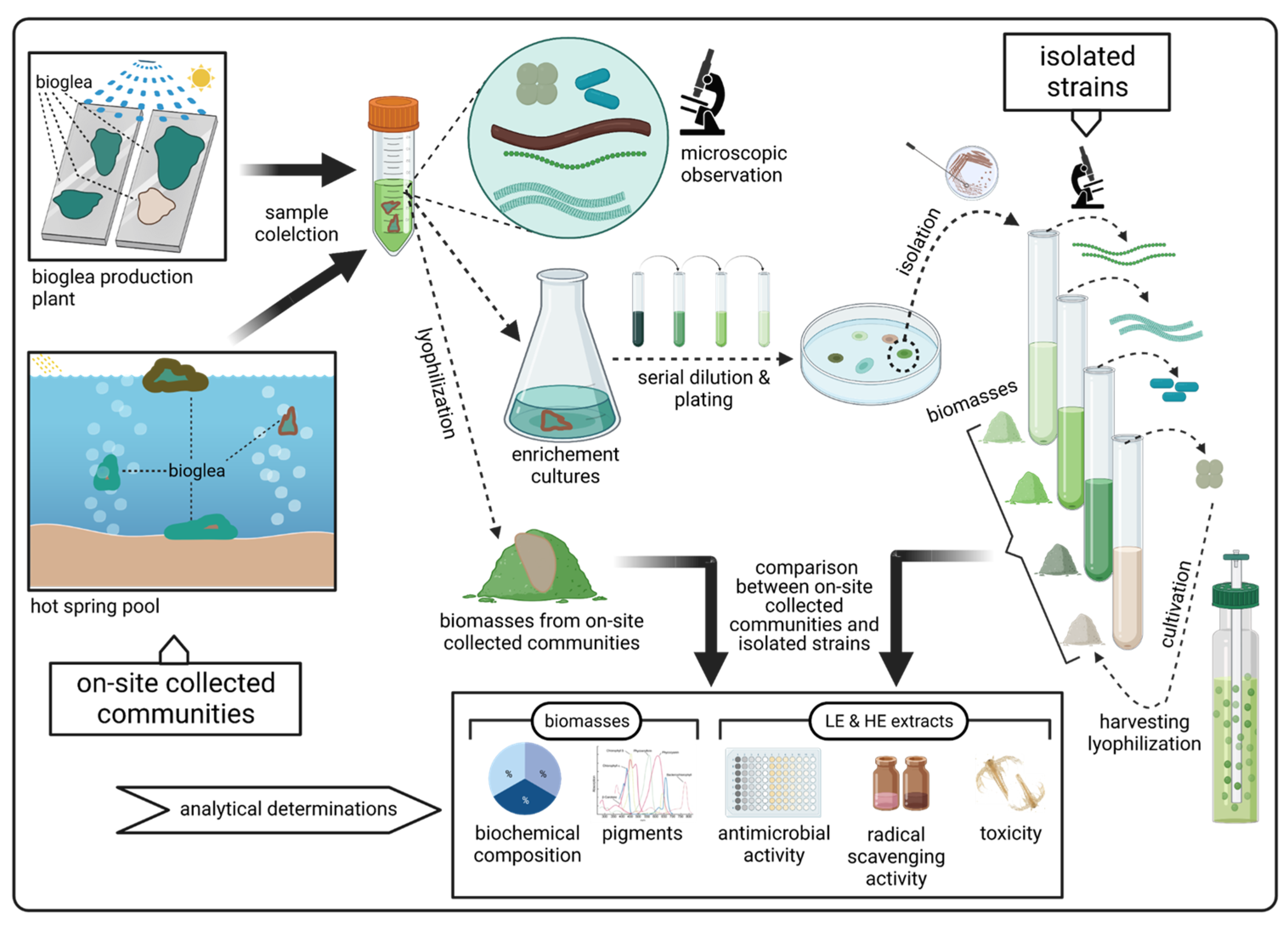
In this work, a comparison between the biochemical composition and the activity of isolated strains and microbial communities collected on site (bioglea) was performed to verify whether strains isolated from bioglea may replicate the effect of the community from which they originated. Bioactivity tests (antimicrobial activity, radical scavenging activity and toxicity) were performed on samples from microbial communities collected in spas with different water physico-chemical characteristics and on strains isolated thereof.
4. Discussion
This work aimed at comparing the composition and biological activities of communities collected in different spas with strains isolated thereof. This type of comparison is seldom dealt with in the literature. At present, the increasing request for natural products is accompanied by the need to meet stringent quality and safety standards. In this respect, the analysis carried out in this work can provide some indications useful to improve the exploitation of microbial resources from hot springs. In particular, we considered microbial communities naturally developing in hot spring environments and communities cultivated using hot spring waters and compared them with isolated strains grown under laboratory conditions. These latter represented the “proxy” for a production process performed independently of the hot spring environment. However, the results showed higher activities for communities than for isolated strains, particularly antimicrobial and radical scavenging activity of lipophilic extracts, while general biochemical composition did not significantly differ in bioglea and isolated strains, except for the bioglea sampled at the bottom of the pool in Saturnia, which showed a lower carbohydrate content. In the next paragraphs, these findings are discussed in detail.
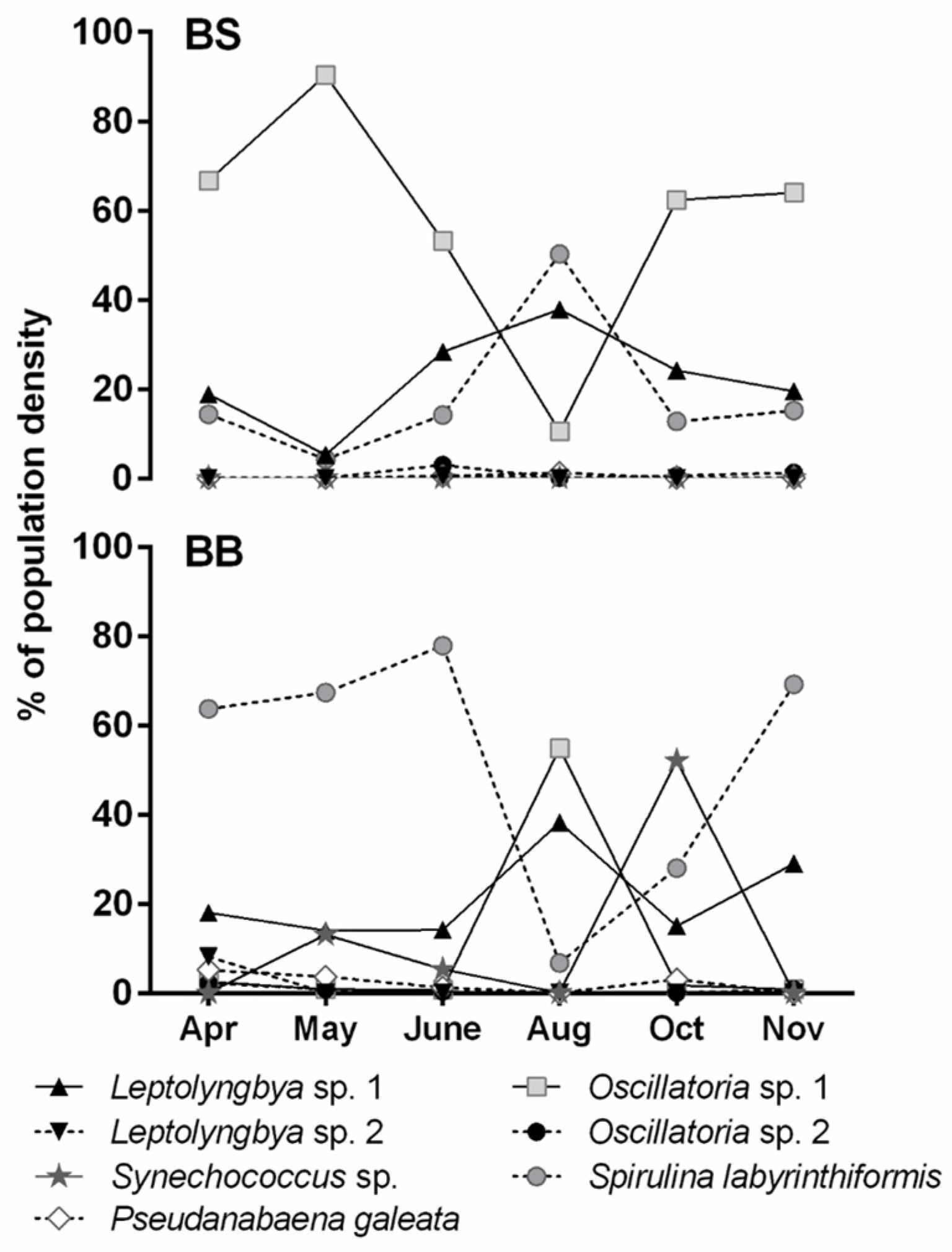
The phototrophic communities developing in the hot springs analysed had different phototrophic community compositions according to the chemico-physical characteristics of the spring water. When seasonal samples were collected (Saturnia), the cyanobacterial community appeared rather stable in its major components, thanks to constant temperature and pH. The variations in the abundance of different taxa were, most probably, due to seasonal variations in solar radiation or to the enrichment of thermal water with exogenous nutrients (e.g., following heavy precipitations). Rather little variations in the correspondent pigment signatures were found because spectra also recorded pigments from degraded trichomes, which were excluded from microscopic counts. Few studies analyse seasonal variation in cyanobacterial composition in hot springs, evidencing the role of different factors, such as sulphide concentration or heavy rainfalls [
40,
41]. In-depth characterisation of the microbial community composition was beyond the scope of the present work, nevertheless, it would be of great interest as future work to thoroughly characterise both phototrophic and chemotrophic members of these communities, by applying multifaceted approaches, including genomic techniques, both isolation-based and isolation-independent [
42,
43,
44].
When the biological activity of on-site communities is considered, besides natural variation in the ratios among the oxygenic phototrophs, several sources of variability may explain activity differences during seasons and between bioglea samples and the isolated strains, considering that more than one factor can concur. It is also worth mentioning that it was not always possible to isolate the dominant component of the community (e.g., Saturnia dark brown Oscillatoria) because of a too strict dependency on on-site conditions not reproducible in the laboratory.
As stated previously, extracts from on-site communities usually showed higher antimicrobial activity than isolated strains, particularly considering lipophilic extracts. Antimicrobial activity was searched for on a panel including common contaminants of cosmetic products (e.g., moulds such as
Penicillium and
Aspergillus) and potential pathogens, such as dermatophytes (here represented by
A. uncinatum as a model organism) or opportunistic bacteria (
S. epidermidis). Positive results on the two types of targets would have led to different potential benefits, for users’ health or for technological applications. However, moulds were not affected, except for hydrophilic extracts from water, cultivated
bioglea and mud from Bagno di Romagna. Dermatophytes were previously reported to be more sensitive to cyanobacterial extracts compared to other groups of fungi [
45], and the results obtained in this work on
A. uncinatum compared to moulds seem to confirm this higher sensitivity. Dermatophyte resistance to antifungal agents commonly used to treat their infections is a growing concern; thus, new antifungal agents would be of great importance [
46,
47]. To this end, extracts from both on-site communities and isolated strains have to confirm their effects on pathogenic dermatophytes.
Considering the total lack of antimicrobial activity in lipophilic extracts from isolated strains, a first hypothesis could be that thermal water accounts for the activity observed against A. uncinatum in lipophilic extracts of bioglea and mud samples from Saturnia and Bagno di Romagna, as well as for antibacterial activity against S. epidermidis of the hydrophilic extract from cultivated bioglea. Activity intensity varied among different community samples, notwithstanding that in the same hot spring, they were exposed to the same water environment. Different hydration levels of the collected samples could explain the variability, which, on the other hand, might also suggest that other antimicrobial molecules besides those present in water are involved. Moreover, if different hydration levels were responsible for variations in lipophilic extract activity, a similar activity pattern should have also been found for hydrophilic extracts. However, a lower frequency of activity (43%) was found in the latter. These considerations suggest that the observed activity in on-site communities was mainly not originated from water.
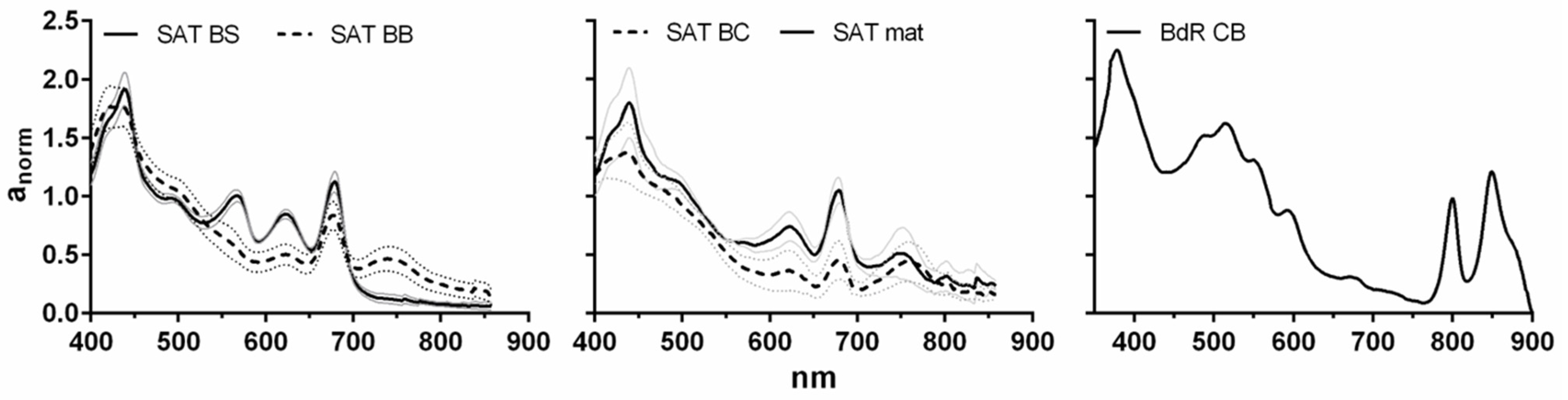
Cyanobacteria and microalgae were subjected to different environmental conditions in their habitat of origin with respect to laboratory cultures for, e.g., temperature, “medium” composition, light intensity, and bubbling. Pigments (chlorophyll
a and its degradation products, and carotenoids) and fatty acids were the major components of the extracts. It is possible that in the mats grown in hot spring environments, a higher amount of chlorophyll degradation products was present compared to biomasses from laboratory-grown isolates. Phaeophytin
a, as well as phaeophorbide
a, has been shown to have antimicrobial activity [
48,
49]. Accumulation of cell degradation products in mats and biofilms might also have led to some fatty acids or their derivatives accumulation compared to laboratory-obtained biomasses. Fatty acids can also be endowed with antifungal activity [
50]. Finally, in Saturnia, but also in Petriolo, microbial communities develop in pools open to the surrounding environment in which people bathe, so exogenous active substances may be absorbed/adsorbed in the microbial biofilms and mats, contributing to their antimicrobial activity. In the case of Saturnia
bioglea, Centini et al. [
51] actually found some contaminant cosmetics preservatives (e.g., sorbic acid, 4-hydroxy benzoic acid, phenoxyethanol) in the chloroform extracts (including those from the same samples analysed in this work), besides plant-derived molecules with known antimicrobial activity (abietic and dehydroabietic acids) [
52] and several phenolics (e.g., methoxy cinnamic acid), which origin is not known but might also have antimicrobial activity [
53]. The hydrophilic extracts contained mainly hydrophilic pigments (phycobiliproteins) and other protein-derived compounds (e.g., some free amino acids). They also contain few fatty acids. Lower activity was found in these extracts, either from
bioglea and or isolated strains, compared to lipophilic extracts, most probably because a lower antimicrobial activity is associated with the main components of these extracts. The antifungal activity of phycobiliprotein-containing crude extracts has been tested for several cyanobacterial and microalgal strains, usually resulting low or absent unless purified compounds were prepared [
54].
Besides abiotic conditions, biotic factors could also have impacted the activity. In their habitat, cyanobacteria and microalgae are growing amidst other oxygenic phototrophs, anoxygenic phototrophs, and chemoautotrophic and heterotrophic bacteria, thus needing to establish a network of interactions, among which competition through the production of antimicrobial or allelopathic metabolites [
55,
56]. The simplified biological community present in the laboratory cultures (including only some non-photosynthetic bacteria besides the oxygenic phototroph) may have reduced the necessity to invest in the production of antimicrobial molecules. Moreover, in laboratory cultures, bioactivity can vary with the growth phase, as observed for several cyanobacteria [
57,
58,
59,
60,
61]; thus, the time of harvesting of the isolated strains may have affected both the presence and the intensity of antimicrobial activity. The antifungal activity during
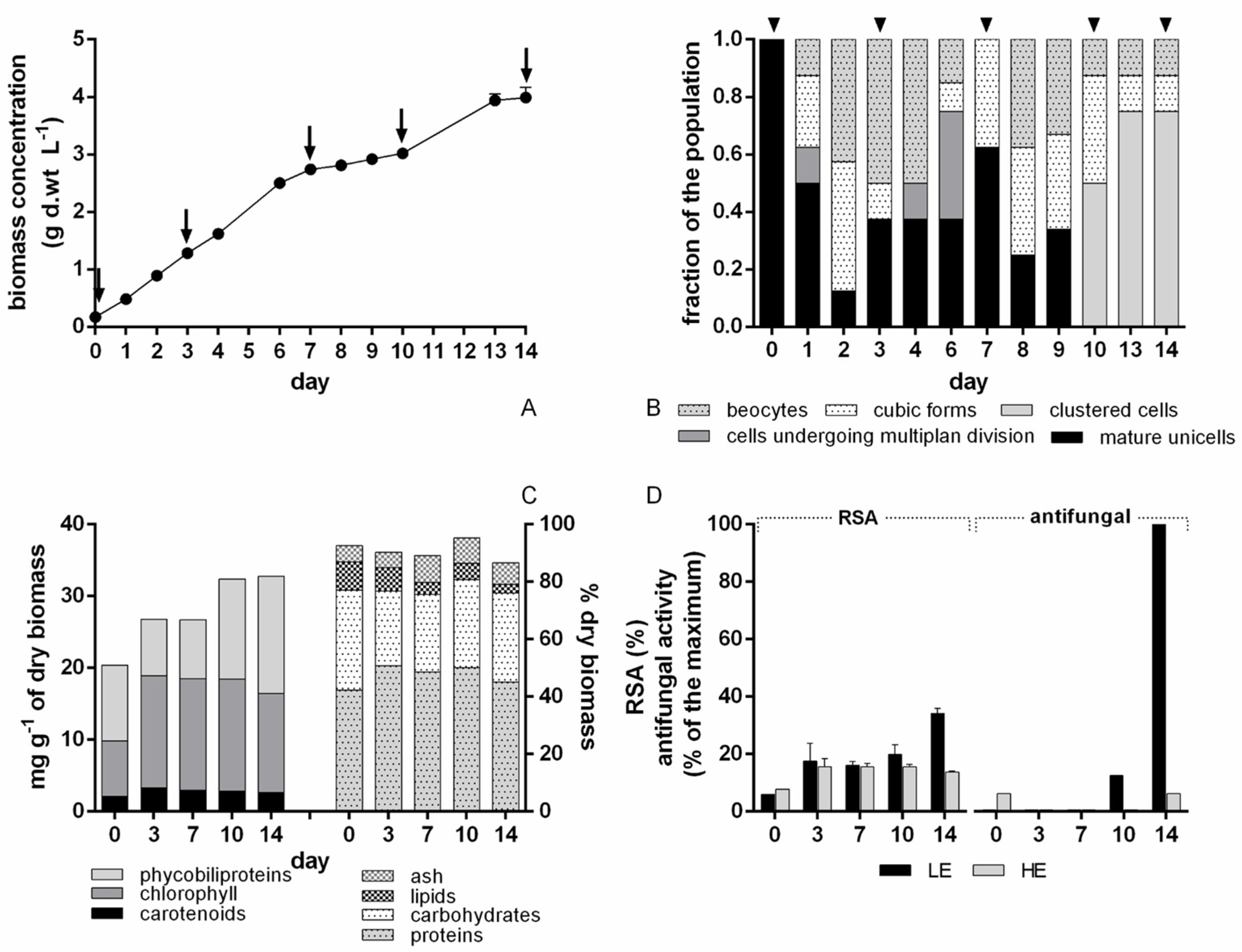
Chroococcidiopsis BIOG3 cultivation at the 10 L scale varied throughout the growth curve and life cycle stages, also appearing in the lipophilic extract (contrary to what was found at the 1 L scale). This highlights the necessity to establish for each isolated strain the best growth phase (and, if appropriate, life cycle stage) in which to harvest the biomass to maximise the active molecule production.
Little literature on the antimicrobial properties of on-site microbial communities is available. Dobretsov et al. [
62] evidence complex interactions in hot spring communities from Oman, as they found antibacterial activity in cyanobacterial mats’ extracts from two of the four analysed springs, for one of which only non-polar extract and for the other both polar and non-polar extracts were active, but against different targets. Additionally, inhibition of quorum sensing was observed in extracts from three of the springs, whereas anti-diatom activity from all four suggested the production of metabolites to regulate interactions among different microbial groups. In contrast to what was observed in our work, cyanobacterial strains isolated from the four hot springs showed a wider and more potent antibacterial activity compared to microbial mats, besides showing antifungal activity [
63]. Antimicrobial properties of strains isolated from hot springs, also for non-photosynthetic microorganisms, have been reported. Methanolic extracts from seven cyanobacterial strains (belonging to
Oscillatoria,
Synechococcus and
Synechocystis) from Geno hot spring (Iran) showed activity mainly against Gram-positive bacteria, but few also against
Escherichia coli and
Candida [
64]. Two cyanobacteria (
Gleocapsa and
Synechocystis) and two green algae (
Scenedesmus and
Chlorella) isolated from Rupite hot spring (Bulgaria) showed activity against Gram-positive bacteria in either exopolysaccharide or fatty acid fractions and, only exopolysaccharides from
Gleocapsa, against the yeast
Candida albicans [
65]. Capsular polysaccharide from a
Mastigocladus laminosus isolate from a hot spring in Néris-les-Bains (France) showed weak antibacterial activity [
66]. A peptide from a thermal spring
Bacillus licheniformis showed strong activity against Gram-positive bacteria [
67]. A
Beggiatoa-like strain from
baregine showed bacteriostatic activity against three pathogens [
68]. Non-photosynthetic bacteria might also contribute to the activity of microbial communities (and isolated strains) analysed in the present work.
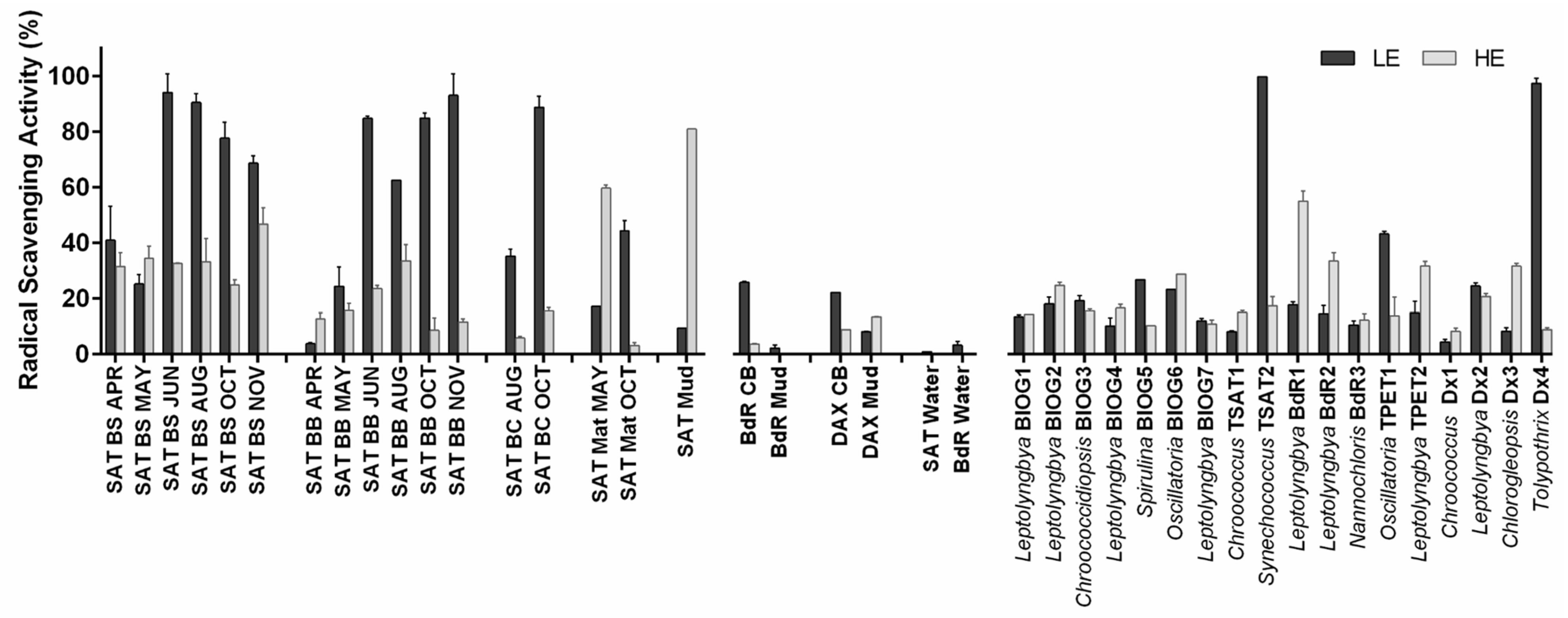
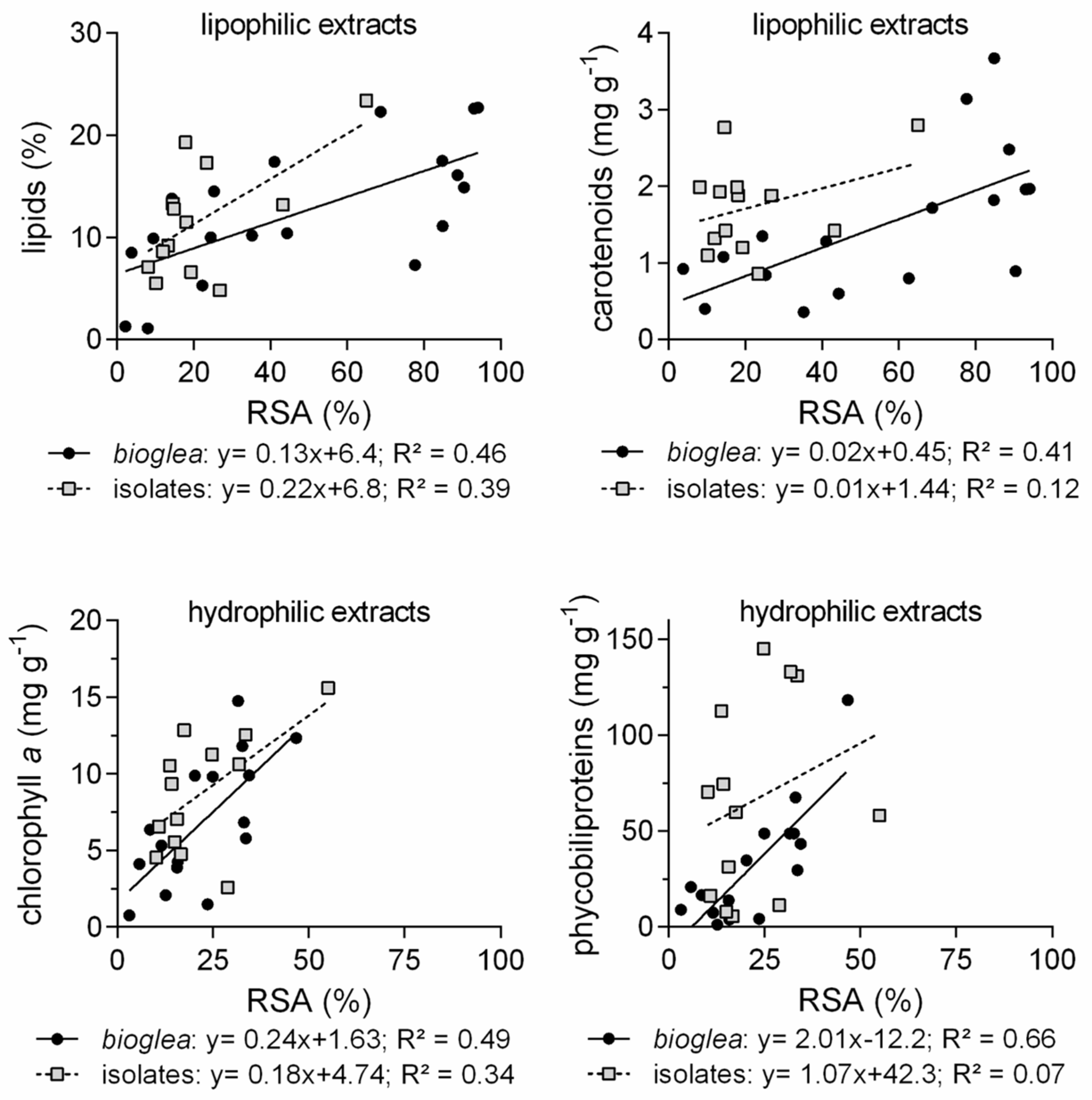
In our study, radical scavenging activity was usually higher in lipophilic extracts from on-site communities than in those from isolated strains, whereas more similar activities were observed in hydrophilic extracts. Regarding
bioglea, biotic components were the main determinant of radical scavenging activity, as thermal waters were almost inactive. Radical scavenging activity of lipophilic extracts positively correlated with total lipids and, only for communities, with carotenoid content, which was higher in summer and autumn samples. Chlorophyll and its derivatives might also have contributed to radical scavenging activity, as they are known as radical scavengers [
69]. In Saturnia
bioglea extracts, several lipophilic compounds with antioxidant activity were detected in chloroform extracts, such as the chlorophyll-derived alcohol phytol, the carotenoid lycopersene and some cinnamic acid derivatives [
51], which might have contributed to the radical scavenging activity observed. Determination of phenolics content and profile in the isolated strains will help to understand whether this component may be responsible for the observed radical scavenging activity. Phospholipid derivatives, such as diacylglycerols might also contribute. Furthermore, exogenous contaminants due to people bathing in the pool can also be responsible for part of the observed radical scavenging activity in Saturnia
bioglea, as antioxidant chemicals deriving from cosmetic products, such as BHT and squalene, were found in the extracts [
51].
Hydrophilic extracts contained phycobiliproteins, which are well-known antioxidant and anti-inflammatory molecules [
70]. Radical scavenging activity of hydrophilic extracts from on-site communities positively correlated with phycobiliprotein content, while no correlation was found for isolated strains. The high variability in phycobiliprotein content, very low (<1% of dry biomass) in some strains (e.g.,
Chroococcus TSAT1 and
Leptolyngbya BIOG4) and rather high (13–14%) in others (
Leptolyngbya strains BIOG2, TPET2 and BdR2), was paired with a fairly low and little variable radical scavenging activity. Positive correlation with chlorophyll
a in both
bioglea and isolates’ extracts could be due to the presence of more hydrophilic chlorophyll derivatives. Although in this work total carbohydrate content did not correlate with the radical scavenging activity of the hydrophilic extracts, the polysaccharidic fraction, which plays an important role in the architecture of mats [
71,
72] and represents a sink of excess energy and carbon under high radiations [
73,
74], may also have contributed to the radical scavenging activity, a well-documented property of these polymers, often coupled with anti-inflammatory activity [
66,
75,
76]. However, high total carbohydrate content (>50%) in isolated strains did not correspond to high radical scavenging activity. Other components, such as enzymes (e.g., peroxidases), might have been responsible for the observed activity in a higher proportion in isolated strains exposed to a potentially stressful regime of continuous light than in communities. Further analysis will be needed to confirm this hypothesis. Moreover, further testing of the extracts aimed at fully establishing antioxidant activity will be needed.
Both extracts from on-site communities and isolated strains were non-toxic toward the model crustacean Artemia salina. However, some of the hydrophilic extracts (mainly from isolates) were too dense and led to increased nauplii death because of poor gas exchange. These extracts derived from biomasses richer in carbohydrates and/or, in some cases, proteins. It seems that a carbohydrate + protein content above 2–3 g L−1 might have led to lethal physical effects on more than 50% of the nauplii. For comparison, a similar outcome was observed with hydrophilic extracts from the food-grade cyanobacterium Arthrospira platensis at the same extracted material concentration, while a 1:2 dilution of the extract totally removed the lethal effects (Biondi et al., unpublished data).
Two of the most interesting isolates emerging from this work are
Tolypothrix Dx4 and
Leptolyngbya BdR1. Both strains were isolated from cultivated
bioglea. In spite of lower radical scavenging and, only in the case of Dax, antimicrobial activity, these communities represent an interesting biotechnological approach for spa development. The lower performance of Dax cultivated
bioglea is easily explained by the time of sampling, i.e., during a maintenance period in autumn, in which the community was at its minimal development. For Bagno di Romagna, the good performances in terms of antimicrobial activity were most probably largely due to the action of thermal water, whereas the low radical scavenging activity may have originated from the reduced development of the oxygenic phototrophic community, usually a main driver of this activity. Optimisation of production and composition of the cultivated
bioglea may lead to improvement and increased stability of their biological activity, an important feature for the application of these communities either as a component in peloid production [
20] or for direct application in cosmetic and wellness treatments [
14,
18,
19], not to mention extract preparation for cosmetic products. For production, e.g., in Saturnia or Petriolo, purposely designed plants would be necessary to optimise exposure to light and moisture, thus allowing the development of a
bioglea similar to that growing in the pools and with comparable bioactivity profiles. The use of cultivated
bioglea instead of isolated strains would allow for preserving the interactions within the microbial community, which may be responsible for part of the activity observed. On the other side, cultivation of isolated strains, which potential will need to be fully elucidated by further studies under culture conditions closer to those of hot springs, together with chemical characterisation of the extracts, would guarantee a higher standardisation of the product and reliability of activity. To this end, future studies, including a comparison between whole
bioglea and synthetic communities built by the cultivation of multiple isolated strains under the same conditions of
bioglea, would provide decisive information to lead these interesting microbial substrates towards actual industrial-level exploitation in terms of availability of the product, quality and safety.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}