Independent and Opposite Associations Between Branched-Chain Amino Acids and Lysophosphatidylcholines With Incident Diabetes in Thais


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Ethical Approval
2.3. Biochemical Measurements
2.4. Targeted Metabolite Assessment
2.5. Statistical Analysis
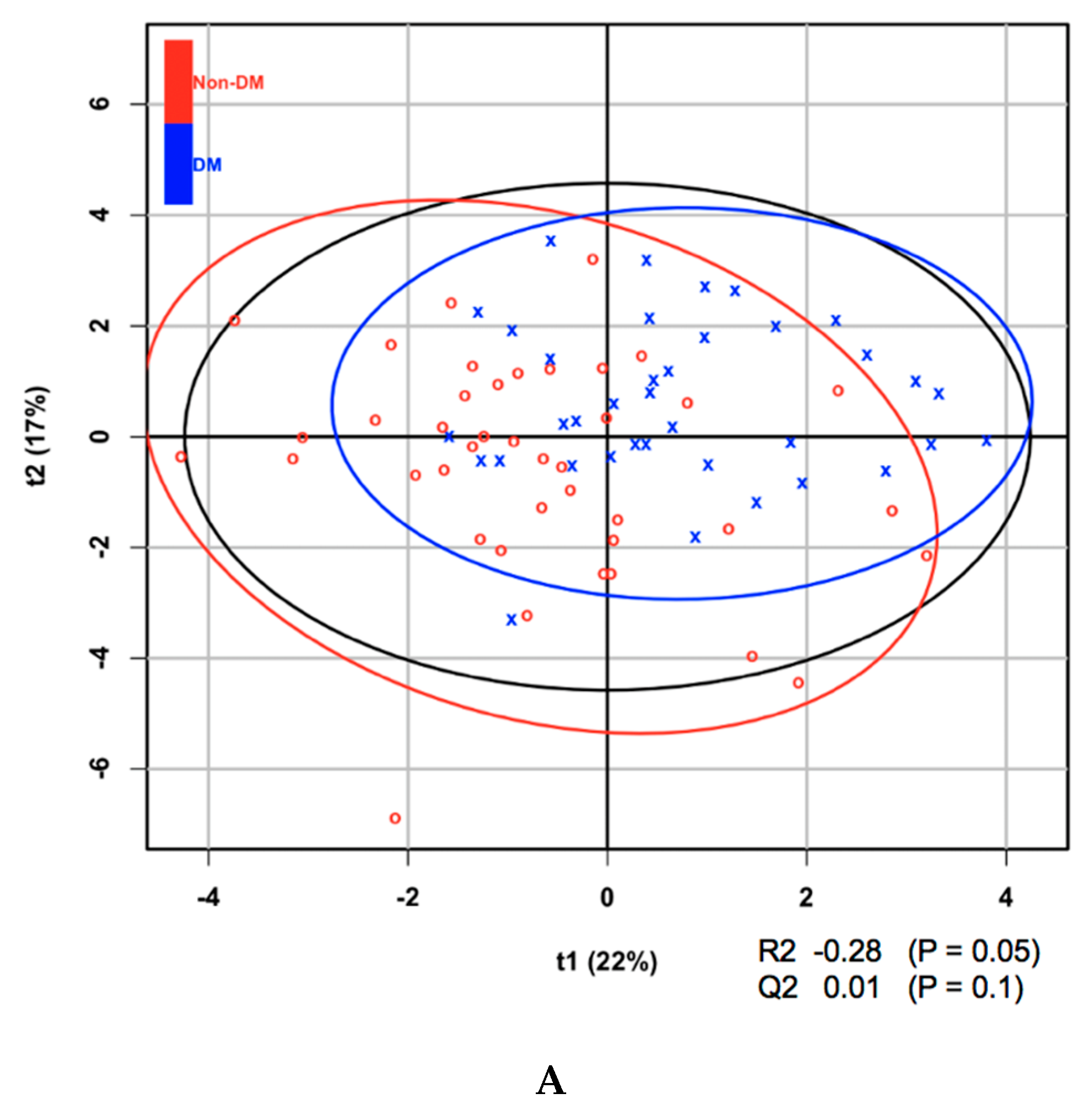
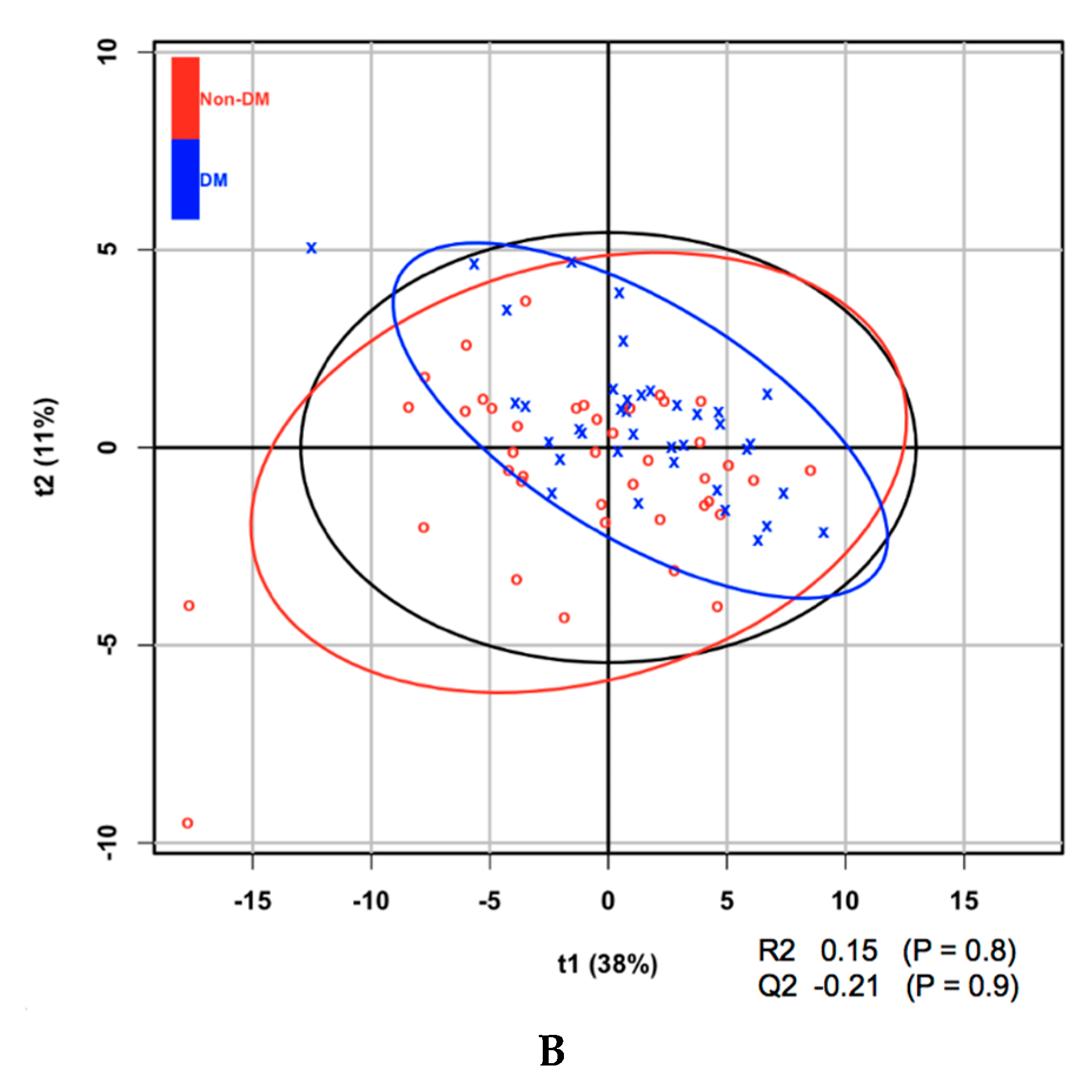
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Wurtz, P.; Soininen, P.; Kangas, A.J.; Ronnemaa, T.; Lehtimaki, T.; Kahonen, M.; Viikari, J.S.; Raitakari, O.T.; Ala-Korpela, M. Branched-chain and aromatic amino acids are predictors of insulin resistance in young adults. Diabetes Care 2013, 36, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Knockenhauer, K.E.; Wolfson, R.L.; Chantranupong, L.; Pacold, M.E.; Wang, T.; Schwartz, T.U.; Sabatini, D.M. Structural basis for leucine sensing by the Sestrin2-mTORC1 pathway. Science 2016, 351, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.F.; Cember, A.T.J.; Matschinsky, F.M. Glutamate dehydrogenase: Role in regulating metabolism and insulin release in pancreatic beta-cells. J. Appl. Physiol. 2018, 125, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Koves, T.R.; Ussher, J.R.; Noland, R.C.; Slentz, D.; Mosedale, M.; Ilkayeva, O.; Bain, J.; Stevens, R.; Dyck, J.R.; Newgard, C.B.; et al. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008, 7, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Ni, Y.; Ma, X.; Bao, Y.; Liu, J.; Huang, F.; Hu, C.; Xie, G.; Zhao, A.; Jia, W.; et al. Branched-chain and aromatic amino acid profiles and diabetes risk in Chinese populations. Sci. Rep. 2016, 6, 20594. [Google Scholar] [CrossRef] [Green Version]
- Nagata, C.; Nakamura, K.; Wada, K.; Tsuji, M.; Tamai, Y.; Kawachi, T. Branched-chain amino acid intake and the risk of diabetes in a Japanese community: The Takayama study. Am. J. Epidemiol. 2013, 178, 1226–1232. [Google Scholar] [CrossRef]
- Tai, E.S.; Tan, M.L.; Stevens, R.D.; Low, Y.L.; Muehlbauer, M.J.; Goh, D.L.; Ilkayeva, O.R.; Wenner, B.R.; Bain, J.R.; Lee, J.J.; et al. Insulin resistance is associated with a metabolic profile of altered protein metabolism in Chinese and Asian-Indian men. Diabetologia 2010, 53, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Fang, C.; Fan, Y.; Lu, Y.; Wen, B.; Ren, H.; Hou, G.; Yang, F.; Xie, H.; Jie, Z.; et al. Lipidomic profiling reveals distinct differences in plasma lipid composition in healthy, prediabetic, and type 2 diabetic individuals. Gigascience 2017, 6, gix036. [Google Scholar] [CrossRef]
- Barber, M.N.; Risis, S.; Yang, C.; Meikle, P.J.; Staples, M.; Febbraio, M.A.; Bruce, C.R. Plasma lysophosphatidylcholine levels are reduced in obesity and type 2 diabetes. PLOS ONE 2012, 7, e41456. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Lam, S.M.; Wan, Q.; Shi, L.; Huo, Y.; Chen, L.; Tang, X.; Li, B.; Wu, X.; Peng, K.; et al. High coverage targeted lipidomics reveals novel serum lipid predictors and lipid pathway dysregulation antecedent to type 2 diabetes onset in normoglycemic chinese adults. Diabetes Care 2019, 42, 2117–2126. [Google Scholar] [CrossRef]
- Wanninger, J.; Neumeier, M.; Weigert, J.; Liebisch, G.; Weiss, T.S.; Schaffler, A.; Aslanidis, C.; Schmitz, G.; Scholmerich, J.; Buechler, C. Metformin reduces cellular lysophosphatidylcholine and thereby may lower apolipoprotein B secretion in primary human hepatocytes. Biochim. Biophys. Acta 2008, 1781, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Huo, T.; Li, N.; Xiong, Z.; Li, F. Lysophosphatidylcholine--biomarker of Metformin action: Studied using UPLC/MS/MS. Biomed. Chromatogr. 2009, 23, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vathesatogkit, P.; Woodward, M.; Tanomsup, S.; Ratanachaiwong, W.; Vanavanan, S.; Yamwong, S.; Sritara, P. Cohort profile: The electricity generating authority of Thailand study. Int. J. Epidemiol. 2012, 41, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, T.; Martens, H.; Saebo, S.; Warringer, J.; Snipen, L. A Partial Least Squares based algorithm for parsimonious variable selection. Algorithms Mol. Biol. 2011, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Guerrero, J.L.; Osté, M.C.; Kieneker, L.M.; Gruppen, E.G.; Wolak-Dinsmore, J.; Otvos, J.D.; Connelly, M.A.; Bakker, S.J.; Dullaart, R.P. Plasma Branched-Chain Amino Acids and Risk of Incident Type 2 Diabetes: Results from the PREVEND Prospective Cohort Study. J. Clin. Med. 2018, 7, 513. [Google Scholar] [CrossRef] [Green Version]
- Okekunle, A.P.; Zhang, M.; Wang, Z.; Onwuka, J.U.; Wu, X.; Feng, R.; Li, C. Dietary branched-chain amino acids intake exhibited a different relationship with type 2 diabetes and obesity risk: A meta-analysis. Acta Diabetol. 2019, 56, 187–195. [Google Scholar] [CrossRef]
- Imamura, F.; Micha, R.; Khatibzadeh, S.; Fahimi, S.; Shi, P.; Powles, J.; Mozaffarian, D. Dietary quality among men and women in 187 countries in 1990 and 2010: A systematic assessment. Lancet Glob. Health 2015, 3, e132–e142. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; Lapworth, A.L.; An, J.; Wang, L.; McGarrah, R.W.; Stevens, R.D.; Ilkayeva, O.; George, T.; Muehlbauer, M.J.; Bain, J.R.; et al. Branched-chain amino acid restriction in Zucker-fatty rats improves muscle insulin sensitivity by enhancing efficiency of fatty acid oxidation and acyl-glycine export. Mol. Metab. 2016, 5, 538–551. [Google Scholar] [CrossRef]
- Cummings, N.E.; Williams, E.M.; Kasza, I.; Konon, E.N.; Schaid, M.D.; Schmidt, B.A.; Poudel, C.; Sherman, D.S.; Yu, D.; Apelo, S.I.A.; et al. Restoration of metabolic health by decreased consumption of branched-chain amino acids. J. Physiol. 2018, 596, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Saha, A.K.; Xu, X.L.; Lawson, E.; Deoliveira, R.; Brandon, A.E.; Kraegen, E.W.; Ruderman, N.B. Downregulation of AMPK accompanies leucine- and glucose-induced increases in protein synthesis and insulin resistance in rat skeletal muscle. Diabetes 2010, 59, 2426–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, F.; Huang, Z.; Li, H.; Yu, J.; Wang, C.; Chen, S.; Meng, Q.; Cheng, Y.; Gao, X.; Li, J.; et al. Leucine Deprivation Increases Hepatic Insulin Sensitivity via GCN2/mTOR/S6K1 and AMPK Pathways. Diabetes 2011, 60, 746–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, C.; Oh, S.F.; Wada, S.; Rowe, G.C.; Liu, L.; Chan, M.C.; Rhee, J.; Hoshino, A.; Kim, B.; Ibrahim, A.; et al. A branched-chain amino acid metabolite drives vascular fatty acid transport and causes insulin resistance. Nat. Med. 2016, 22, 421–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigruener, A.; Kleber, M.E.; Heimerl, S.; Liebisch, G.; Schmitz, G.; Maerz, W. Glycerophospholipid and sphingolipid species and mortality: The Ludwigshafen Risk and Cardiovascular Health (LURIC) study. PLoS ONE 2014, 9, e85724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drzazga, A.; Sowinska, A.; Koziolkiewicz, M. Lysophosphatidylcholine and lysophosphatidylinosiol--novel promissing signaling molecules and their possible therapeutic activity. Acta Pol. Pharm. 2014, 71, 887–899. [Google Scholar] [PubMed]
- Lee, J.Y.; Min, H.K.; Moon, M.H. Simultaneous profiling of lysophospholipids and phospholipids from human plasma by nanoflow liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2011, 400, 2953–2961. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Ohishi, T.; Matsui, T.; Saito, T.; Matsumoto, M.; Takasaki, J.; Matsumoto, S.; Kamohara, M.; Hiyama, H.; Yoshida, S.; et al. Lysophosphatidylcholine enhances glucose-dependent insulin secretion via an orphan G-protein-coupled receptor. Biochem. Biophys. Res. Commun. 2005, 326, 744–751. [Google Scholar] [CrossRef]
- Overton, H.A.; Babbs, A.J.; Doel, S.M.; Fyfe, M.C.; Gardner, L.S.; Griffin, G.; Jackson, H.C.; Procter, M.J.; Rasamison, C.M.; Tang-Christensen, M.; et al. Deorphanization of a G protein-coupled receptor for oleoylethanolamide and its use in the discovery of small-molecule hypophagic agents. Cell Metab. 2006, 3, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Reimann, M.; Peitzsch, M.; Ziemssen, T.; Julius, U.; Eisenhofer, G. Metabolomic distinction of microvascular effects of lipoprotein apheresis—A pilot study. Atheroscler. Suppl. 2013, 14, 143–149. [Google Scholar] [CrossRef]
- Slinker, B.K.; Glantz, S.A. Multiple regression for physiological data analysis: The problem of multicollinearity. Am. J. Physiol. 1985, 249, R1–R12. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | Mean ± Standard Deviation or Number (%) |
|---|---|
| Age (year) | 43.7 ± 6.8 |
| Male (%) | 70 (88.6%) |
| Body mass index (kg/m2) | 27.1 ± 0 4.4 |
| Fasting plasma glucose (mmol/L) | 5.97 ± 0.44 |
| Metabolite | VIP Score |
|---|---|
| Amino acid | |
| Isoleucine | 2.1 |
| Leucine | 1.8 |
| Valine | 1.6 |
| Glycerophospholipid | |
| Lysophosphatidylcholine acyl C28:1 | 1.92 |
| Phosphatidylcholine diacyl C28:1 | 1.87 |
| Phosphatidylcholine acyl-alkyl C40:1 | 1.83 |
| Phosphatidylcholine diacyl C42:4 | 1.77 |
| Phosphatidylcholine acyl-alkyl C38:2 | 1.77 |
| Phosphatidylcholine acyl-alkyl C42:5 | 1.74 |
| Phosphatidylcholine acyl-alkyl C40:5 | 1.73 |
| Phosphatidylcholine acyl-alkyl C42:4 | 1.62 |
| Phosphatidylcholine acyl-alkyl C36:1 | 1.61 |
| Phosphatidylcholine diacyl C40:3 | 1.58 |
| Phosphatidylcholine acyl-alkyl C40:3 | 1.55 |
| Phosphatidylcholine acyl-alkyl C38:0 | 1.53 |
| Phosphatidylcholine acyl-alkyl C40:2 | 1.51 |
| Variables | Non-DM (n = 41) | DM (n = 38) | p-Value | ||
|---|---|---|---|---|---|
| Mean | SE | Mean | SE | ||
| Age (year) | 43.10 | 1.11 | 43.89 | 1.05 | 0.60 |
| Male (%) | 86.10 | 88.90 | 0.68 | ||
| BMI (kg/m2) | 25.88 | 0.62 | 28.11 | 0.77 | <0.05 |
| FPG (mmol/L) | 5.75 | 0.07 | 6.13 | 0.08 | <0.001 |
| Ile (nmol/L) | 116.34 | 2.98 | 131.50 | 3.04 | <0.01 |
| Leu (nmol/L) | 264.89 | 9.18 | 298.39 | 9.01 | <0.05 |
| Val (nmol/L) | 320.00 | 5.67 | 339.50 | 5.46 | <0.05 |
| Lysophosphatidylcholine acyl C28:1 (nmol/L) | 0.27 | 0.01 | 0.23 | 0.01 | <0.05 |
| Phosphatidylcholine diacyl C28:1 (nmol/L) | 2.33 | 0.09 | 2.06 | 0.08 | <0.05 |
| Phosphatidylcholine acyl-alkyl C40:1 (nmol/L) | 0.40 | 0.01 | 0.38 | 0.01 | 0.29 |
| Phosphatidylcholine diacyl C42:4 (nmol/L) | 0.68 | 0.03 | 0.59 | 0.02 | <0.05 |
| Phosphatidylcholine acyl-alkyl C38:1 (nmol/L) | 2.73 | 0.29 | 2.82 | 0.28 | 0.82 |
| Phosphatidylcholine acyl-alkyl C42:5 (nmol/L) | 0.28 | 0.02 | 0.26 | 0.01 | 0.41 |
| Phosphatidylcholine acyl-alkyl C40:5 (nmol/L) | 9.15 | 0.52 | 8.49 | 0.45 | 0.34 |
| Phosphatidylcholine acyl-alkyl C42:4 (nmol/L) | 0.22 | 0.02 | 0.18 | 0.01 | <0.05 |
| Phosphatidylcholine acyl-alkyl C36:1 (nmol/L) | 39.89 | 1.55 | 38.68 | 1.52 | 0.58 |
| Phosphatidylcholine diacyl C40:3 (nmol/L) | 1.36 | 0.20 | 0.94 | 0.06 | 0.06 |
| Phosphatidylcholine acyl-alkyl C40:3 (nmol/L) | 0.47 | 0.02 | 0.41 | 0.02 | 0.05 |
| Phosphatidylcholine acyl-alkyl C38:0 (nmol/L) | 2.80 | 0.11 | 2.70 | 0.12 | 0.53 |
| Phosphatidylcholine acyl-alkyl C40:2 (nmol/L) | 0.40 | 0.01 | 0.38 | 0.01 | 0.29 |
| Variable | Leucine | Isoleucine | Valine | LPC C28:1 | Age | Sex | BMI |
|---|---|---|---|---|---|---|---|
| Leucine | 1 | ||||||
| Isoleucine | 0.87 p < 0.001 | 1 | |||||
| Valine | 0.68 p < 0.001 | 0.69 p < 0.01 | 1 | ||||
| LPC C28:1 | 0.06 p = 0.59 | 0.05 p = 0.6 | 0.02 p = 0.88 | 1 | |||
| Age | −0.27 p < 0.05 | −0.21 p = 0.06 | −0.19 p = 0.09 | 0.08 p = 0.46 | 1 | ||
| Sex | −0.13 p = 0.23 | −0.15 p = 0.18 | −0.08 p = 0.46 | −0.15 p = 0.18 | −0.002 p = 0.99 | 1 | |
| BMI | 0.30 p < 0.01 | 0.31 p < 0.05 | 0.30 p < 0.01 | −0.15 p = 0.17 | −0.16 p = 0.14 | 0.24 p < 0.05 | 1 |
| Variables | Odds Ratio | 95% Confidence Interval | p-Value |
|---|---|---|---|
| Isoleucine | 1.00025 | 1.000003–1.00051 | <0.05 |
| LPC acyl C28:1 | 0.976 | 0.972–0.999 | <0.05 |
| Baseline FPG | 1.080 | 1.009–1.057 | <0.05 |
| Baseline age | 1.567 | 0.965–1.157 | 0.24 |
| Baseline BMI | 1.082 | 0.938–1.249 | 0.28 |
| Female | 0.936 | 0.140–6.256 | 0.95 |
| Variable | Coefficient (×10−3) | p-Value |
|---|---|---|
| Isoleucine | 0.095 | <0.05 |
| LPC acyl C28:1 | −5.15 | <0.05 |
| Baseline FPG | 31.91 | <0.05 |
| Baseline age | 32.81 | <0.05 |
| Baseline BMI | 57.40 | 0.052 |
| Female | −346.20 | 0.25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chailurkit, L.-o.; Paiyabhroma, N.; Sritara, P.; Vathesatogkit, P.; Yamwong, S.; Thonmung, N.; Ongphiphadhanakul, B. Independent and Opposite Associations Between Branched-Chain Amino Acids and Lysophosphatidylcholines With Incident Diabetes in Thais. Metabolites 2020, 10, 76. https://doi.org/10.3390/metabo10020076
Chailurkit L-o, Paiyabhroma N, Sritara P, Vathesatogkit P, Yamwong S, Thonmung N, Ongphiphadhanakul B. Independent and Opposite Associations Between Branched-Chain Amino Acids and Lysophosphatidylcholines With Incident Diabetes in Thais. Metabolites. 2020; 10(2):76. https://doi.org/10.3390/metabo10020076
Chicago/Turabian StyleChailurkit, La-or, Nitchawat Paiyabhroma, Piyamit Sritara, Prin Vathesatogkit, Sukit Yamwong, Nisakron Thonmung, and Boonsong Ongphiphadhanakul. 2020. "Independent and Opposite Associations Between Branched-Chain Amino Acids and Lysophosphatidylcholines With Incident Diabetes in Thais" Metabolites 10, no. 2: 76. https://doi.org/10.3390/metabo10020076
APA StyleChailurkit, L. -o., Paiyabhroma, N., Sritara, P., Vathesatogkit, P., Yamwong, S., Thonmung, N., & Ongphiphadhanakul, B. (2020). Independent and Opposite Associations Between Branched-Chain Amino Acids and Lysophosphatidylcholines With Incident Diabetes in Thais. Metabolites, 10(2), 76. https://doi.org/10.3390/metabo10020076