Untargeted Metabolomics and Steroid Signatures in Urine of Male Pattern Baldness Patients after Finasteride Treatment for a Year

 ,
,  and
and
Abstract
:1. Introduction
2. Results
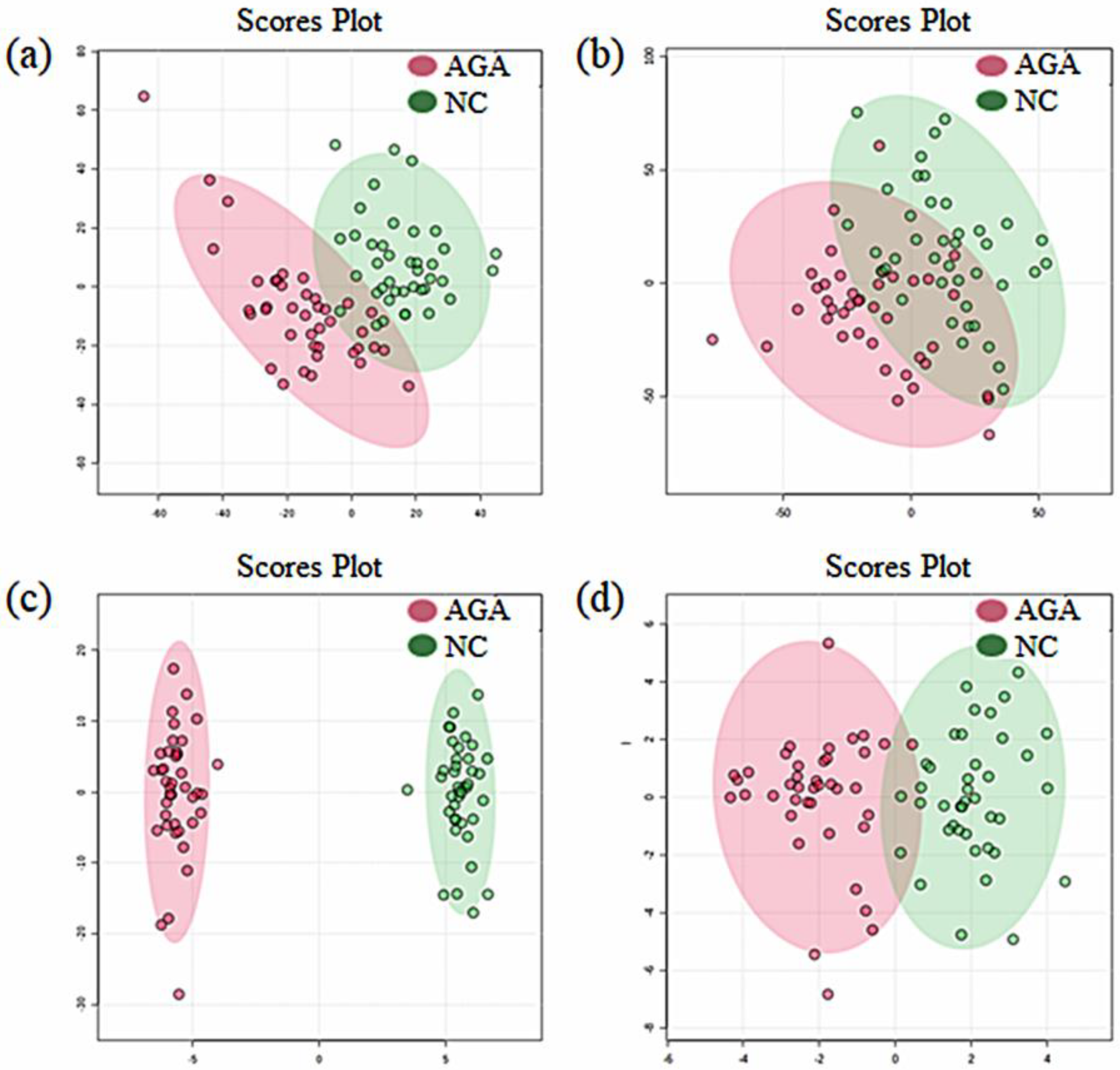
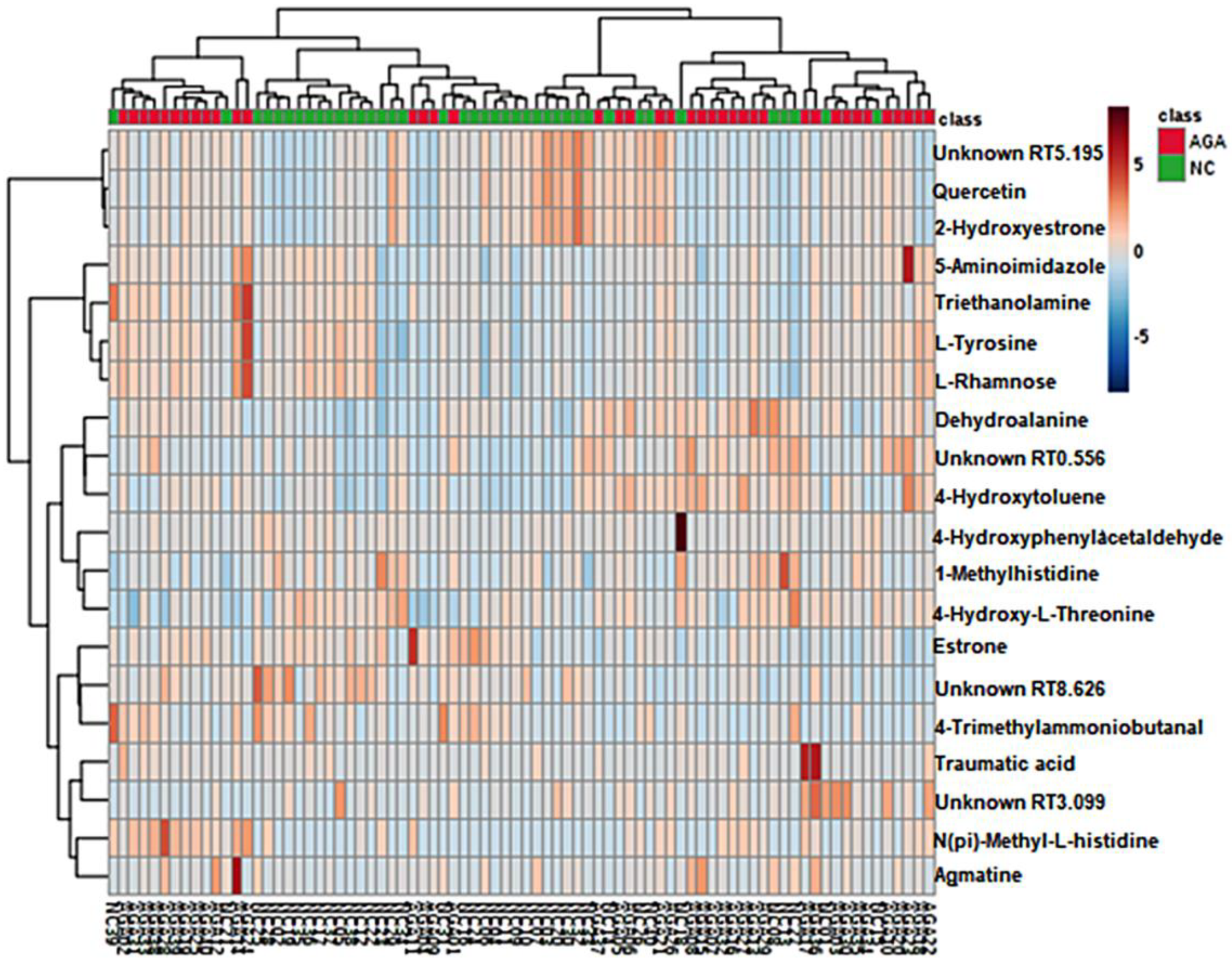
2.1. Pattern Discovery in the Urine Samples of MPB Patients Using Untargeted Metabolomics
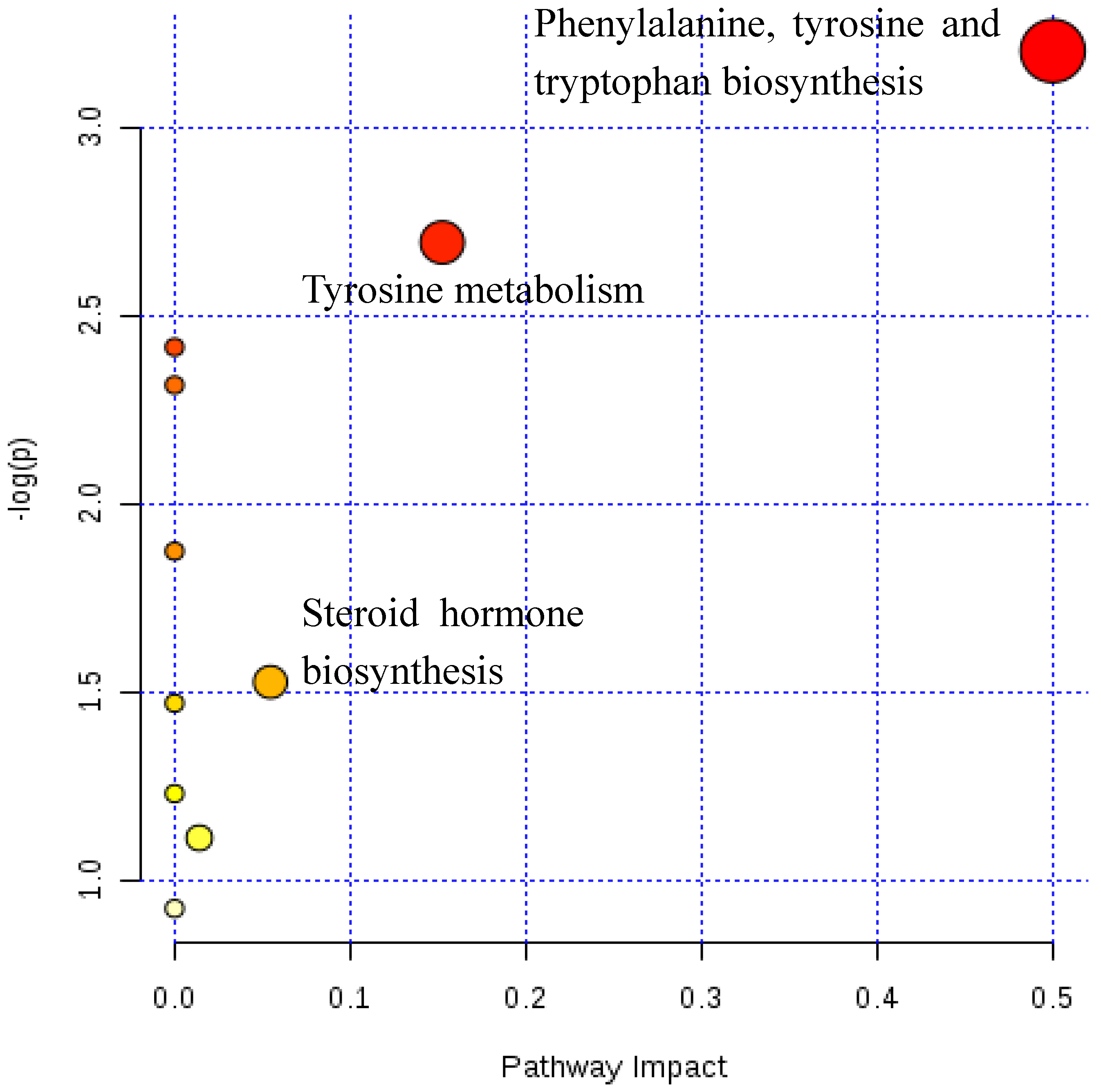
2.2. Metabolic Pathway Analysis
2.3. Quantification of Androgens and Estrogens in Urine Samples of MPB Patients Who Have Been Taking Finasteride for One Year
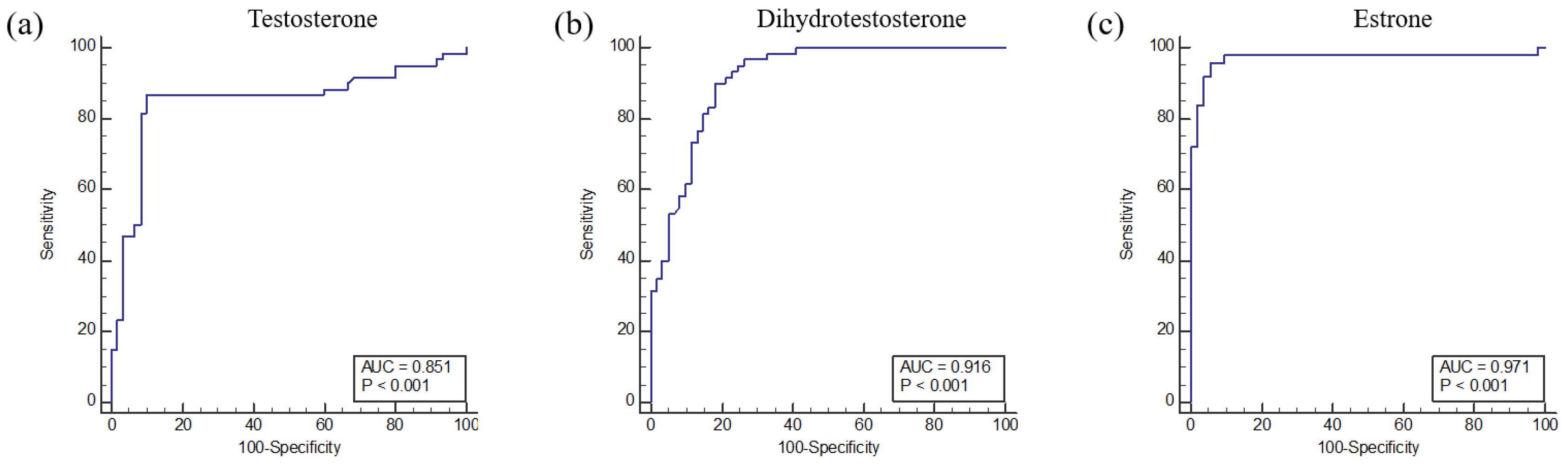
2.4. Receiver Operating Characteristic Curve
2.5. Method Reproducibility
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Urine Sample Collection
4.3. Sample Preparation for the Untargeted Approach
4.4. Ultra-Performance Liquid Chromatography-Mass Spectrometry (UPLC-MS)
4.5. Sample Preparation and Pretreatment for Targeted Profiling
4.6. Liquid Chromatography-Tandem Mass Spectrometry for Targeted Profiling
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Choi, M.H.; Kim, S.J.; Lew, B.L.; Sim, W.Y.; Chung, B.C. Hair steroid profiling reveals racial differences in male pattern baldness between Korean and Caucasian populations. J. Investig. Dermatol. 2013, 133, 822–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, H.J.; Yang, Y.J.; Lho, D.S.; Lee, W.Y.; Sim, W.Y.; Chung, B.C. Comparative studies on level of androgens in hair and plasma with premature male-pattern baldness. J. Dermatol. Sci. 2004, 34, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.H.; Yoo, Y.S.; Chung, B.C. Biochemical roles of testosterone and epitestosterone to 5 alpha-reductase as indicators of male-pattern baldness. J. Investig. Dermatol. 2001, 116, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, S.A. 5alpha-reductase inhibitors: What role should they play? Urology 2001, 58, 65–70. [Google Scholar] [CrossRef]
- Rhodes, L.; Harper, J.; Uno, H.; Gaito, G.; Audette-Arruda, J.; Kurata, S.; Berman, C.; Primka, R.; Pikounis, B. The effects of finasteride (Proscar) on hair growth, hair cycle stage, and serum testosterone and dihydrotestosterone in adult male and female stumptail macaques (Macaca arctoides). J. Clin. Endocrinol. Metab. 1994, 79, 991–996. [Google Scholar] [CrossRef]
- Drake, L.; Hordinsky, M.; Fiedler, V.; Swinehart, J.; Unger, W.P.; Cotterill, P.C.; Thiboutot, D.M.; Lowe, N.; Jacobson, C.; Whiting, D.; et al. The effects of finasteride on scalp skin and serum androgen levels in men with androgenetic alopecia. J. Am. Acad. Dermatol. 1999, 41, 550–554. [Google Scholar]
- Van Neste, D.; Fuh, V.; Sanchez-Pedreno, P.; Lopez-Bran, E.; Wolff, H.; Whiting, D.; Roberts, J.; Kopera, D.; Stene, J.J.; Calvieri, S.; et al. Finasteride increases anagen hair in men with androgenetic alopecia. Br. J. Dermatol. 2000, 143, 804–810. [Google Scholar] [CrossRef]
- Kaufman, K.D.; Olsen, E.A.; Whiting, D.; Savin, R.; DeVillez, R.; Bergfeld, W.; Price, V.H.; Van Neste, D.; Roberts, J.L.; Hordinsky, M.; et al. Finasteride in the treatment of men with androgenetic alopecia. Finasteride Male Pattern Hair Loss Study Group. J. Am. Acad. Dermatol. 1998, 39, 578–589. [Google Scholar] [CrossRef]
- Price, V.H.; Menefee, E.; Sanchez, M.; Ruane, P.; Kaufman, K.D. Changes in hair weight and hair count in men with androgenetic alopecia after treatment with finasteride, 1 mg, daily. J. Am. Acad. Dermatol. 2002, 46, 517–523. [Google Scholar] [CrossRef]
- Stough, D.B.; Rao, N.A.; Kaufman, K.D.; Mitchell, C. Finasteride improves male pattern hair loss in a randomized study in identical twins. Eur. J. Dermatol. 2002, 12, 32–37. [Google Scholar]
- Ryu, H.K.; Kim, K.M.; Yoo, E.A.; Sim, W.Y.; Chung, B.C. Evaluation of androgens in the scalp hair and plasma of patients with male-pattern baldness before and after finasteride administration. Br. J. Dermatol. 2006, 154, 730–734. [Google Scholar] [CrossRef] [PubMed]
- Carani, C.; Qin, K.; Simoni, M.; Faustini-Fustini, M.; Serpente, S.; Boyd, J.; Korach, K.S.; Simpson, E.R. Effect of testosterone and estradiol in a man with aromatase deficiency. N. Engl. J. Med. 1997, 337, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Kawakami, M.; Robert, M.; Soga, T.; Tomita, M. Bioinformatics Tools for Mass Spectroscopy-Based Metabolomic Data Processing and Analysis. Curr. Bioinform. 2012, 7, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metabolomics 2013, 1, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Tikunov, Y.M.; Laptenok, S.; Hall, R.D.; Bovy, A.; de Vos, R.C. MSClust: A tool for unsupervised mass spectra extraction of chromatography-mass spectrometry ion-wise aligned data. Metabolomics 2012, 8, 714–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, J.E.; Uzqueda, H.R. The influence of citrate and phosphate on the Mancini single radial immunodiffusion technique and suggested improvements for the determination of urinary albumin. Clin. Chim. Acta 1978, 90, 249–257. [Google Scholar] [CrossRef]
- Im, E.; Lew, B.L.; Lee, M.Y.; Lee, J.; Paeng, K.J.; Chung, B.C. Simultaneous determination of androgens and prostaglandins in human urine using ultra-high-performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2019, 1109, 45–53. [Google Scholar] [CrossRef]
- Gowda, D.; Premalatha, V.; Imtiyaz, D.B. Prevalence of Nutritional Deficiencies in Hair Loss among Indian Participants: Results of a Cross-sectional Study. Int. J. Trichology 2017, 9, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.R.; Lew, B.L.; Sim, W.Y.; Lee, J.; Hong, J.; Chung, B.C. Altered polyamine profiling in the hair of patients with androgenic alopecia and alopecia areata. J. Dermatol. 2019, 46, 985–992. [Google Scholar] [CrossRef]
- Lee, Y.R.; Kim, H.; Lew, B.L.; Sim, W.Y.; Lee, J.; Oh, H.B.; Hong, J.; Chung, B.C. Sex-related differences in urinary immune-related metabolic profiling of alopecia areata patients. Metabolomics 2020, 16, 15. [Google Scholar] [CrossRef]
- Shapiro, I.P.; Masliah, E.; Saitoh, T. Altered protein tyrosine phosphorylation in Alzheimer’s disease. J. Neurochem. 1991, 56, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kurup, P.; Nairn, A.C.; Lombroso, P.J. Striatal-enriched protein tyrosine phosphatase in Alzheimer’s disease. Adv. Pharmacol. 2012, 64, 303–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Magana, M.; Angulo, M.; Fuentes, B.; Canas, A.; Sarrantonio, M.; Arguello, R.; Vitollo, P. Effect of finasteride on human testicular steroidogenesis. J. Androl. 1996, 17, 516–521. [Google Scholar]
- Caruso, D.; Abbiati, F.; Giatti, S.; Romano, S.; Fusco, L.; Cavaletti, G.; Melcangi, R.C. Patients treated for male pattern hair with finasteride show, after discontinuation of the drug, altered levels of neuroactive steroids in cerebrospinal fluid and plasma. J. Steroid Biochem. Mol. Biol. 2015, 146, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Melcangi, R.C.; Santi, D.; Spezzano, R.; Grimoldi, M.; Tabacchi, T.; Fusco, M.L.; Diviccaro, S.; Giatti, S.; Carra, G.; Caruso, D.; et al. Neuroactive steroid levels and psychiatric and andrological features in post-finasteride patients. J. Steroid Biochem. Mol. Biol. 2017, 171, 229–235. [Google Scholar] [CrossRef]
- Melcangi, R.C.; Caruso, D.; Abbiati, F.; Giatti, S.; Calabrese, D.; Piazza, F.; Cavaletti, G. Neuroactive steroid levels are modified in cerebrospinal fluid and plasma of post-finasteride patients showing persistent sexual side effects and anxious/depressive symptomatology. J. Sex. Med. 2013, 10, 2598–2603. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.E.; Ponder, B.; Ross, R.K. Hormones, Genes, and Cancer. Cancer. 2003, 98, 2521. [Google Scholar] [CrossRef]
- Drury, J.E.; Di Costanzo, L.; Penning, T.M.; Christianson, D.W. Inhibition of human steroid 5beta-reductase (AKR1D1) by finasteride and structure of the enzyme-inhibitor complex. J. Biol. Chem. 2009, 284, 19786–19790. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Kim, S.J.; Lee, W.Y.; Chung, B.C.; Choi, M.H. Gas chromatography/mass spectrometry based hair steroid profiling may reveal pathogenesis in hair follicles of the scalp. Rapid Commun. Mass Spectrom. 2011, 25, 1184–1192. [Google Scholar] [CrossRef]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genomics 2006, 7, 142. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Normal Controls (n = 60) | MPB Patients (n = 63) | p-Value | ||
|---|---|---|---|---|---|
| Mean ± SD | Median, Range | Mean ± SD | Median, Range | ||
| T | 15.2 ± 40.88 | 4.18, 0.3 – 247.37 | 120.53 ± 135.05 | 67.73, 0.29 – 580.73 | 1.04−7 |
| DHT | 1.81 ± 2.9 | 0.58, 0.06 – 12.93 | 9.43 ± 6.50 | 7.95, 0.85 – 28.62 | 9.19−12 |
| EpiT | 332.37 ± 306.86 | 246.71, 7.28 – 1,376.31 | 1603.89 ± 2676.78 | 58.53, 8.75 – 8,518.28 | 0.08 |
| DHEA | 7606.52 ± 13,258.45 | 2833.75, 8.49 – 61,246 | 7549.49 ± 11,681.70 | 1560.56, 46.11 – 34,394.64 | 0.86 |
| E1 | 3.71 ± 3.12 | 2.99, 0.21 – 16.9 | 39.54 ± 28.73 | 29.04, 11.43 – 116.01 | 5.21−9 |
| E2 | 13.89 ± 10.60 | 11.18, 0.04 – 39.51 | 24.56 ± 17.71 | 17.49, 1.66 – 75.13 | 0.001 |
| E3 | 85.07 ± 71.49 | 72.63, 26.9 – 292.11 | 61.30 ± 24.13 | 73.39, 31.95 – 94.14 | 0.31 |
| DHT/T ratio | 0.25 ± 0.32 | 0.14, 0.02 – 1.73 | 0.16 ± 0.22 | 0.1, 0.02 – 1.08 | 0.06 |
| Category | MPB Patients | Controls |
|---|---|---|
| Patients number | 63 | 60 |
| Age at diagnosis of MPB, Mean ± SD | 31.59 ± 8 | 27.63 ± 4.8 |
| Age range | 18 ~ 51 | 20 ~ 39 |
| Nationality, n (%) | Korean (100) | Korean (100) |
| N-H classification, n (%) | ||
| F1 | 12 (19) | |
| F2 | 3 (5) | |
| II | 20 (32) | |
| III | 22 (35) | |
| IV | 4 (6) | |
| V | 2 (3) | |
| MPB duration, Mean ± SD | 5.29 ± 4.01 | |
| Past Tx Hx (-) | None (100) | |
| Present Tx | Finasteride (100) | |
| Other chronic illness, n (%) | ||
| None | 60 (95) | |
| HBV carrier | 1 (1.6) | |
| Liver cirrhosis | 1 (1.6) | |
| Thyroid cancer, kidney cancer | 1 (1.6) | |
| Other dermatitis diseases, n (%) | ||
| None | 59 (93) | |
| Acne | 1 (1.6) | |
| Alopecia areata | 1 (1.6) | |
| Seborrheic dermatitis | 2 (3.2) |
| Compound | Precursor Ion (m/z) | Product Iona (m/z, Collision Energy) |
|---|---|---|
| Androgens | ||
| T | 380.2 | 272.1 (45), 134.1 (47) |
| DHT | 382.1 | 288.2 (33), 365.2 (33) |
| EpiT | 380.2 | 272.1 (43), 134.1 (47) |
| DHEA | 380.1 | 268.1 (35), 209.1 (41) |
| d6-DHEA (IS) | 386.2 | 274.1 (37), 227.1 (41) |
| Estrogens | ||
| E1 | 504.3 | 440.3 (39), 425.4 (22) |
| E2 | 506.4 | 442.3 (38), 427.3(21) |
| E3 | 522.3 | 458.3 (30), 443.3(24) |
| d4-E2 (IS) | 510.3 | 466.3 (21), 431.3 (21) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.R.; Im, E.; Kim, H.; Lew, B.L.; Sim, W.-Y.; Lee, J.; Oh, H.B.; Paeng, K.J.; Hong, J.; Chung, B.C. Untargeted Metabolomics and Steroid Signatures in Urine of Male Pattern Baldness Patients after Finasteride Treatment for a Year. Metabolites 2020, 10, 131. https://doi.org/10.3390/metabo10040131
Lee YR, Im E, Kim H, Lew BL, Sim W-Y, Lee J, Oh HB, Paeng KJ, Hong J, Chung BC. Untargeted Metabolomics and Steroid Signatures in Urine of Male Pattern Baldness Patients after Finasteride Treatment for a Year. Metabolites. 2020; 10(4):131. https://doi.org/10.3390/metabo10040131
Chicago/Turabian StyleLee, Yu Ra, Eunju Im, Haksoon Kim, Bark Lynn Lew, Woo-Young Sim, Jeongae Lee, Han Bin Oh, Ki Jung Paeng, Jongki Hong, and Bong Chul Chung. 2020. "Untargeted Metabolomics and Steroid Signatures in Urine of Male Pattern Baldness Patients after Finasteride Treatment for a Year" Metabolites 10, no. 4: 131. https://doi.org/10.3390/metabo10040131
APA StyleLee, Y. R., Im, E., Kim, H., Lew, B. L., Sim, W. -Y., Lee, J., Oh, H. B., Paeng, K. J., Hong, J., & Chung, B. C. (2020). Untargeted Metabolomics and Steroid Signatures in Urine of Male Pattern Baldness Patients after Finasteride Treatment for a Year. Metabolites, 10(4), 131. https://doi.org/10.3390/metabo10040131