Proteobacteria Overgrowth and Butyrate-Producing Taxa Depletion in the Gut Microbiota of Glycogen Storage Disease Type 1 Patients

 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Cohort Description
2.2. Dietary Assessment
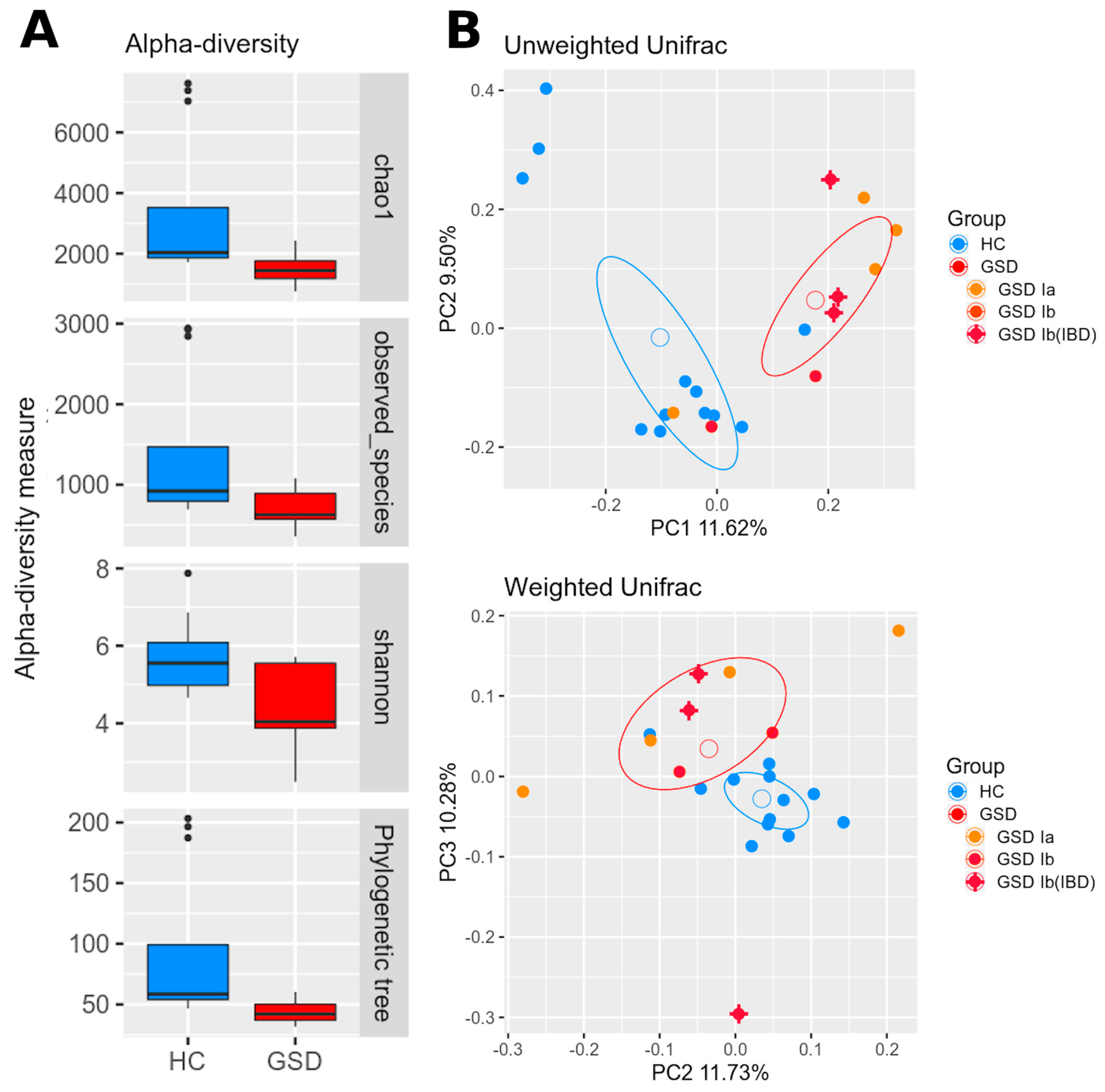
2.3. Microbiota Profiling
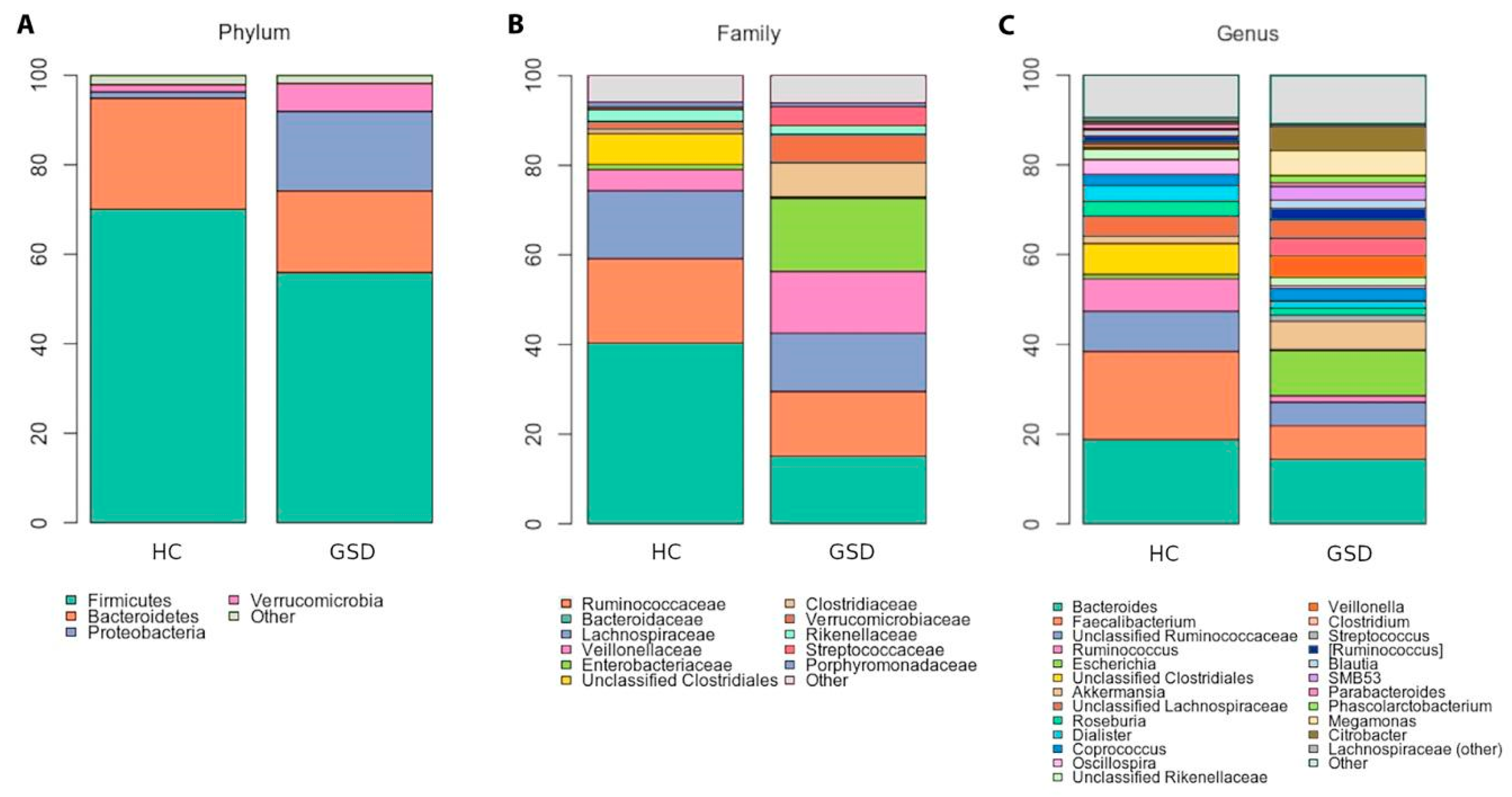
2.3.1. Taxonomic Characterization
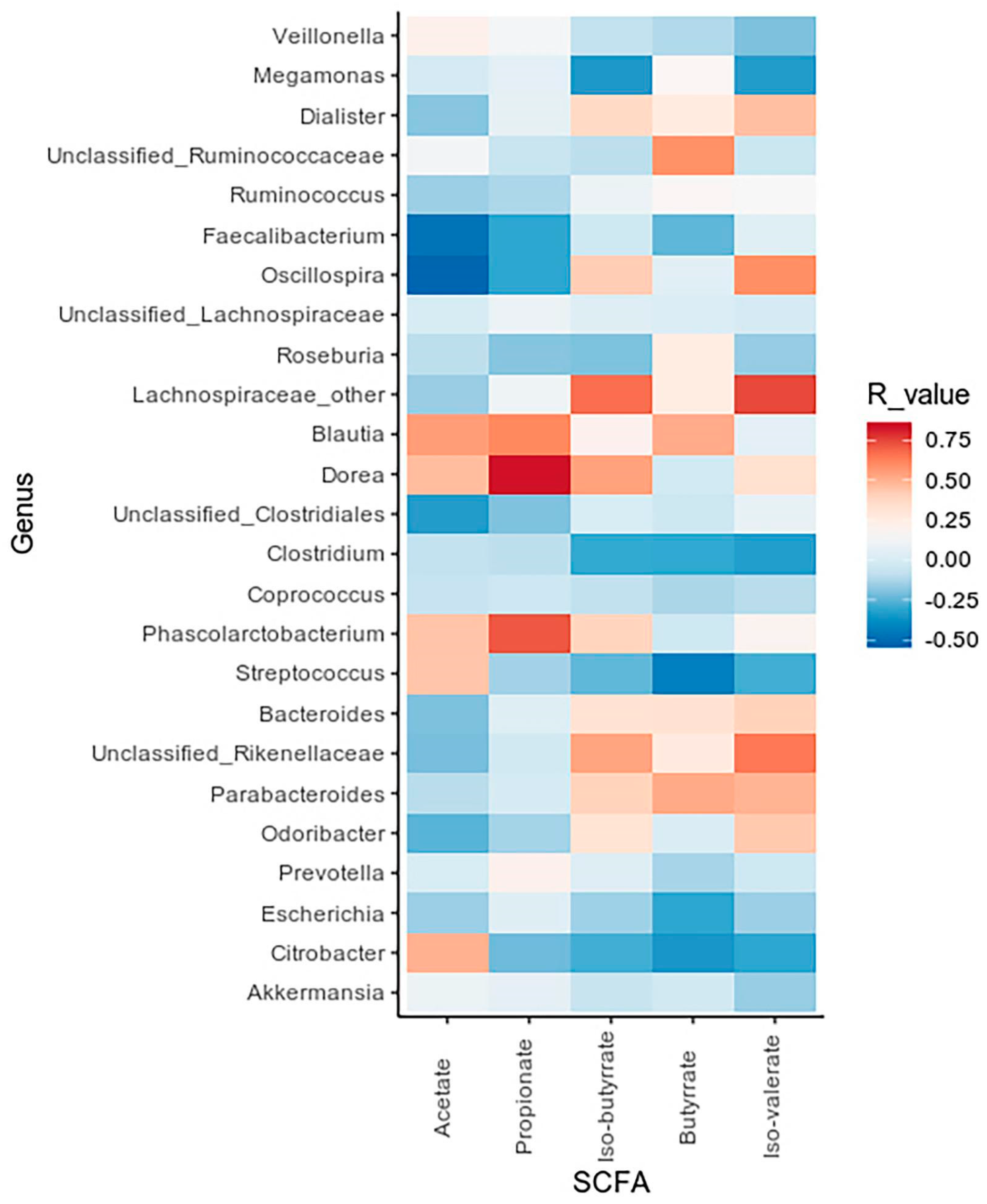
2.3.2. Fecal Microbial Metabolites
2.3.3. Functional Prediction
2.4. Relationship between Microbial Population, Metabolite Content and Diet
2.4.1. Gut Microbiota and Fecal Microbial Metabolites
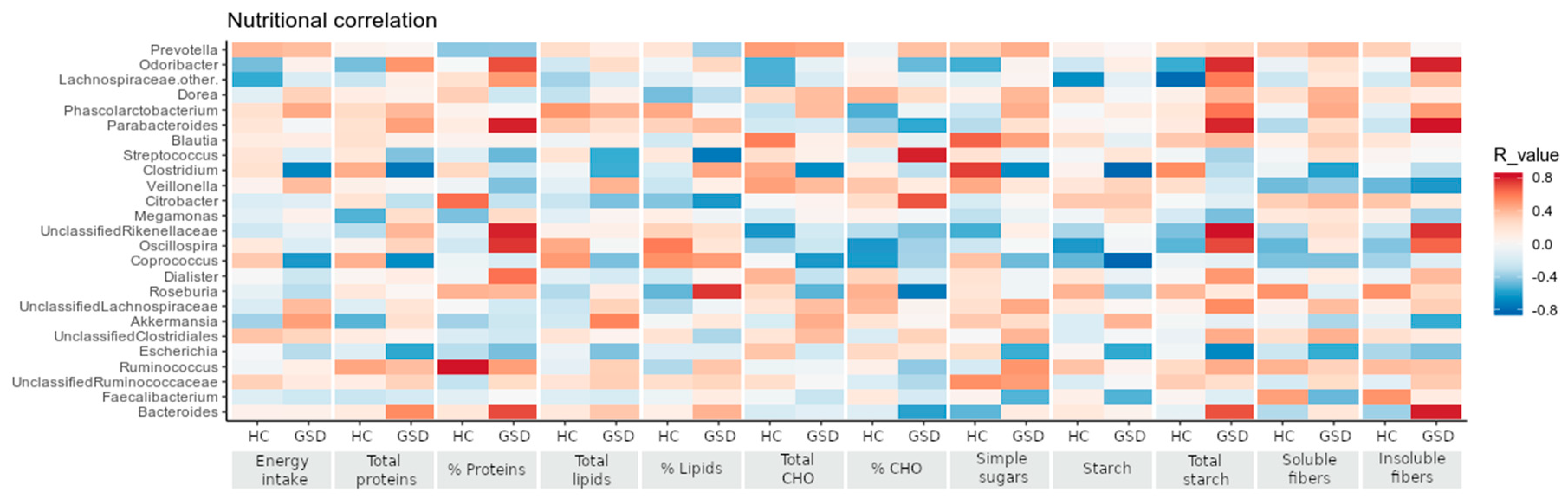
2.4.2. Impact of Diet on Microbial Taxa Relative Abundance
3. Discussion
4. Materials and Methods
4.1. Subject Recruitment and Sampling
4.2. Gut Microbial DNA Extraction and Sequencing
4.3. Microbiota Profiling
4.4. Fecal Short Chain Fatty Acids Measurement
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ding, R.X.; Goh, W.R.; Wu, R.N.; Yue, X.Q.; Luo, X.; Khine, W.W.T.; Wu, J.R.; Lee, Y.K. Revisit gut microbiota and its impact on human health and disease. J. Food Drug Anal. 2019, 27, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Marchesi, N.; Marelli, C.; Coppola, A.; Luzi, L.; Govoni, S.; Giustina, A.; Gazzaruso, C. Microbiota and metabolic diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Heller, S.; Worona, L.; Consuelo, A. Nutritional therapy for glycogen storage diseases. J. Pediatr. Gastroenterol. Nutr. 2008, 47, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kanungo, S.; Wells, K.; Tribett, T.; El-Gharbawy, A. Glycogen metabolism and glycogen storage disorders. Ann. Transl. Med. 2018, 6, 474. [Google Scholar] [CrossRef]
- Beyzaei, Z.; Geramizadeh, B. Molecular diagnosis of glycogen storage disease type I: A review. EXCLI J. 2019, 18, 30–46. [Google Scholar]
- Chen, Y.T. Glycogen storage diseases. In The Metabolic and Molecular Bases of Inherited Disease, 8th ed.; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Childs, B., Kinzler, K.W., Vogelstein, B., Eds.; McGraw-Hill Education-Europe: New York, NY, USA, 2001; pp. 1521–1555. [Google Scholar]
- Santos, B.L.; Souza, C.F.; Schuler-Faccini, L.; Refosco, L.; Epifanio, M.; Nalin, T.; Vieira, S.M.; Schwartz, I.V. Glycogen storage disease type I: Clinical and laboratory profile. J. Pediatr. (Rio J) 2014, 90, 572–579. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, N.; Gautschi, M.; Bosanska, L.; Meienberg, F.; Baumgartner, M.R.; Spinas, G.A.; Hochuli, M. Glycemic control and complications in glycogen storage disease type I: Results from the Swiss registry. Mol. Genet. Metab. 2019, 126, 355–361. [Google Scholar] [CrossRef]
- Froissart, R.; Piraud, M.; Boudjemline, A.M.; Vianey-Saban, C.; Petit, F.; Hubert-Buron, A.; Eberschweiler, P.T.; Gajdos, V.; Labrune, P. Glucose-6-phosphatase deficiency. Orphanet J. Rare Dis. 2011, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.Y.; Jun, H.S.; Mansfield, B.C. Type I glycogen storage diseases: Disorders of the glucose-6-phosphatase/glucose-6-phosphate transporter complexes. J. Inherit. Metab. Dis. 2015, 38, 511–519. [Google Scholar] [CrossRef]
- Ozen, H. Glycogen storage diseases: New perspectives. World J. Gastroenterol. 2007, 13, 2541–2553. [Google Scholar] [CrossRef]
- Derks, T.G.; Martens, D.H.; Sentner, C.P.; van Rijn, M.; de Boer, F.; Smit, G.P.; van Spronsen, F.J. Dietary treatment of glycogen storage disease type Ia: Uncooked cornstarch and/or continuous nocturnal gastric drip-feeding? Mol. Genet. Metab. 2013, 109, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, N.T.; Chengsupanimit, T.; Brown, L.M.; Derks, T.G.; Smit, G.P.; Weinstein, D.A. Inflammatory Bowel Disease in Glycogen Storage Disease Type Ia. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 52–54. [Google Scholar] [CrossRef] [PubMed]
- Carnero-Gregorio, M.; Molares-Vila, A.; Corbalán-Rivas, A.; Villaverde-Taboada, C.; Rodríguez-Cerdeira, C. Effect of VSL#3 Probiotic in a Patient with Glycogen Storage Disease Type Ia and Irritable Bowel Disease-like Disease. Probiotics Antimicro. Prot. 2019, 11, 143–149. [Google Scholar] [CrossRef]
- Kishnani, P.S.; Austin, S.L.; Abdenur, J.E.; Arn, P.; Bali, D.S.; Boney, A.; Chung, W.K.; Dagli, A.I.; Dale, D.; Koeberl, D.; et al. Diagnosis and management of glycogen storage disease type I: A practice guideline of the American College of Medical Genetics and Genomics. Genet. Med. 2014, 16, e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.A.; Weinstein, D.A. Glycogen storage diseases: Diagnosis, treatment and outcome. Transl. Sci. Rare Dis. 2016, 1, 45–72. [Google Scholar] [CrossRef] [Green Version]
- Rake, J.P.; Visser, G.; Labrune, P.; Leonard, J.V.; Ullrich, K.; Smit, G.P. Glycogen storage disease type I: Diagnosis, management, clinical course and outcome. Results of the European Study on Glycogen Storage Disease Type I (ESGSD I). Eur. J. Pediatr. 2002, 161, 20–34. [Google Scholar] [CrossRef]
- Kishnani, P.S.; Koeberl, D.; Chen, Y.T. Glycogen storage diseases. In Pediatric Nutrition in Chronic Diseases and Developmental Disorders: Prevention, Assessment, and Treatment, 2nd ed.; Ekvall, S.W., Ekvall, V.K., Eds.; Oxford University Press: New York, NY, USA, 2005; pp. 350–355. [Google Scholar]
- Goldberg, T.; Slonim, A.E. Nutrition therapy for hepatic glycogen storage diseases. J. Am. Diet. Assoc. 1993, 93, 1423–1430. [Google Scholar] [CrossRef]
- Burda, P.; Hochuli, M. Hepatic glycogen storage disorders: What have we learned in recent years? Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Bassanini, G.; Ceccarani, C.; Borgo, F.; Severgnini, M.; Rovelli, V.; Morace, G.; Verduci, E.; Borghi, E. Phenylketonuria Diet Promotes Shifts in Firmicutes Populations. Front. Cell. Infect. Microbiol. 2019, 9, 101. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazar, V.; Ditu, L.M.; Pircalabioru, G.G.; Gheorghe, I.; Curutiu, C.; Holban, A.M.; Picu, A.; Petcu, L.; Chifiriuc, M.C. Aspects of Gut Microbiota and Immune System Interactions in Infectious Diseases, Immunopathology, and Cancer. Front. Immunol. 2018, 9, 1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colonetti, K.; Bento Dos Santos, B.; Nalin, T.; Moura de Souza, C.F.; Triplett, E.W.; Dobbler, P.T.; Schwartz, I.V.D.; Roesch, L.F.W. Hepatic glycogen storage diseases are associated to microbial dysbiosis. PLoS ONE 2019, 14, e0214582. [Google Scholar] [CrossRef]
- Webb, C.R.; Koboziev, I.; Furr, K.L.; Grisham, M.B. Protective and Pro-Inflammatory Roles of Intestinal Bacteria. Pathophysiol 2016, 23, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, G.P.; Booth, I.R. Importance of Glutathione for Growth and Survival of Escherichia coli Cells: Detoxification of Methylglyoxal and Maintenance of Intracellular K+. J. Bacteriol. 1998, 180, 4314–4318. [Google Scholar] [CrossRef] [Green Version]
- Kitamoto, S.; Alteri, C.J.; Rodrigues, M.; Nagao-Kitamoto, H.; Sugihara, K.; Himps, S.D.; Bazzi, M.; Miyoshi, M.; Nishioka, T.; Hayashi, A.; et al. Dietary l-serine confers a competitive fitness advantage to Enterobacteriaceae in the inflamed gut. Nat. Microbiol. 2020, 5, 116–125. [Google Scholar] [CrossRef]
- Tuovinen, E.; Keto, J.; Nikkilä, J.; Mättö, J.; Lähteenmäki, K. Cytokine response of human mononuclear cells induced by intestinal Clostridium species. Anaerobe 2013, 19, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, F.; Grander, C.; Effenberger, M.; Adolph, T.E.; Tilg, H. Gut Dysfunction and Non-alcoholic Fatty Liver Disease. Front. Endocrinol. 2019, 10, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Quevrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermúdez-Humarán, L.G.; Pigneur, B.; et al. Identification of an antiinflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut 2016, 65, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Walters, W.A.; Xu, Z.; Knight, R. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Lett. 2014, 588, 4223–4233. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef]
- Tims, S.; Derom, C.; Jonkers, D.M.; Vlietinck, R.; Saris, W.H.; Kleerebezem, M.; de Vos, W.M.; Zoetendal, E.G. Microbiota conservation and BMI signatures in adult monozygotic twins. ISME J. 2013, 7, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human genetics shape the gut microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Byndloss, M.X.; Olsan, E.E.; Rivera-Chávez, F.; Tiffany, C.R.; Cevallos, S.A.; Lokken, K.L.; Torres, T.P.; Byndloss, A.J.; Faber, F.; Gao, Y.; et al. Microbiota-activated PPAR-γ signaling inhibits dysbiotic Enterobacteriaceae expansion. Science 2017, 357, 570–575. [Google Scholar] [CrossRef]
- Sokol, H.; Seksik, P.; Rigottier-Gois, L.; Lay, C.; Lepage, P.; Podglajen, I.; Marteau, P.; Doré, J. Specificities of the fecal microbiota in inflammatory bowel disease. Inflamm. Bowel. Dis. 2006, 12, 106–111. [Google Scholar] [CrossRef]
- Samuel, B.S.; Gordon, J.I. A humanized gnotobiotic mouse model of host-archaeal-bacterial mutualism. Proc. Natl. Acad. Sci. USA 2006, 103, 10011–10016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González Hernández, M.A.; Canfora, E.E.; Jocken, J.W.E.; Blaak, E.E. The Short-Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity. Nutrients 2019, 11, E1943. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, J.; Shi, Y.; Su, W.; Chen, J.; Zhang, Z.; Wang, G.; Wang, F. Short chain fatty acid acetate protects against ethanol-induced acute gastric mucosal lesion in mice. Biol. Pharm. Bull. 2017, 40, 1439–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Q.; Li, H.; Wen, C.; He, Z. Alterations of the Gut Microbiome Associated With the Treatment of Hyperuricaemia in Male Rats. Front. Microbiol. 2018, 9, 2233. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization; WHO Global InfoBase Team. The SuRF Report 2. Surveillance of Chronic Disease Risk Factors: Country-level data and comparable estimates. Geneva, World Health Organization. 2005, p.22, Table 3.1. Available online: http://apps.who.int/iris/bitstream/handle/10665/43190/9241593024_eng.pdf?sequence=1&isAllowed=y&ua=1 (accessed on 10 November 2018).
- WHO Multicentre Growth Reference Study Group. WHO Child Growth Standards: Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for-Age: Methods and Development; World Health Organization: Geneva, Switzerland, 2006; Available online: http://www.who.int/childgrowth/standards/Technical_report.pdf?ua=1 (accessed on 10 November 2018).
- D’Annunzio, G.; Vanelli, M.; Pistorio, A.; Minuto, N.; Bergamino, L.; Iafusco, D.; Lorini, R.; Diabetes Study Group of the Italian Society for Pediatric Endocrinology and Diabetes. Insulin resistance and secretion indexes in healthy Italian children and adolescents: A multicentre study. Acta Biomed. 2009, 80, 21–28. [Google Scholar]
- Simental-Mendía, L.E.; Rodríguez-Morán, M.; Guerrero-Romero, F. The product of fasting glucose and triglycerides as surrogate for identifying insulin resistance in apparently healthy subjects. Metab. Syndr. Relat. Disord. 2008, 6, 299–304. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: ftp://greengenes.microbio.me/greengenes_release/gg_13_8_otus (accessed on 15 October 2019).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package “Vegan”. R Package Version 2.0–8. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 15 October 2019).
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [Green Version]
- Borgo, F.; Riva, A.; Benetti, A.; Casiraghi, M.C.; Bertelli, S.; Garbossa, S.; Anselmetti, S.; Scarone, S.; Pontiroli, A.E.; Morace, G.; et al. Microbiota in anorexia nervosa: The triangle between bacterial species, metabolites and psychological tests. PLoS ONE 2017, 12, e0179739. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Variable | HC | GSD | p-Value | |
|---|---|---|---|---|
| Mean ± SD | Mean ± SD | |||
| Energy Intake | ||||
| kcals | 1907 ± 603 | 2420 ± 549 | 0.0468 | * |
| Proteins | ||||
| g | 74.67 ± 22.92 | 83.06 ± 20.56 | 0.3824 | |
| % energy | 16.70 ± 3.77 | 13.80 ± 2.20 | 0.0815 | |
| Lipids | ||||
| g | 77.96 ± 47.06 | 62.35 ± 15.47 | 0.7021 | |
| % energy | 36.58 ± 10.65 | 23.40 ± 3.26 | 0.0013 | ** |
| Carbohydrates | ||||
| g | 216.19 ± 54.55 | 390.03 ± 97.78 | 0.0007 | *** |
| % energy | 46.28 ± 9.01 | 60.22 ± 4.54 | <0.0001 | **** |
| Sugars | ||||
| g | 58.56 ± 25.44 | 23.75 ± 9.11 | 0.0013 | ** |
| % energy | 11.98 ± 4.74 | 3.56 ± 1.06 | <0.0001 | **** |
| Fiber | ||||
| overall, g | 15.44 ± 4.80 | 21.01 ± 4.37 | 0.0148 | * |
| overall, g/1000 kcal | 8.58 ± 2.16 | 9.10 ± 2.72 | 0.7021 | |
| insoluble fiber, g | 6.43 ± 4.44 | 9.59 ± 4.43 | 0.1285 | |
| soluble fiber, g | 2.01 ± 1.35 | 3.57 ± 1.23 | 0.0227 | * |
| Genus | Average Relative Abundance | p-Value | |
|---|---|---|---|
| HC | GSD | ||
| Bacteroides | 18.83 | 14.43 | 0.2410 |
| Faecalibacterium | 19.61 | 7.44 | 0.0209 * |
| Unclassified Ruminococcaceae | 8.94 | 5.27 | 0.0700 |
| Ruminococcus | 7.25 | 1.42 | 0.0173 * |
| Escherichia | 0.99 | 10.01 | 0.0077 ** |
| Unclassified Clostridiales | 6.87 | 0.29 | 0.0025 ** |
| Akkermansia | 1.63 | 6.26 | 0.2323 |
| Unclassified Lachnospiraceae | 4.48 | 1.41 | 0.0428 * |
| Roseburia | 3.27 | 1.50 | 0.0428 * |
| Dialister | 3.57 | 1.64 | 0.0360 * |
| Coprococcus | 2.43 | 2.80 | 0.4138 |
| Oscillospira | 3.35 | 0.64 | 0.0020 ** |
| Unclassified Rikenellaceae | 2.35 | 1.87 | 0.1657 |
| Veillonella | 0.41 | 4.73 | 0.1265 |
| Clostridium | 0.76 | 3.94 | 0.4996 |
| Streptococcus | 0.51 | 4.16 | 0.1886 |
| Blautia | 1.40 | 1.83 | 0.4996 |
| SMB53 | 0.29 | 3.05 | 0.0360 * |
| Parabacteroides | 1.07 | 0.84 | 0.1658 |
| Phascolarctobacterium | 0.51 | 1.58 | 0.4946 |
| Megamonas | 0.00 | 5.64 | 0.0092 ** |
| Citrobacter | 0.07 | 5.33 | 0.3609 |
| Bifidobacterium | 0.23 | 0.70 | 0.7754 |
| Other genera | 10.68 | 13.41 | -- |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceccarani, C.; Bassanini, G.; Montanari, C.; Casiraghi, M.C.; Ottaviano, E.; Morace, G.; Biasucci, G.; Paci, S.; Borghi, E.; Verduci, E. Proteobacteria Overgrowth and Butyrate-Producing Taxa Depletion in the Gut Microbiota of Glycogen Storage Disease Type 1 Patients. Metabolites 2020, 10, 133. https://doi.org/10.3390/metabo10040133
Ceccarani C, Bassanini G, Montanari C, Casiraghi MC, Ottaviano E, Morace G, Biasucci G, Paci S, Borghi E, Verduci E. Proteobacteria Overgrowth and Butyrate-Producing Taxa Depletion in the Gut Microbiota of Glycogen Storage Disease Type 1 Patients. Metabolites. 2020; 10(4):133. https://doi.org/10.3390/metabo10040133
Chicago/Turabian StyleCeccarani, Camilla, Giulia Bassanini, Chiara Montanari, Maria Cristina Casiraghi, Emerenziana Ottaviano, Giulia Morace, Giacomo Biasucci, Sabrina Paci, Elisa Borghi, and Elvira Verduci. 2020. "Proteobacteria Overgrowth and Butyrate-Producing Taxa Depletion in the Gut Microbiota of Glycogen Storage Disease Type 1 Patients" Metabolites 10, no. 4: 133. https://doi.org/10.3390/metabo10040133
APA StyleCeccarani, C., Bassanini, G., Montanari, C., Casiraghi, M. C., Ottaviano, E., Morace, G., Biasucci, G., Paci, S., Borghi, E., & Verduci, E. (2020). Proteobacteria Overgrowth and Butyrate-Producing Taxa Depletion in the Gut Microbiota of Glycogen Storage Disease Type 1 Patients. Metabolites, 10(4), 133. https://doi.org/10.3390/metabo10040133