Nicotinic Acid Improves Endurance Performance of Mice Subjected to Treadmill Exercise

 , ,
, ,
Abstract
:1. Introduction
2. Results
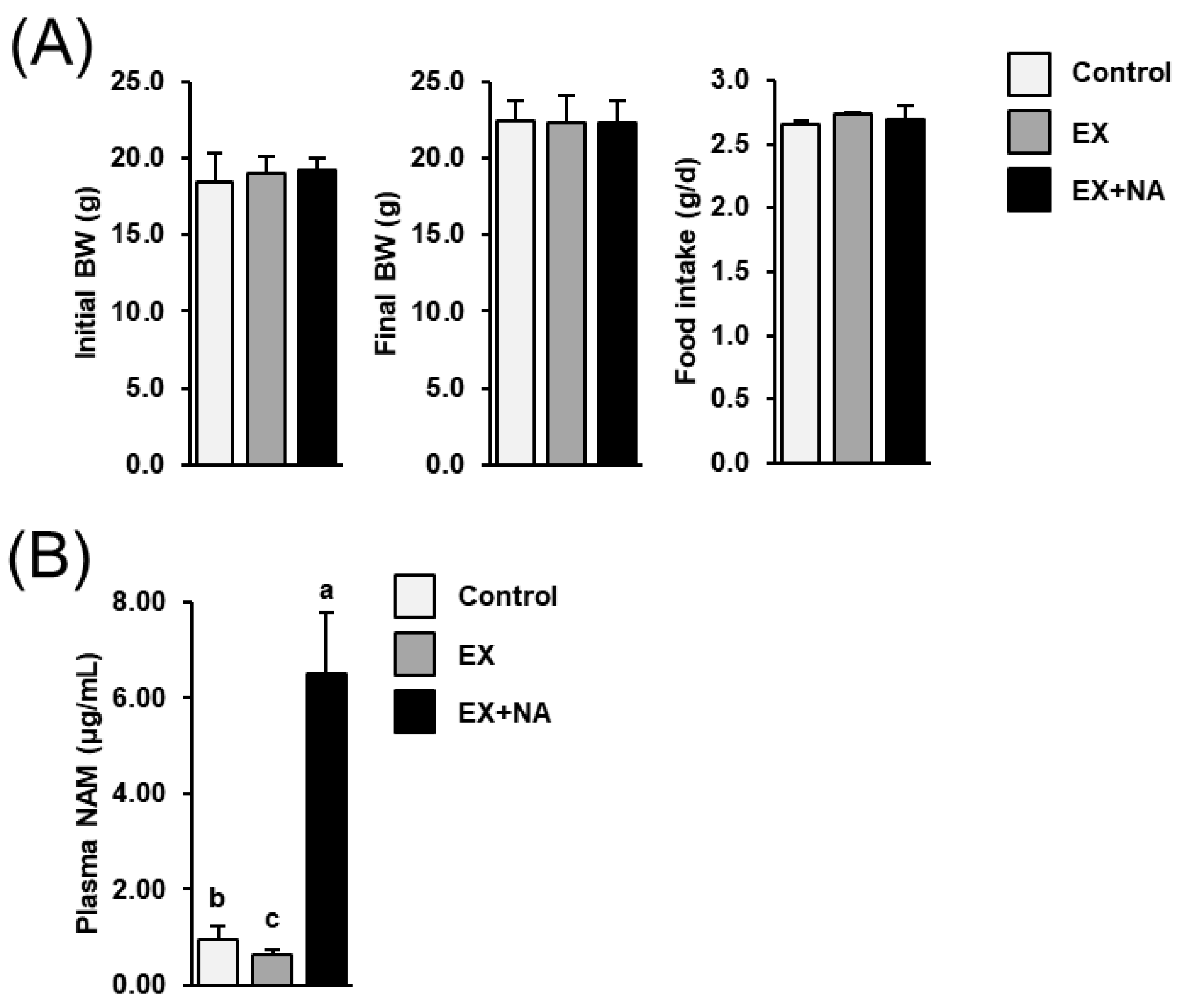
2.1. Body Weight Development and Food Intake of the Mice
2.2. Development of Endurance Exercise Capacity of the Mice
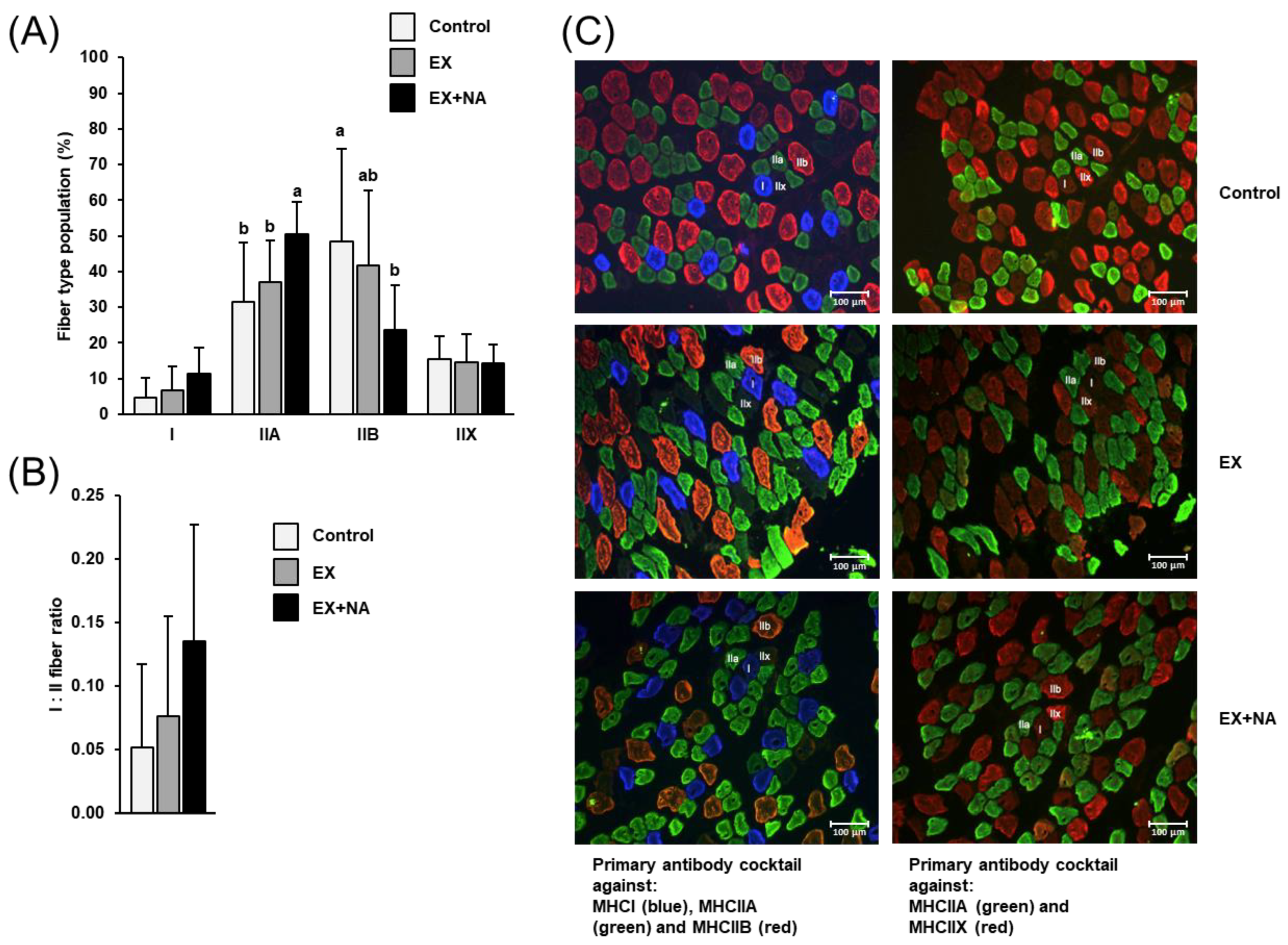
2.3. Fiber Type Composition of Gastrocnemius Muscle of the Mice
2.4. Expression of Genes involved in Mitochondrial Biogenesis and Genes Encoding Mitochondrial Enzymes in GN Muscle of the Mice
2.5. Plasma Concentrations of Metabolites Related to Fatty Acid Metabolism in the Mice
2.6. Expression of Genes Encoding Enzymes of Carbohydrate Metabolism and Concentration of Glycogen in GN Muscle of the Mice
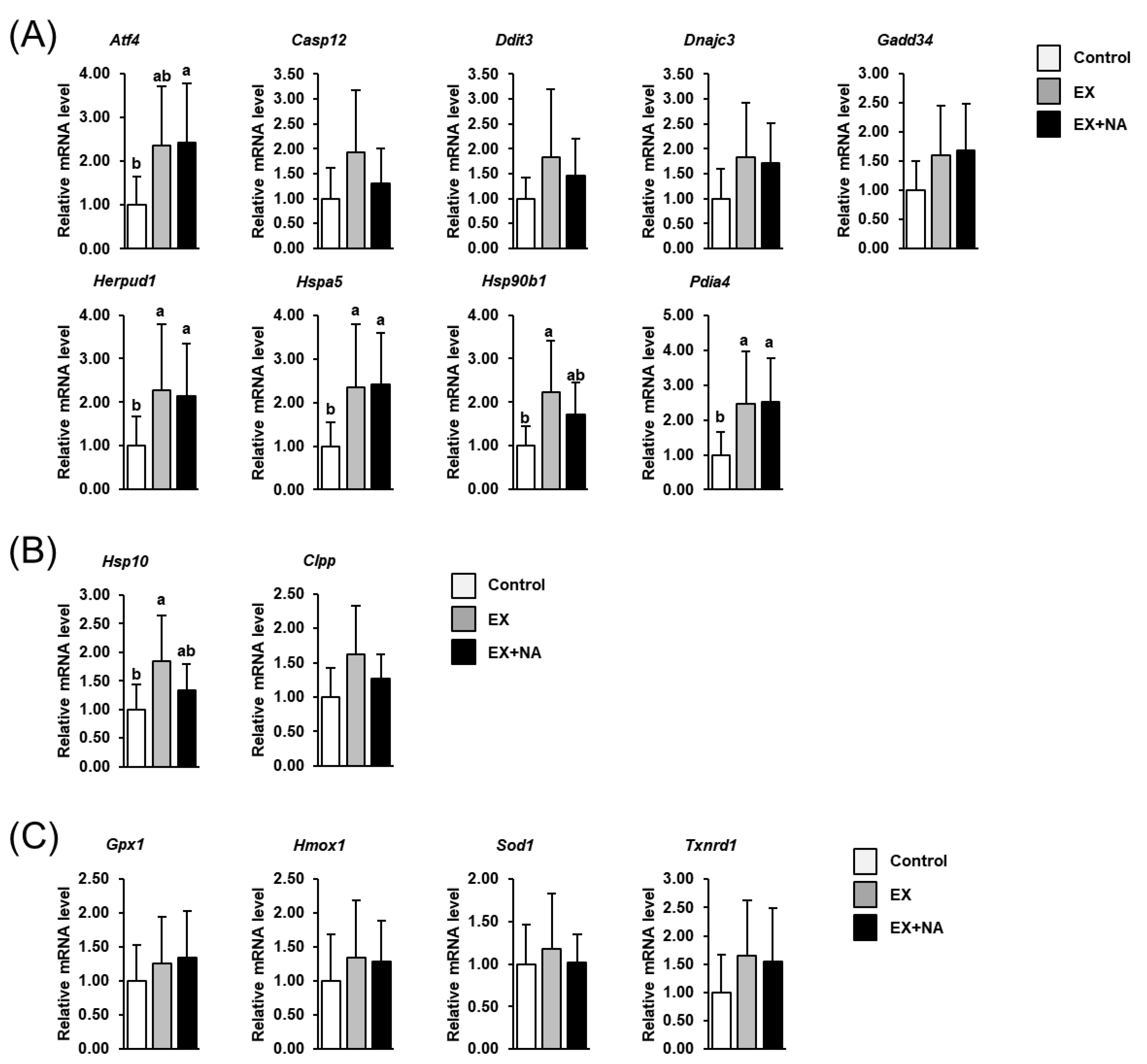
2.7. Expression of Genes Involved in the Cellular Stress Response in GN Muscle of the Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Diets
4.3. Determination of Endurance Exercise Performance
4.4. Endurance Exercise Program
4.5. Sample Collection
4.6. Immunohistochemical Muscle Fiber Typing Using Fluorescence-Labelled Antibodies
4.7. RNA Isolation and qPCR Analysis
4.8. Determination of Biochemical Parameters
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gille, A.; Bodor, E.T.; Ahmed, K.; Offermanns, S. Nicotinic acid: Pharmacological effects and mechanisms and mechanisms of action. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 79–106. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.N. The plasma free fatty acid rebound induced by nicotinic acid. J. Lipid Res. 1967, 8, 239–244. [Google Scholar] [PubMed]
- Ringseis, R.; Rosenbaum, S.; Gessner, D.K.; Herges, L.; Kubens, J.F.; Mooren, F.C.; Krüger, K.; Eder, K. Supplementing obese Zucker rats with niacin induces the transition of glycolytic to oxidative skeletal muscle fibers. J. Nutr. 2013, 143, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou Mrad, J.; Yakubu, F.; Lin, D.; Peters, J.C.; Atkinson, J.B.; Hill, J.O. Skeletal muscle composition in dietary obesity-susceptible and dietary obesity-resistant rats. Am. J. Physiol. 1992, 262, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Tanner, C.J.; Barakat, H.A.; Dohm, G.L.; Pories, W.J.; MacDonald, K.G.; Cunningham, P.R.; Swanson, M.S.; Houmard, J.A. Muscle fiber type is associated with obesity and weight loss. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 1191–1196. [Google Scholar] [CrossRef] [Green Version]
- Barnard, R.J.; Edgerton, V.R.; Furukawa, T.; Peter, J.B. Histochemical, biochemical, and contractile properties of red, white, and intermediate fibers. Am. J. Physiol. 1971, 220, 410–414. [Google Scholar] [CrossRef]
- Pette, D. Training effects on the contractile apparatus. Acta Physiol. Scand. 1998, 162, 367–376. [Google Scholar] [CrossRef]
- Wu, J.; Ruas, J.L.; Estall, J.L.; Rasbach, K.A.; Choi, J.H.; Ye, L.; Boström, P.; Tyra, H.M.; Crawford, R.W.; Campbell, K.P.; et al. The unfolded protein response mediates adaptation to exercise in skeletal muscle through a PGC-1α/ATF6α complex. Cell Metab. 2011, 13, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Mottis, A.; Jovaisaite, V.; Auwerx, J. The mitochondrial unfolded protein response in mammalian physiology. Mamm. Genome 2014, 25, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Scholz, K.; Kynast, A.M.; Couturier, A.; Mooren, F.C.; Krüger, K.; Most, E.; Eder, K.; Ringseis, R. Supplementing healthy rats with a high-niacin dose has no effect on muscle fiber distribution and muscle metabolic phenotype. Eur. J. Nutr. 2014, 53, 1229–1236. [Google Scholar] [CrossRef]
- Kirkland, J.B. Niacin status, NAD distribution and ADP-ribose metabolism. Curr. Pharm. Des. 2009, 15, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Röckl, K.S.; Witczak, C.A.; Goodyear, L.J. Signaling mechanisms in skeletal muscle: Acute responses and chronic adaptations to exercise. IUBMB Life 2008, 60, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, H.W.; Toedebusch, R.G.; Ruegsegger, G.; Mobley, C.B.; Fox, C.D.; McGinnis, G.R.; Quindry, J.C.; Booth, F.W.; Roberts, M.D.; Kavazis, A.N. Comparative adaptations in oxidative and glycolytic muscle fibers in a low voluntary wheel running rat model performing three levels of physical activity. Physiol. Rep. 2015, 3, e12619. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ren, X.; Lamb, G.D.; Murphy, R.M. Physiological and biochemical characteristics of skeletal muscles in sedentary and active rats. J. Muscle Res. Cell Motil. 2018, 39, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Southgate, R.J.; Holmes, A.G.; Febbraio, M.A. Suppression of plasma free fatty acids upregulates peroxisome proliferator-activated receptor (PPAR) α and δ and PPAR coactivator 1α in human skeletal muscle, but not lipid regulatory genes. J. Mol. Endocrinol. 2004, 33, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Ringseis, R.; Mooren, F.C.; Krüger, K.; Most, E.; Eder, K. Niacin supplementation increases the number of oxidative type I fibers in skeletal muscle of growing pigs. BMC Vet. Res. 2013, 9, 177. [Google Scholar] [CrossRef] [Green Version]
- Garvey, S.M.; Russ, D.W.; Skelding, M.B.; Dugle, J.E.; Edens, N.K. Molecular and metabolomic effects of voluntary running wheel activity on skeletal muscle in late middle-aged rats. Physiol. Rep. 2015, 3, e12319. [Google Scholar] [CrossRef]
- Hawley, J.A.; Leckey, J.J. Carbohydrate Dependence During Prolonged, Intense Endurance Exercise. Sports Med. 2015, 45, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Hearris, M.A.; Hammond, K.M.; Fell, J.M.; Morton, J.P. Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations. Nutrients 2018, 10, 298. [Google Scholar] [CrossRef] [Green Version]
- Hearris, M.A.; Hammond, K.M.; Seaborne, R.A.; Stocks, B.; Shepherd, S.O.; Philp, A.; Sharples, A.P.; Morton, J.P.; Louis, J.B. Graded reductions in preexercise muscle glycogen impair exercise capacity but do not augment skeletal muscle cell signaling: Implications for CHO periodization. J. Appl. Physiol. 2019, 126, 1587–1597. [Google Scholar] [CrossRef]
- Poynten, A.M.; Gan, S.K.; Kriketos, A.D.; O’Sullivan, A.; Kelly, J.J.; Ellis, B.A.; Chisholm, D.J.; Campbell, L.V. Nicotinic acid-induced insulin resistance is related to increased circulating fatty acids and fat oxidation but not muscle lipid content. Metabolism 2003, 52, 699–704. [Google Scholar] [CrossRef]
- Memme, J.M.; Erlich, A.T.; Hood, D.A.; Phukan, G. Exercise and mitochondrial health. J. Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.M.; Haynes, C.M. Mitochondrial protein quality control during biogenesis and aging. Trends Biochem. Sci. 2011, 36, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Quabbe, H.J.; Luyckx, A.S.; L’age, M.; Schwarz, C. Growth hormone, cortisol, and glucagon concentrations during plasma free fatty acid depression: Different effects of nicotinic acid and an adenosine derivative (BM 11.189). J. Clin. Endocrinol. Metab. 1983, 57, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Westphal, S.; Borucki, K.; Taneva, E.; Makarova, R.; Luley, C. Extended-release niacin raises adiponectin and leptin. Atherosclerosis 2007, 193, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Houtkooper, R.H.; Cantó, C.; Wanders, R.J.; Auwerx, J. The secret of life of NAD+: An old metabolite controlling new metabolic signaling pathways. Endocr. Rev. 2010, 31, 194–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundby, C.; Montero, D.; Joyner, M. Biology of VO(2) max: Looking under the physiology lamp. Acta Physiol. 2017, 220, 218–228. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Laboratory Animals; National Academy Press: Washington, DC, USA, 1995. [Google Scholar]
- Kaup, D.; Keller, J.; Most, E.; Geyer, J.; Eder, K.; Ringseis, R. The carnitine status does not affect the contractile and metabolic phenotype of skeletal muscle in pigs. Nutr. Metab. 2018, 15, 2. [Google Scholar] [CrossRef] [Green Version]
- Keller, J.; Ringseis, R.; Koc, A.; Lukas, I.; Kluge, H.; Eder, K. Supplementation with L-carnitine downregulates genes of the ubiquitin proteasome system in the skeletal muscle and liver of piglets. Animal 2012, 6, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Hirche, F.; Fischer, M.; Keller, J.; Eder, K. Determination of carnitine, its short chain acyl esters and metabolic precursors trimethyllysine and gamm-butyrobetaine by quasi-solid phase extraction and MS/MS detection. J. Chromatogr. B 2009, 877, 2158–2162. [Google Scholar] [CrossRef]
- Dawson, K.D.; Baker, D.J.; Greenhaff, P.L.; Gibala, M.J. An acute decrease in TCA cycle intermediates does not affect aerobic energy delivery in contracting rat skeletal muscle. J. Physiol. 2005, 565, 637–643. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adequate NA Diet (g/kg) (30 mg niacin/kg diet) | High NA Diet (g/kg) (780 mg niacin/kg diet) | |
|---|---|---|
| Maize starch | 532 | 532 |
| Casein | 200 | 200 |
| Saccharose | 100 | 100 |
| Soybean oil | 70 | 70 |
| Cellulose | 50 | 49.25 |
| Mineral mix 1 | 35 | 35 |
| Vitamin mix 2 | 10 | 10 |
| L-Cystein | 3 | 3 |
| Supplemental NA | - | 0.75 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ringseis, R.; Gessner, D.K.; Beer, A.M.; Albrecht, Y.; Wen, G.; Most, E.; Krüger, K.; Eder, K. Nicotinic Acid Improves Endurance Performance of Mice Subjected to Treadmill Exercise. Metabolites 2020, 10, 138. https://doi.org/10.3390/metabo10040138
Ringseis R, Gessner DK, Beer AM, Albrecht Y, Wen G, Most E, Krüger K, Eder K. Nicotinic Acid Improves Endurance Performance of Mice Subjected to Treadmill Exercise. Metabolites. 2020; 10(4):138. https://doi.org/10.3390/metabo10040138
Chicago/Turabian StyleRingseis, Robert, Denise K. Gessner, Anna M. Beer, Yvonne Albrecht, Gaiping Wen, Erika Most, Karsten Krüger, and Klaus Eder. 2020. "Nicotinic Acid Improves Endurance Performance of Mice Subjected to Treadmill Exercise" Metabolites 10, no. 4: 138. https://doi.org/10.3390/metabo10040138
APA StyleRingseis, R., Gessner, D. K., Beer, A. M., Albrecht, Y., Wen, G., Most, E., Krüger, K., & Eder, K. (2020). Nicotinic Acid Improves Endurance Performance of Mice Subjected to Treadmill Exercise. Metabolites, 10(4), 138. https://doi.org/10.3390/metabo10040138