
Evidence That Agouti-Related Peptide May Directly Regulate Kisspeptin Neurons in Male Sheep

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
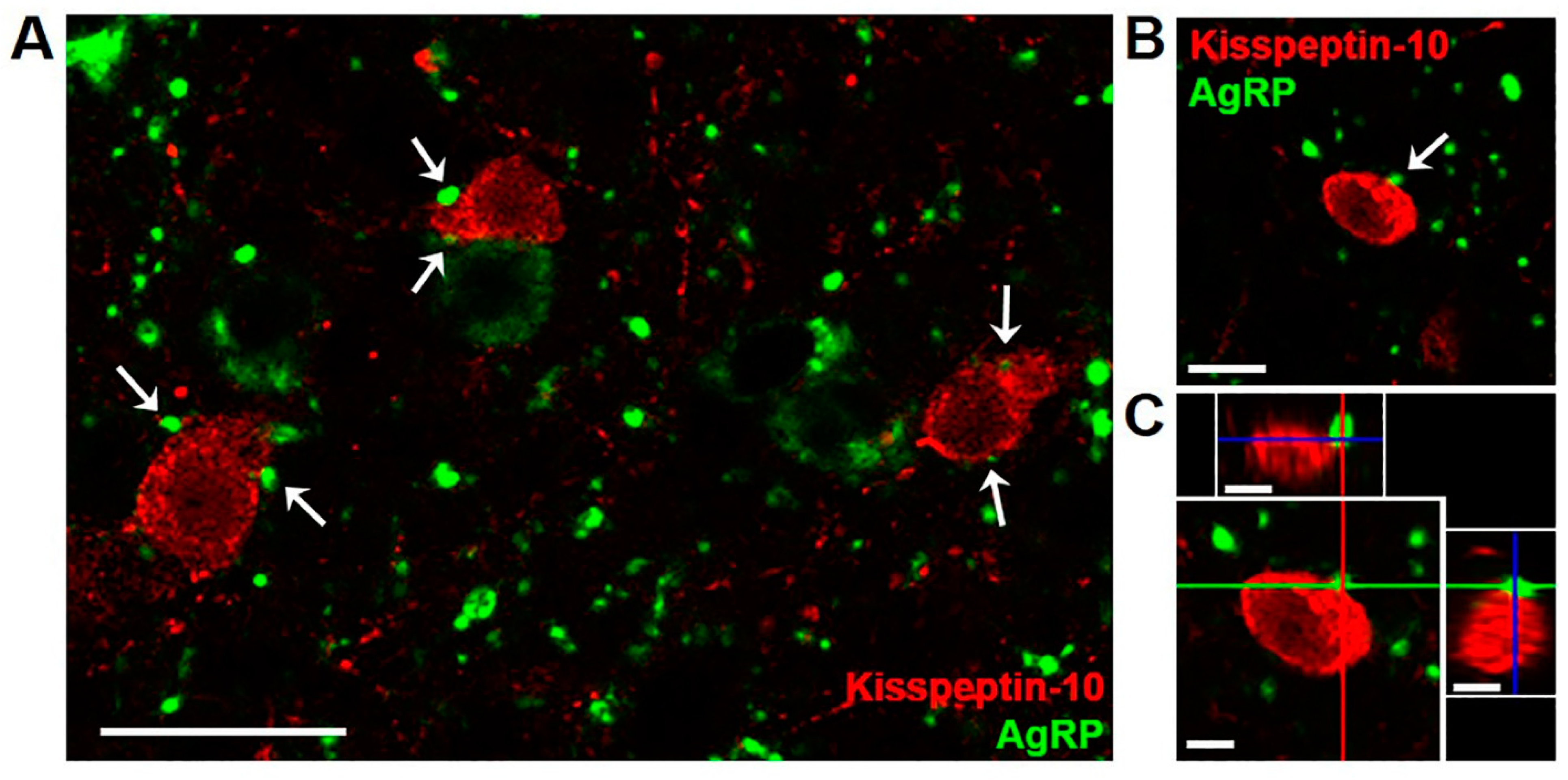
2.1. AgRP Inputs to Kisspeptin Cells
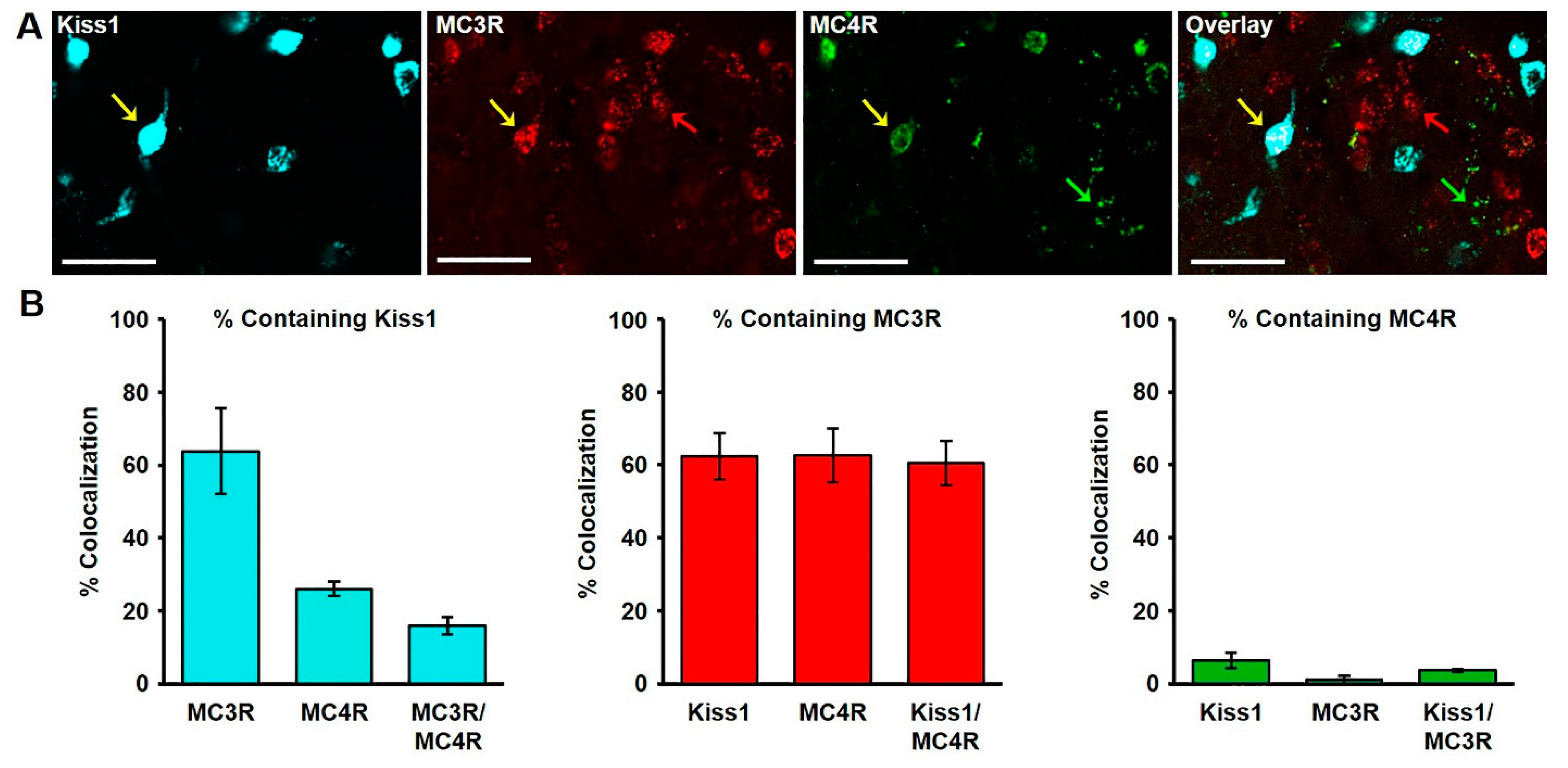
2.2. Colocalization of Kisspeptin, MC3R, and MC4R
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Tissue Collection
4.3. Dual-Label Immunofluorescent Detection of Kisspeptin and AgRP
4.4. RNAscope In Situ Hybridization for Kisspeptin and Melanocortin Receptors (MC3R, MC4R)
4.5. Confocal Analyses
4.5.1. Immunofluorescence
4.5.2. RNAscope
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ebling, F.; Wood, R.; Karsch, F.; Vannerson, L.; Suttie, J.; Bucholtz, D.; Schall, R.; Foster, D. Metabolic interfaces between growth and reproduction. Iii. Central mechanisms controlling pulsatile luteinizing hormone secretion in the nutritionally growth-limited female lamb. Endocrinology 1990, 126, 2719–2727. [Google Scholar] [CrossRef]
- Prasad, B.; Conover, C.; Sarkar, D.; Rabii, J.; Advis, J. Feed restriction in prepubertal lambs: Effect on puberty onset and on in vivo release of luteinizing-hormone-releasing hormonE.; neuropeptide y and beta-endorphin from the posterior-lateral median eminence. Neuroendocrinology 1993, 57, 1171–1181. [Google Scholar] [CrossRef]
- I’anson, H.; Manning, J.; Herbos, C.; Pelt, J.; Friedman, C.; Wood, R.; Bucholtz, D.; Foster, D. Central inhibition of gonadotropin-releasing hormone secretion in the growth-restricted hypogonadotropic female sheep. Endocrinology 2000, 141, 520–527. [Google Scholar] [CrossRef]
- Finn, P.; Cunningham, M.; Pau, K.; Spies, H.; Clifton, D.; Steiner, R. The stimulatory effect of leptin on the neuroendocrine reproductive axis of the monkey. Endocrinology 1998, 139, 4652–4662. [Google Scholar] [CrossRef]
- Hakansson, M.; Brown, H.; Ghilardi, N.; Skoda, R.; Meister, B. Leptin receptor immunoreactivity in chemically defined target neurons of the hypothalamus. J. Neurosci. 1998, 18, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Quennell, J.; Mulligan, A.; Tups, A.; Liu, X.; Phipps, S.; Kemp, C.; Herbison, A.; Grattan, D.; Anderson, G. Leptin indirectly regulates gonadotropin-releasing hormone neuronal function. Endocrinology 2009, 150, 2805–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottsch, M.; Cunningham, M.; Smith, J.; Popa, S.; Acohido, B.; Crowley, W.; Seminara, S.; Clifton, D.; Steiner, R. A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology 2004, 145, 4073–4077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschini, I.; Lomet, D.; Cateau, M.; Delsol, G.; Tillet, Y.; Caraty, A. Kisspeptin immunoreactive cells of the ovine preoptic area and arcuate nucleus co-express estrogen receptor alpha. Neurosci. Lett. 2006, 401, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Lehman, M.; Merkley, C.; Coolen, L.; Goodman, R. Anatomy of the kisspeptin neural network in mammals. Brain Res. 2010, 1364, 90–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbara, A.; Ratnasabapathy, R.; Jayasena, C.; Dhillo, W. The effects of kisspeptin on gonadotropin release in non-human mammals. Adv. Exp. Med. Biol. 2013, 784, 63–87. [Google Scholar] [PubMed]
- Irwig, M.; Fraley, G.; Smith, J.; Acohido, B.; Popa, S.; Cunningham, M.; Gottsch, M.; Clifton, D.; Steiner, R. Kisspeptin activation of gonadotropin releasing hormone neurons and regulation of kiss-1 mrna in the male raT. Neuroendocrinology 2004, 80, 264–272. [Google Scholar] [CrossRef]
- Herbison, A.; De Tassigny, X.; Doran, J.; Colledge, W. Distribution and postnatal development of gpr54 gene expression in mouse brain and gonadotropin-releasing hormone neurons. Endocrinology 2010, 151, 312–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.; Li, Q.; Yap, K.; Shahab, M.; Roseweir, A.; Millar, R.; Clarke, I. Kisspeptin is essential for the full preovulatory lh surge and stimulates gnrh release from the isolated ovine median eminence. Endocrinology 2011, 152, 1001–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, M.; Tonsfeldt, K.; Ronnekleiv, O. Mrna expression of ion channels in gnrh neurons: Subtype-specific regulation by 17beta-estradiol. Mol. Cell Endocrinol. 2013, 367, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestor, C.; Bedenbaugh, M.; Hileman, S.; Coolen, L.; Lehman, M.; Goodman, R. Regulation gnrh pulsatility in ewes. Reproduction 2018, 156, R83–R99. [Google Scholar] [CrossRef]
- Lehman, M.; Coolen, L.; Goodman, R. Importance of neuroanatomical data from domestic animals to the development and testing of the kndy hypothesis for gnrh pulse generation. Domest. Anim. Endocrinol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Merkley, C.; Coolen, L.; Goodman, R.; Lehman, M. Evidence for changes in numbers of synaptic inputs onto kndy and gnrh neurones during the preovulatory lh surge in the ewe. J. Neuroendocrinol. 2015, 27, 624–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, J.; Navarro, V.; Fernandez-Fernandez, R.; Nogueiras, R.; Tovar, S.; Roa, J.; Vazquez, M.; Vigo, E.; Casanueva, F.; Aguilar, E.; et al. Changes in hypothalamic kiss-1 system and restoration of pubertal activation of the reproductive axis by kisspeptin in undernutrition. Endocrinology 2005, 146, 3917–3925. [Google Scholar] [CrossRef] [Green Version]
- True, C.; Kirigiti, M.; Kievit, P.; Grove, K.; Smith, M. Leptin is not the critical signal for kisspeptin or luteinising hormone restoration during exit from negative energy balance. J. Neuroendocrinol. 2011, 23, 1099–1112. [Google Scholar] [CrossRef] [Green Version]
- Navarro, V.; Ruiz-Pino, F.; Sanchez-Garrido, M.; Garcia-Galiano, D.; Hobbs, S.; Manfredi-Lozano, M.; Leon, S.; Sangiao-Alvarellos, S.; Castellano, J.; Clifton, D.; et al. Role of neurokinin b in the control of female puberty and its modulation by metabolic status. J. Neurosci. 2012, 32, 2388–2397. [Google Scholar] [CrossRef] [Green Version]
- Polkowska, J.; Cieslak, M.; Wankowska, M.; Wojcik-Gladysz, A. The effect of short fasting on the hypothalamic neuronal system of kisspeptin in peripubertal female lambs. Anim. Reprod. Sci. 2015, 159, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Merkley, C.; Renwick, A.; Shuping, S.; Harlow, K.; Sommer, J.; Nestor, C. Undernutrition reduces kisspeptin and neurokinin b expression in castrated male sheep. Reprod. Fertil. 2020, 1, 21–33. [Google Scholar] [CrossRef]
- Smith, J.; Acohido, B.; Clifton, D.; Steiner, R. Kiss-1 neurones are direct targets for leptin in the ob/ob mouse. J. Neuroendocr. 2006, 18, 298–303. [Google Scholar] [CrossRef]
- Backholer, K.; Smith, J.; Rao, A.; Pereira, A.; Iqbal, J.; Ogawa, S.; Li, Q.; Clarke, I. Kisspeptin cells in the ewe brain respond to leptin and communicate with neuropeptide y and proopiomelanocortin cells. Endocrinology 2010, 151, 2233–2243. [Google Scholar] [CrossRef] [PubMed]
- Cravo, R.; Margatho, L.; Osborne-Lawrence, S.; Donato, J.; Atkin, S.; Bookout, A.; Rovinsky, S.; Frazao, R.; Lee, E.; Gautron, L.; et al. Characterization of kiss1 neurons using transgenic mouse models. Neuroscience 2011, 173, 37–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, G.; Greenwald-Yarnell, M.; Phillips, R.; Coolen, L.; Lehman, M.; Myers, M. Molecular mapping of the neural pathways linking leptin to the neuroendocrine reproductive axis. Endocrinology 2011, 152, 2302–2310. [Google Scholar] [CrossRef] [Green Version]
- Donato, J.; Cravo, R.; Frazao, R.; Gautron, L.; Scott, M.; Lachey, J.; Castro, I.; Margatho, L.; Lee, S.; Lee, C.; et al. Leptin’s effect on puberty in mice is relayed by the ventral premammillary nucleus and does not require signaling in kiss1 neurons. J. Clin. Invest. 2011, 121, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Egan, O.; Inglis, M.; Anderson, G. Leptin signaling in agrp neurons modulates puberty onset and adult fertility in mice. J. Neurosci. 2017, 37, 3875–3886. [Google Scholar] [CrossRef]
- Vulliemoz, N.; Xiao, E.; Xia-Zhang, L.; Wardlaw, S.; Ferin, M. Central infusion of agouti-related peptide suppresses pulsatile luteinizing hormone release in the ovariectomized rhesus monkey. Endocrinology 2005, 146, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Adam, C.; Archer, Z.; Findlay, P.; Thomas, L.; Marie, M. Hypothalamic gene expression in sheep for cocaine- and amphetamine-regulated transcript, pro-opiomelanocortin, neuropeptide y, agouti-related peptide and leptin receptor and responses to negative energy balance. Neuroendocrinology 2002, 75, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Mcmahon, C.; Marks, D.; Daniel, J.; Steele, B.; Sartin, J. A role for agouti-related protein in appetite regulation in a species with continuous nutrient delivery. Neuroendocrinology 2004, 80, 210–218. [Google Scholar] [CrossRef]
- Hahn, T.; Breininger, J.; Baskin, D.; Schwartz, M. Coexpression of agrp and npy in fasting-activated hypothalamic neurons. Nat. Neurosci. 1998, 1, 271–272. [Google Scholar] [CrossRef]
- Chaillou, E.; Baumont, R.; Chilliard, Y.; Tillet, T. Several subpopulations of neuropeptide y-containing neurons exist in the infundibular nucleus of sheep: An immunohistochemical study of animals on different diets. J. Comp. Neurol. 2002, 444, 129–143. [Google Scholar] [CrossRef]
- Büch, T.; Heling, D.; Damm, E.; Gudermann, T.; Breit, A. Pertussis toxin-sensitive signaling of melanocortin-4 receptors in hypothalamic gt1–7 cells defines agouti-related protein as a biased agonisT. J. Biol. Chem. 2009, 284, 26411–26420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Tao, Y. Biased signaling initiated by agouti-related peptide through human melanocortin-3 and-4 receptors. Biochim. Biophys. Acta 2016, 1862, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Merkley, C.; Shuping, S.; Nestor, C. Neuronal networks that regulate gonadotropin-releasing hormone/luteinizing hormone secretion during undernutrition: Evidence from sheep. Domest. Anim. Endocrinol. 2020. [Google Scholar] [CrossRef]
- Goodman, R.; Lehman, M.; Smith, J.; Coolen, L.; De Oliveira, C.; Jafarzadehshirazi, M.; Pereira, A.; Iqbal, J.; Caraty, A.; Ciofi, P.; et al. Kisspeptin neurons in the arcuate nucleus of the ewe express both dynorphin a and neurokinin b. Endocrinology 2007, 148, 5752–5760. [Google Scholar] [CrossRef] [PubMed]
- Broberger, C.; Johansen, J.; Johnasson, C.; Schalling, M.; Hokfelt, T. The neuropeptide y/agouti gene-related protein (agrp) brain circuitry in normal, anorectiC.; and monosodium glutamate-treated mice. Proc. Natl. Acad. Sci. USA 1998, 95, 15043–15048. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, K.; Padmanabhan, V.; Coolen, L.; Lehman, M. Prenatal programming by testosterone of hypothalamic metabolic control neurones in the ewe. J. Neuroendocrinol. 2011, 23, 401–411. [Google Scholar] [CrossRef] [Green Version]
- Padilla, S.; Qiu, J.; Nestor, C.; Zhang, C.; Smith, A.; Whiddon, B.; Ronnekleiv, O.; Kelly, M.; Palmiter, R. Agrp to kiss1 neuron signaling links nutritional state and fertility. Proc. Natl. Acad. Sci. USA 2017, 114, 2413–2418. [Google Scholar] [CrossRef] [Green Version]
- Mercer, R.; Chee, M.; Colmers, W. The role of npy in hypothalamic mediated food intake. Front. Neuroendocrinol. 2011, 32, 398–415. [Google Scholar] [CrossRef]
- Anderson, E.; Cakir, I.; Carrington, S.; Cone, R.; Ghamari-Langroudi, M.; Gillyard, T.; Gimenez, L.; Litt, M. 60 years of pomc: Regulation of feeding and energy homeostasis by alpha-msh. J. Mol. Endocrinol. 2016, 56, t157–t174. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S.; Small, C.; Kim, M.; Heath, M.; Seal, L.; Russell, S.; Ghatei, M.; Bloom, S. Agouti related peptide (agrp) stimulates the hypothalamo pituitary gonadal axis in vivo and in vitro in male rats. Endocrinology 1999, 140, 5459–5462. [Google Scholar] [CrossRef]
- Israel, D.; Sheffer-Babila, S.; De Luca, C.; Jo, Y.; Liu, S.; Xia, G.; Spergel, D.; Dun, S.; Dun, N.; Chua, S.J. Effects of leptin and melanocortin signaling interactions on pubertal development and reproduction. Endocrinology 2012, 153, 2408–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roa, J.; Herbison, A. Direct regulation of gnrh neuron excitability by arcuate nucleus pomc and npy neuron neuropeptides in female mice. Endocrinology 2012, 153, 5587–5599. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-K.; Thompson, D.A.; Dickinson, C.J.; Wilken, J.; Barsh, G.S.; Kent, S.B.; Gantz, I. Characterization of agouti-related protein binding to melanocortin receptors. Mol. Endocrinol. 1999, 13, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Ollmann, M.M.; Wilson, B.D.; Yang, Y.-K.; Kerns, J.A.; Chen, Y.; Gantz, I.; Barsh, G.S. Antagonism of Central Melanocortin Receptors in Vitro and in Vivo by Agouti-Related Protein. Sci. 1997, 278, 135–138. [Google Scholar] [CrossRef]
- Backholer, K.; Smith, J.; Clarke, I. Melanocortins may stimulate reproduction by activating orexin neurons in the dorsomedial hypothalamus and kisspeptin neurons in the preoptic area of the ewe. Endocrinology 2009, 150, 5488–5497. [Google Scholar] [CrossRef] [Green Version]
- Manfredi-Lozano, M.; Roa, J.; Ruiz-Pino, F.; Piet, R.; Garcia-Galiano, D.; Pineda, R.; Zamora, A.; Leon, S.; Sanchez-Garrido, M.; Romero-Ruiz, A.; et al. Defining a novel leptin-melanocortin-kisspeptin pathway involved in the metabolic control of puberty. Mol. Metab. 2016, 5, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Sarvestani, F.; Tamadon, A.; Hematzadeh, A.; Jahanara, M.; Shirazi, M.; Moghadam, A.; Niazi, A.; Moghiminasr, R. Expression of melanocortin-4 receptor and agouti-related peptide mrnas in arcuate nucleus during long term malnutrition of female ovariectomized rats. Iran. J. Basic Med. Sci. 2015, 18, 104–107. [Google Scholar]
- Mercer, A.; Stuart, R.; Attard, C.; Otero-Corchon, V.; Nillni, E.; Low, M. Temporal changes in nutritional state affect hypothalamic pomc peptide levels independently of leptin in adult male mice. Am. J. Physiol. Endocrinol. Metab. 2014, 306, e904–e915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foradori, C.; Amstalden, M.; Goodman, R.; Lehman, M. Colocalisation of dynorphin a and neurokinin b immunoreactivity in the arcuate nucleus and median eminence of the sheep. J. Neuroendocrinol. 2006, 18, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Merkley, C.; Porter, K.; Coolen, L.; Hileman, S.; Billings, H.; Drews, S.; Goodman, R.; Lehman, M. Kndy (kisspeptin/neurokinin b/dynorphin) neurons are activated during both pulsatile and surge secretion of lh in the ewe. Endocrinology 2012, 153, 5406–5414. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merkley, C.M.; Shuping, S.L.; Sommer, J.R.; Nestor, C.C. Evidence That Agouti-Related Peptide May Directly Regulate Kisspeptin Neurons in Male Sheep. Metabolites 2021, 11, 138. https://doi.org/10.3390/metabo11030138
Merkley CM, Shuping SL, Sommer JR, Nestor CC. Evidence That Agouti-Related Peptide May Directly Regulate Kisspeptin Neurons in Male Sheep. Metabolites. 2021; 11(3):138. https://doi.org/10.3390/metabo11030138
Chicago/Turabian StyleMerkley, Christina M., Sydney L. Shuping, Jeffrey R. Sommer, and Casey C Nestor. 2021. "Evidence That Agouti-Related Peptide May Directly Regulate Kisspeptin Neurons in Male Sheep" Metabolites 11, no. 3: 138. https://doi.org/10.3390/metabo11030138
APA StyleMerkley, C. M., Shuping, S. L., Sommer, J. R., & Nestor, C. C. (2021). Evidence That Agouti-Related Peptide May Directly Regulate Kisspeptin Neurons in Male Sheep. Metabolites, 11(3), 138. https://doi.org/10.3390/metabo11030138