
Alterations in Pattern Baldness According to Sex: Hair Metabolomics Approach


Abstract
:1. Introduction
2. Results
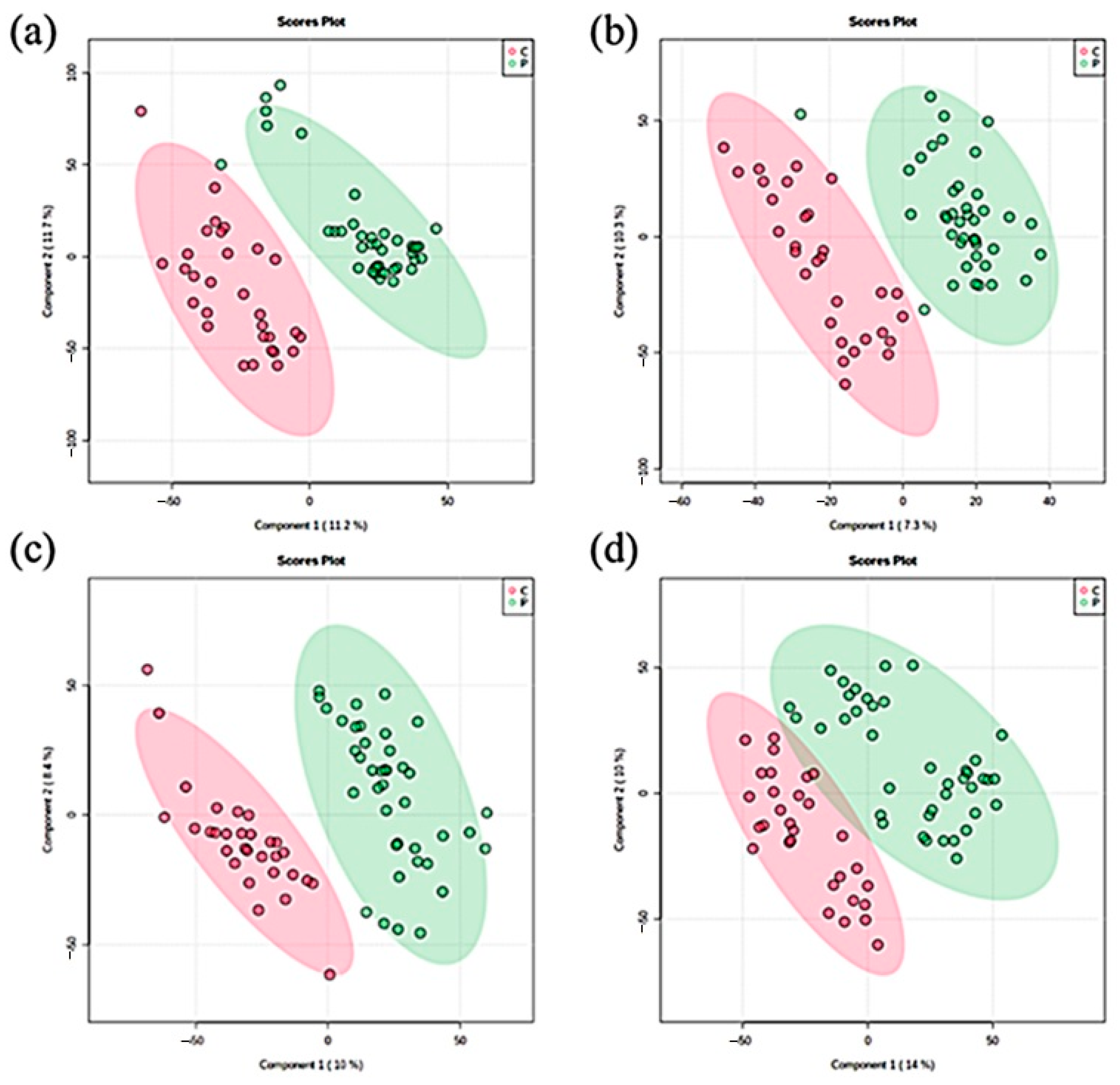
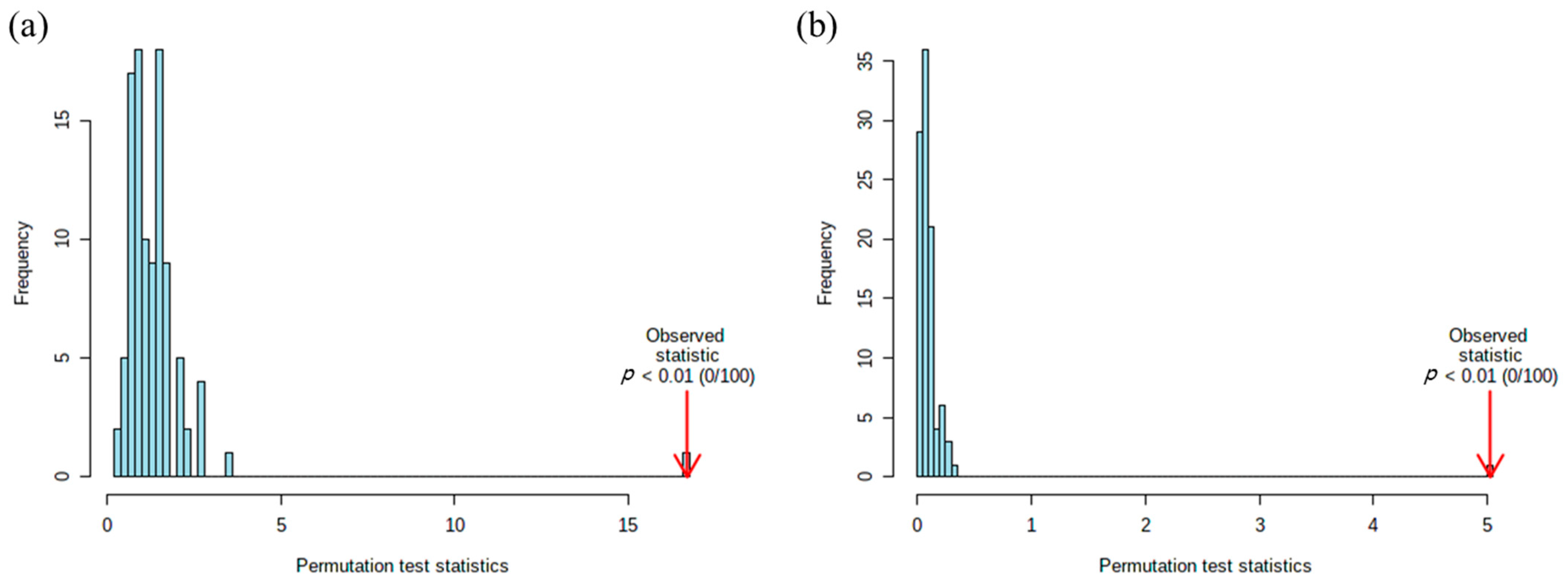
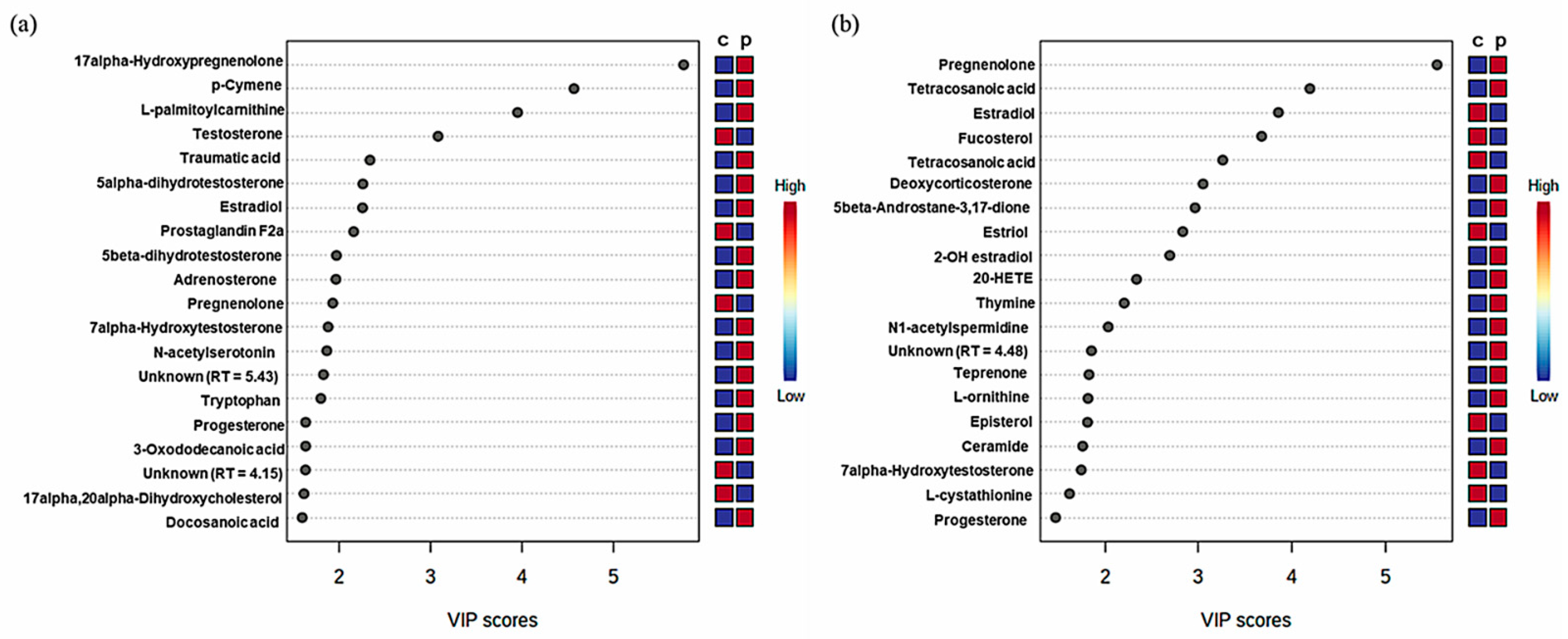
2.1. Metabolic Patterns Detected in Hair Samples of Pattern Baldness Patients
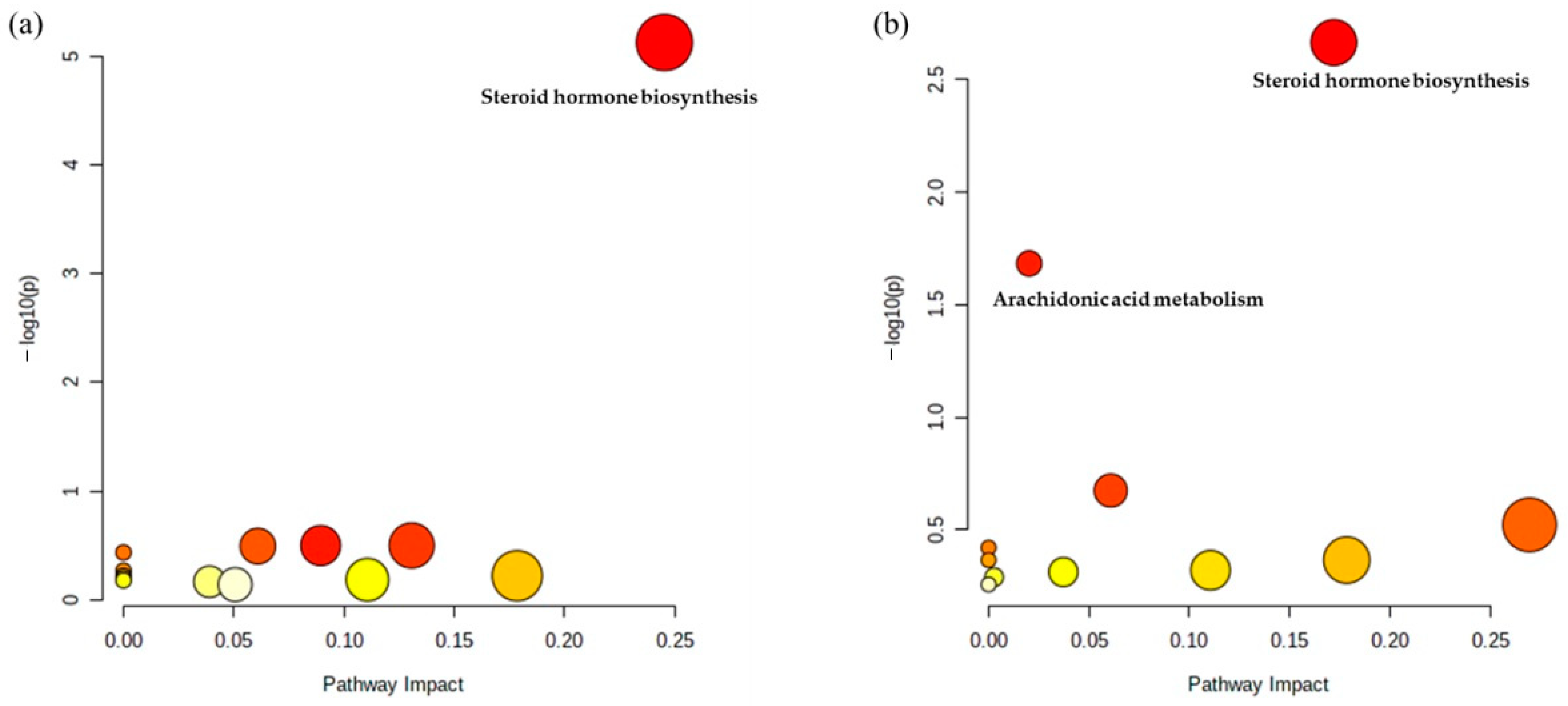
2.2. Metabolic Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Hair Sample Collection
4.3. Chemicals and Materials
4.4. Sample Preparation
4.5. Experimental Conditions
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Orentreich, N. Pathogenesis of alopecia. J. Soc. Cosmet. Chem. 1960, 11, 479–499. [Google Scholar]
- Adachi, K.; Takayasu, S.; Takashima, I.; Kano, M.; Kondo, S. Human hair follicles: Metabolism and control mechanisms. J. Soc. Cosmet. Chem. 1970, 21, 901–924. [Google Scholar]
- Ebling, F.J. The biology of hair. Dermatol. Clin. 1987, 5, 467–481. [Google Scholar] [CrossRef]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweikert, H.U.; Wilson, J.D. Regulation of human hair growth by steroid hormones. I. Testerone metabolism in isolated hairs. J. Clin. Endocrinol. Metab. 1974, 38, 811–819. [Google Scholar] [CrossRef]
- Chew, E.G.; Tan, J.H.; Bahta, A.W.; Ho, B.S.-Y.; Liu, X.; Lim, T.C.; Sia, Y.Y.; Bigliardi, P.L.; Heilmann, S.; Wan, A.C. Differential expression between human dermal papilla cells from balding and non-balding scalps reveals new candidate genes for androgenetic alopecia. J. Investig. Dermatol. 2016, 136, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Higgins, C.A.; Christiano, A.M. Regenerative medicine and hair loss: How hair follicle culture has advanced our understanding of treatment options for androgenetic alopecia. Regen. Med. 2014, 9, 101–111. [Google Scholar] [CrossRef]
- Yip, L.; Rufaut, N.; Sinclair, R. Role of genetics and sex steroid hormones in male androgenetic alopecia and female pattern hair loss: An update of what we now know. Australas. J. Dermatol. 2011, 52, 81–88. [Google Scholar] [CrossRef]
- Choi, M.H.; Yoo, Y.S.; Chung, B.C. Biochemical roles of testosterone and epitestosterone to 5 alpha-reductase as indicators of male-pattern baldness. J. Investig. Dermatol. 2001, 116, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Olsen, E.A. Androgenetic alopecia. In Disorders of Hair Growth: Diagnosis and Treatment; McGraw-Hill Professional: New York, NY, USA, 1994. [Google Scholar]
- Patel, M.; Perez, A.; Sinclair, R. An overview of female pattern hair loss. Expert Rev. Dermatol. 2011, 6, 317–327. [Google Scholar] [CrossRef]
- Olsen, E.A. Female pattern hair loss. J. Am. Acad. Dermatol. 2001, 45, S70–S80. [Google Scholar] [CrossRef]
- Birch, M.P.; Lalla, S.C.; Messenger, A.G. Female pattern hair loss. Clin. Exp. Dermatol. 2002, 27, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Futterweit, W.; Dunaif, A.; Yeh, H.C.; Kingsley, P. The prevalence of hyperandrogenism in 109 consecutive female patients with diffuse alopecia. J. Am. Acad. Dermatol. 1988, 19, 831–836. [Google Scholar] [CrossRef]
- Miller, J.A.; Darley, C.R.; Karkavitsas, K.; Kirby, J.D.; Munro, D.D. Low sex-hormone binding globulin levels in young women with diffuse hair loss. Br. J. Dermatol. 1982, 106, 331–336. [Google Scholar] [CrossRef]
- Montalto, J.; Whorwood, C.B.; Funder, J.W.; Yong, A.B.; Callan, A.; Davies, H.E.; Connelly, J.F. Plasma C19 steroid sulphate levels and indices of androgen bioavailability in female pattern androgenic alopecia. Clin. Endocrinol. 1990, 32, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Legro, R.S.; Carmina, E.; Stanczyk, F.Z.; Gentzschein, E.; Lobo, R.A. Alterations in androgen conjugate levels in women and men with alopecia. Fertil. Steril. 1994, 62, 744–750. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metab. 2013, 1, 92–107. [Google Scholar] [CrossRef]
- Sugimoto, M.; Kawakami, M.; Robert, M.; Soga, T.; Tomita, M. Bioinformatics Tools for Mass Spectroscopy-Based Metabolomic Data Processing and Analysis. Curr. Bioinform. 2012, 7, 96–108. [Google Scholar] [CrossRef]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass. Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T. Hair and nails: Advantages and pitfalls when used in biological monitoring. In Biological Monitoring of Toxic Metals; Springer: Berlin/Heidelberg, Germany, 1988; pp. 623–640. [Google Scholar]
- Pozebon, D.; Scheffler, G.L.; Dressler, V.L. Elemental hair analysis: A review of procedures and applications. Anal. Chim. Acta 2017, 992, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Jurado, C. Forensic applications of hair analysis. In Hair Analysis in Clinical and Forensic Toxicology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 241–273. [Google Scholar]
- Bang, H.J.; Yang, Y.J.; Lho, D.S.; Lee, W.Y.; Sim, W.Y.; Chung, B.C. Comparative studies on level of androgens in hair and plasma with premature male-pattern baldness. J. Dermatol. Sci. 2004, 34, 11–16. [Google Scholar] [CrossRef]
- Ryu, H.K.; Kim, K.M.; Yoo, E.A.; Sim, W.Y.; Chung, B.C. Evaluation of androgens in the scalp hair and plasma of patients with male-pattern baldness before and after finasteride administration. Br. J. Dermatol. 2006, 154, 730–734. [Google Scholar] [CrossRef]
- Ryu, H.K.; Jung, B.H.; Kim, K.M.; Yoo, E.A.; Woo, J.T.; Chung, B.C. Determination of cholesterol in human hair using gas chromatography-mass spectrometry. Biomed. Chromatogr. 2006, 20, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Binz, T.M.; Braun, U.; Baumgartner, M.R.; Kraemer, T. Development of an LC-MS/MS method for the determination of endogenous cortisol in hair using (13)C3-labeled cortisol as surrogate analyte. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1033–1034, 65–72. [Google Scholar] [CrossRef]
- Barberini, L.; Noto, A.; Saba, L.; Palmas, F.; Fanos, V.; Dessi, A.; Zavattoni, M.; Fattuoni, C.; Mussap, M. Multivariate data validation for investigating primary HCMV infection in pregnancy. Data Brief 2016, 9, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.H.; Kim, S.J.; Lew, B.L.; Sim, W.Y.; Chung, B.C. Hair steroid profiling reveals racial differences in male pattern baldness between Korean and Caucasian populations. J. Investig. Dermatol. 2013, 133, 822–824. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, K.D. Androgens and alopecia. Mol. Cell. Endocrinol. 2002, 198, 89–95. [Google Scholar] [CrossRef]
- Wallace, M.L.; Smoller, B.R. Estrogen and progesterone receptors in androgenic alopecia versus alopecia areata. Am. J. Dermatopathol. 1998, 20, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A. Stress and the hair follicle: Exploring the connections. Am. J. Pathol. 2003, 162, 709–712. [Google Scholar] [CrossRef]
- Biondo, S.; Sinclair, R. Quality of life in Australian women with female pattern hair loss. Open Dermatol. J. 2010, 4, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Ohnemus, U.; Uenalan, M.; Inzunza, J.; Gustafsson, J.-A.; Paus, R. The hair follicle as an estrogen target and source. Endocr. Rev. 2006, 27, 677–706. [Google Scholar] [CrossRef] [Green Version]
- Niiyama, S.; Happle, R.; Hoffmann, R. Influence of estrogens on the androgen metabolism in different subunits of human hair follicles. Eur. J. Dermatol. 2001, 11, 195–198. [Google Scholar]
- Springer, K.; Brown, M.; Stulberg, D.L. Common hair loss disorders. Am. Fam. Physician 2003, 68, 93–102. [Google Scholar]
- Riedel-Baima, B.; Riedel, A. Female pattern hair loss may be triggered by low oestrogen to androgen ratio. Endocr. Regul. 2008, 42, 13–16. [Google Scholar]
- Shin, H.S.; Lee, S.H.; Kim, D.H.; An, J.S.; Kwon, O.S.; Eun, H.C.; Kim, K.H. The efficacy and safety of AP-FHG0604T on female pattern hair loss: A randomized double-blind placebo-controlled clinical trial. Korean J. Dermatol. 2007, 45, 119–126. [Google Scholar]
- Kim, J.H.; Lee, S.Y.; Lee, H.J.; Yoon, N.Y.; Lee, W.S. The Efficacy and Safety of 17alpha-Estradiol (Ell-Cranell(R) alpha 0.025%) Solution on Female Pattern Hair Loss: Single Center, Open-Label, Non-Comparative, Phase IV Study. Ann. Dermatol. 2012, 24, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Munkhbayar, S.; Jang, S.; Cho, A.R.; Choi, S.J.; Shin, C.Y.; Eun, H.C.; Kim, K.H.; Kwon, O. Role of Arachidonic Acid in Promoting Hair Growth. Ann. Dermatol. 2016, 28, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, S.; Hozumi, Y.; Kondo, S. Influence of prostaglandin F2alpha and its analogues on hair regrowth and follicular melanogenesis in a murine model. Exp. Dermatol. 2005, 14, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Levy, L.L.; Emer, J.J. Female pattern alopecia: Current perspectives. Int. J. Women’s Health 2013, 5, 541. [Google Scholar]
- Soler, A.P.; Gilliard, G.; Megosh, L.C.; O’Brien, T.G. Modulation of murine hair follicle function by alterations in ornithine decarboxylase activity. J. Investig. Dermatol. 1996, 106, 1108–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nancarrow, M.J.; Nesci, A.; Hynd, P.I.; Powell, B.C. Dynamic expression of ornithine decarboxylase in hair growth. Mech. Dev. 1999, 84, 161–164. [Google Scholar] [CrossRef]
- Lee, Y.R.; Lew, B.L.; Sim, W.Y.; Lee, J.; Hong, J.; Chung, B.C. Altered polyamine profiling in the hair of patients with androgenic alopecia and alopecia areata. J. Dermatol. 2019, 46, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Lee, J.; Lew, B.L.; Sim, W.Y.; Hong, J.; Chung, B.C. Distribution of polyamines may be altered in different scalp regions of patients with hair loss. Exp. Dermatol. 2019, 28, 1083–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.R.; Im, E.; Kim, H.; Lew, B.L.; Sim, W.Y.; Lee, J.; Oh, H.B.; Paeng, K.J.; Hong, J.; Chung, B.C. Untargeted Metabolomics and Steroid Signatures in Urine of Male Pattern Baldness Patients after Finasteride Treatment for a Year. Metabolites 2020, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.R.; An, K.Y.; Jeon, J.; Kim, N.K.; Lee, J.W.; Hong, J.; Chung, B.C. Untargeted Metabolomics and Polyamine Profiling in Serum before and after Surgery in Colorectal Cancer Patients. Metabolites 2020, 10, 487. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
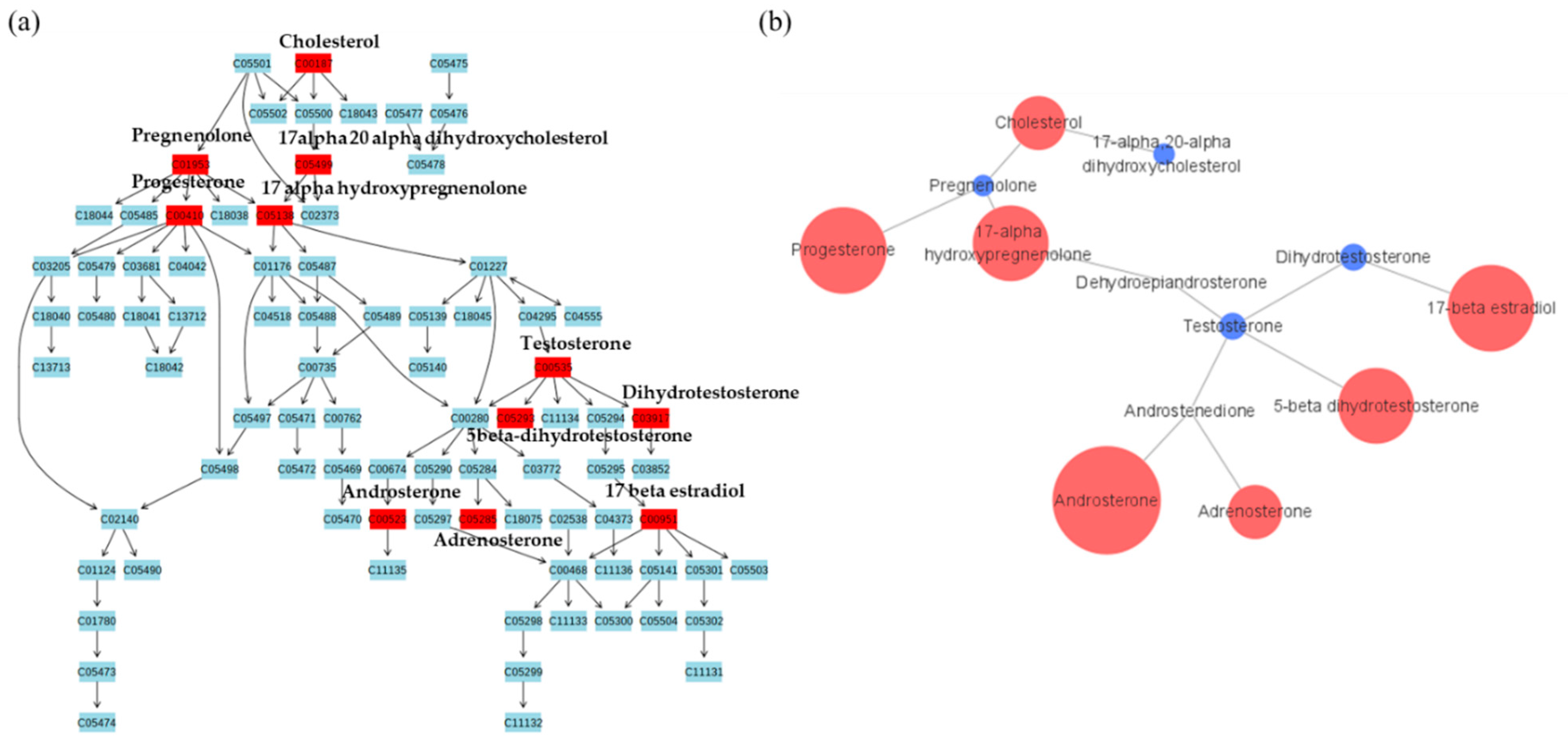{kind=link}
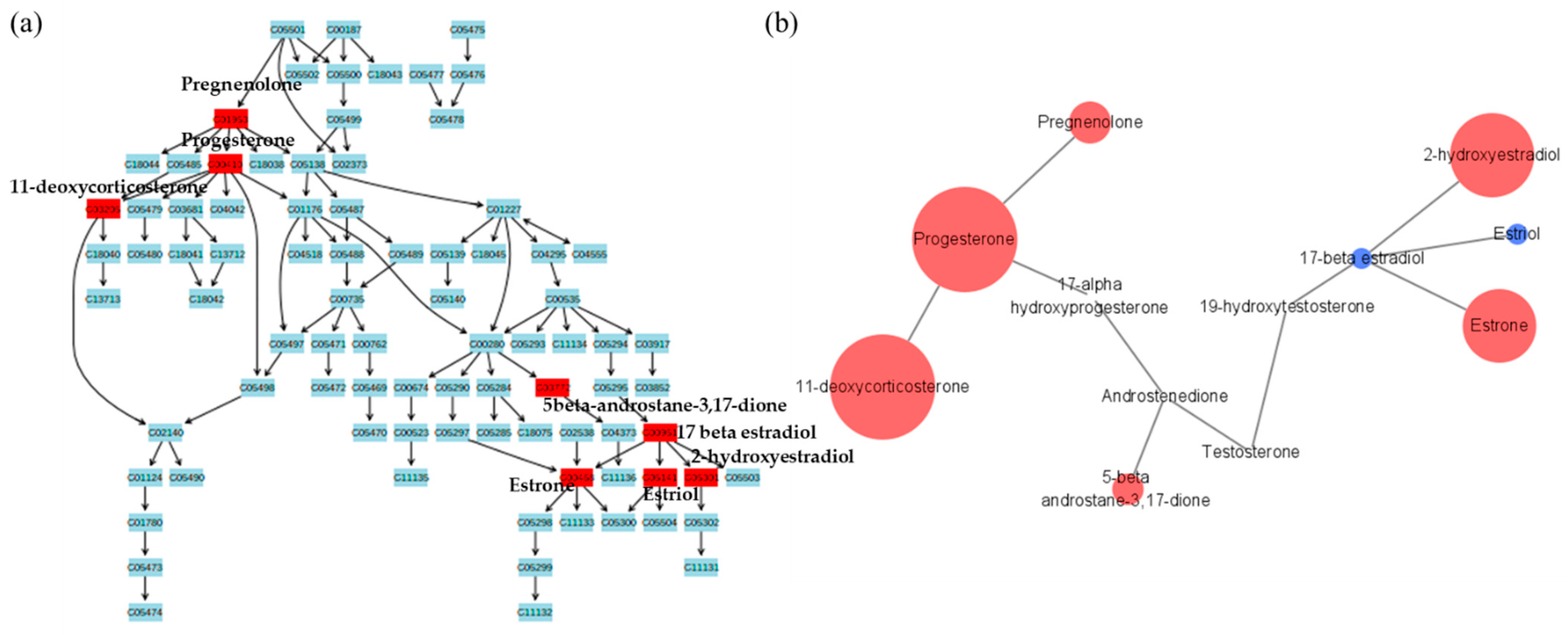{kind=link}
| Metabolites | Related Pathways | VIP | p-Value | FC | Regulation |
|---|---|---|---|---|---|
| l-Ornithine | Arginine and proline metabolism | 1.1985 | 0.003 | 1.5919753 | Up |
| Cholesterol | Steroid hormone biosynthesis | 1.0319 | 0.004 | 2.0443951 | Up |
| Progesterone | Steroid hormone biosynthesis | 1.8311 | <0.0001 | 10.00754 | Up |
| Androsterone | Steroid hormone biosynthesis | 1.0189 | <0.0001 | 23.17902 | Up |
| Prostaglandin F2alpha | Arachidonic acid metabolism | 3.0556 | 0.001 | 0.2634053 | Down |
| Tryptophan | Unknown | 2.2307 | <0.0001 | 2.4243554 | Up |
| Estradiol | Steroid hormone biosynthesis | 3.8042 | <0.0001 | 9.6410812 | Up |
| N-Acetylserotonin | Tyrptophan metabolism | 2.237 | <0.0001 | 15.45131 | Up |
| 5-Aminopentanamide | Unknown | 1.2417 | <0.0001 | 48.086484 | Up |
| Unknown (RT = 5.21) | Unknown | 1.0823 | 0.023 | 3.6537206 | Up |
| Lanosterol | Steroid biosynthesis | 1.1285 | 0.038 | 0.488275 | Down |
| Pregnenolone | Steroid hormone biosynthesis | 2.3772 | 0.001 | 0.4563306 | Down |
| 2-Methylmaleate | Unknown | 1.2134 | <0.0001 | 2.6569257 | Up |
| l-Cystathionine | Cystein and methionine metabolism | 1.2592 | <0.0001 | 0.346639 | Down |
| 3-Oxododecanoic acid | Unknown | 1.828 | <0.0001 | 8.1830723 | Up |
| Nonane-4,6-dione | Unknown | 1.4105 | 0.015 | 0.0427089 | Down |
| Dodecanoic acid | Fatty acid biosynthesis | 1.6129 | 0.005 | 27.655866 | Up |
| l-Palmitoylcarnitine | Fatty acid degradation | 4.6032 | <0.0001 | 15.272926 | Up |
| Dopamine | Tyrosine metabolism | 1.4983 | <0.0001 | 28.58511 | Up |
| 5alpha-Dihydrotestosterone | Steroid hormone biosynthesis | 4.0064 | 0.004 | 12.736311 | Up |
| Diethyl 2-methyl-3-oxosuccinate | Unknown | 1.4048 | <0.0001 | 16.252694 | Up |
| 17alpha-Hydroxypregnenolone | Steroid hormone biosynthesis | 11.727 | <0.0001 | 5.4864425 | Up |
| Adrenosterone | Steroid hormone biosynthesis | 2.4995 | <0.0001 | 2.4143371 | Up |
| 17alpha,20alpha-Dihydroxycholesterol | Steroid hormone biosynthesis | 1.6553 | 0.002 | 0.1035019 | Down |
| l-Metanephrine | Tyrosine metabolism | 1.2238 | <0.0001 | 211.3841 | Up |
| Tetradecanoic acid | Fatty acid biosynthesis | 1.126 | <0.0001 | 11.466312 | Up |
| p-Cymene | Unknown | 7.1135 | <0.0001 | 24.273712 | Up |
| Unknown (RT = 3.76) | Unknown | 1.264 | 0.011 | 2.7952495 | Up |
| Unknown (RT = 4.15) | Unknown | 1.7684 | <0.0001 | 0.4185244 | Down |
| Docosanoic acid | Unknown | 1.4008 | <0.0001 | 6.895777 | Up |
| Humulene | Unknown | 1.2381 | <0.0001 | 24.509825 | Up |
| beta-Citronellol | Unknown | 1.2889 | <0.0001 | 2.2233273 | Up |
| Thymol | Unknown | 1.2472 | 0.02 | 6.2627942 | Up |
| Tridecane | Unknown | 1.4906 | 0.013 | 5.6917082 | Up |
| 4-Propylphenol | Unknown | 1.2452 | <0.0001 | 0.3275366 | Down |
| Tetraethylenepentamine | Unknown | 1.6745 | <0.0001 | 8.9096476 | Up |
| Traumatic acid | Unknown | 4.0178 | 0.007 | 3.5804527 | Up |
| Unknown (RT = 5.43) | Unknown | 2.2358 | <0.0001 | 339.30032 | Up |
| Neocnidilide | Unknown | 1.4039 | 0.002 | 1.4515461 | Up |
| Testosterone | Steroid hormone biosynthesis | 4.334 | 0.002 | 0.4662096 | Down |
| 7alpha-Hydroxytestosterone | Unknown | 2.3012 | <0.0001 | 1.8760545 | Up |
| 5beta-Dihydrotestosterone | Steroid hormone biosynthesis | 2.5426 | 0.01 | 6.9586924 | Up |
| Metabolites | Related Pathways | VIP | p Value | FC | Regulation |
|---|---|---|---|---|---|
| l-Ornithine | Arginine biosynthesis | 1.3668 | <0.0001 | 11.910219 | Up |
| Thymine | Pyrimidine metabolism | 1.9371 | 0.025 | 15.622769 | Up |
| Progesterone | Steroid hormone biosynthesis | 1.1846 | <0.0001 | 17.966704 | Up |
| N1-Acetylspermidine | Unknown | 1.8138 | <0.0001 | 5.4335943 | Up |
| Prostaglandin F2alpha | Arachidonic acid metabolism | 1.0325 | 0.045 | 0.5604433 | Down |
| Estrone | Steroid hormone biosynthesis | 1.7482 | <0.0001 | 10.18276 | Up |
| Octadecanal | Unknown | 1.14 | 0.001 | 4.7250936 | Up |
| Pregnenolone | Steroid hormone biosynthesis | 9.4143 | <0.0001 | 6.1234272 | Up |
| l-Cystathionine | Glycine, serine and threonine metabolism | 1.2179 | <0.0001 | 0.3661377 | Down |
| Unknown (RT = 5.22) | Unknown | 1.1014 | <0.0001 | 7.8823964 | Up |
| 5beta-Androstane-3,17-dione | Steroid hormone biosynthesis | 2.7125 | <0.0001 | 3.8285677 | Up |
| 7alpha-Hydroxytestosterone | Unknown | 1.273 | <0.0001 | 0.3856094 | Down |
| Tetradecanoic acid | Fatty acid biosynthesis | 6.6464 | <0.0001 | 6.082907 | Up |
| Unknown (RT = 4.48) | Unknown | 1.6103 | 0.002 | 1.4591549 | Up |
| Tetracosanoic acid | Unknown | 4.0729 | 0.007 | 0.6894739 | Down |
| Fucosterol | Unknown | 4.3796 | 0.011 | 0.7134959 | Down |
| Ribalinium | Unknown | 1.108 | <0.0001 | 4.9206557 | Up |
| Teprenone | Unknown | 1.5799 | <0.0001 | 9.3409894 | Up |
| Episterol | Steroid biosynthesis | 1.3281 | 0.043 | 0.4078797 | Down |
| Estriol | Steroid hormone biosynthesis | 2.5591 | <0.0001 | 0.5797319 | Down |
| Ceramide | Sphingolipid metabolism | 1.2892 | 0.01 | 9.651607 | Up |
| Estradiol | Steroid hormone biosynthesis | 5.5694 | 0.04 | 0.8441624 | Down |
| Leukotriene B4 | Arachidonic acid metabolism | 1.2626 | <0.0001 | 22.611004 | Up |
| 11-Deoxycorticosterone | Steroid hormone biosynthesis | 3.5377 | <0.0001 | 18.558709 | Up |
| 2-OH-Estradiol | Steroid hormone biosynthesis | 2.3525 | 0.02 | 13.21151 | Up |
| 20-HETE | Arachidonic acid metabolism | 2.0165 | 0.02 | 1.3674874 | Up |
| LC Condition | |||
| Instrument | ACQUITY™ ultra-performance liquid chromatography | ||
| Column | ACQUITY UPLC BEH C18 (2.1 × 100 mm, 1.7 μm) | ||
| Column Temperature | 40 °C | ||
| Sampler Temperature | 4 °C | ||
| Injection Volume | 5 μL | ||
| Flow Rate | 0.35 mL/min | ||
| Mobile Phase | A: 0.1% formic acid water | ||
| B: 0.1% formic acid acetonitrile | |||
| Gradient System | Time (min) | %A | %B |
| 0 | 95 | 5 | |
| 3 | 50 | 50 | |
| 10 | 5 | 95 | |
| 11.5 | 5 | 95 | |
| 12 | 95 | 5 | |
| 14 | 95 | 5 | |
| MS Condition | |||
| Instrument | Q-Time of flight Premier™ quadrupole/ time-of-flight hybrid mass spectrometer | ||
| Scan Type | Full scan (m/z range: 50–1200) | ||
| Cone Voltage | 2.5 kV | ||
| Capillary Temperature | 120 °C | ||
| Desolvation Temperature | 350 °C | ||
| Cone Gas Flow | 30 L/h | ||
| Capillary Voltage | 2.5 kV | ||
| Desolvation Gas Flow | 600 L/h | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.R.; Lew, B.L.; Sim, W.Y.; Hong, J.; Chung, B.C. Alterations in Pattern Baldness According to Sex: Hair Metabolomics Approach. Metabolites 2021, 11, 178. https://doi.org/10.3390/metabo11030178
Lee YR, Lew BL, Sim WY, Hong J, Chung BC. Alterations in Pattern Baldness According to Sex: Hair Metabolomics Approach. Metabolites. 2021; 11(3):178. https://doi.org/10.3390/metabo11030178
Chicago/Turabian StyleLee, Yu Ra, Bark Lynn Lew, Woo Young Sim, Jongki Hong, and Bong Chul Chung. 2021. "Alterations in Pattern Baldness According to Sex: Hair Metabolomics Approach" Metabolites 11, no. 3: 178. https://doi.org/10.3390/metabo11030178
APA StyleLee, Y. R., Lew, B. L., Sim, W. Y., Hong, J., & Chung, B. C. (2021). Alterations in Pattern Baldness According to Sex: Hair Metabolomics Approach. Metabolites, 11(3), 178. https://doi.org/10.3390/metabo11030178