
Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, and Functions

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Genetics, Biochemistry, and Evolution of DAPG Biosynthesis
2.1. The Phl Biosynthetic Gene Cluster
2.2. Biosynthesis and Degradation of DAPG
2.3. Distribution and Evolution of the Phl Biosynthetic Gene Cluster
3. Regulation of DAPG Biosynthesis
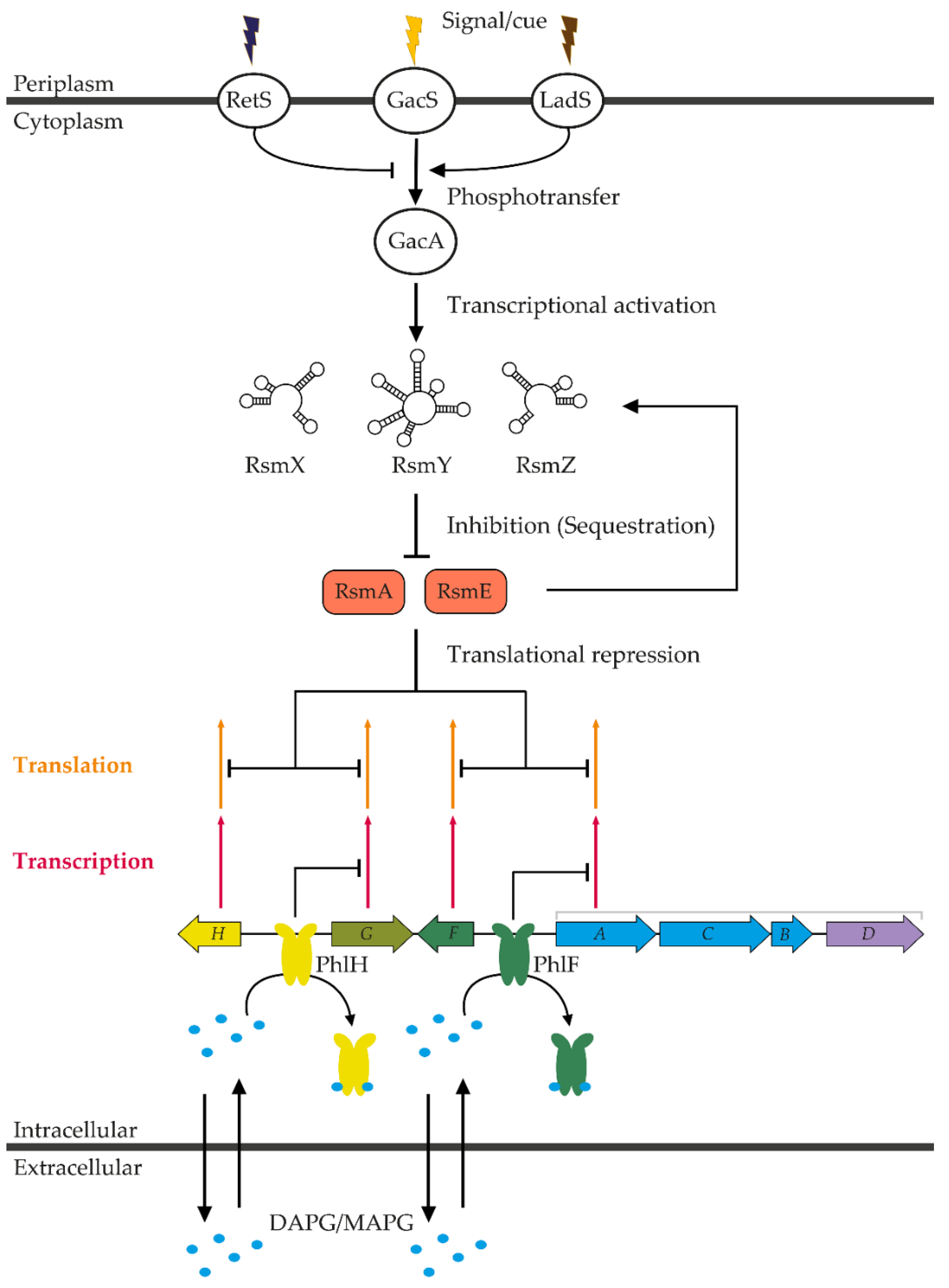
3.1. Regulation by Translational Repressors of the TetR Family
3.2. Regulation by the Gac/Rsm Regulatory Network
3.3. Co-regulation of DAPG and Pyoluteorin Production
3.4. Environmental Factors Influencing DAPG Production
4. Role of DAPG-Producing Pseudomonas spp. in Natural Soil Suppressiveness
4.1. Take-All Decline
4.2. Natural Soil Suppressiveness to Tobacco Black Root Rot
4.3. Role of DAPG-Producing Pseudomonas in Other Suppressive Soils
5. Mode of Action
5.1. Direct Inhibition of Soil-Borne Plant Pathogens
5.2. DAPG in Plant-Bacteria Interaction
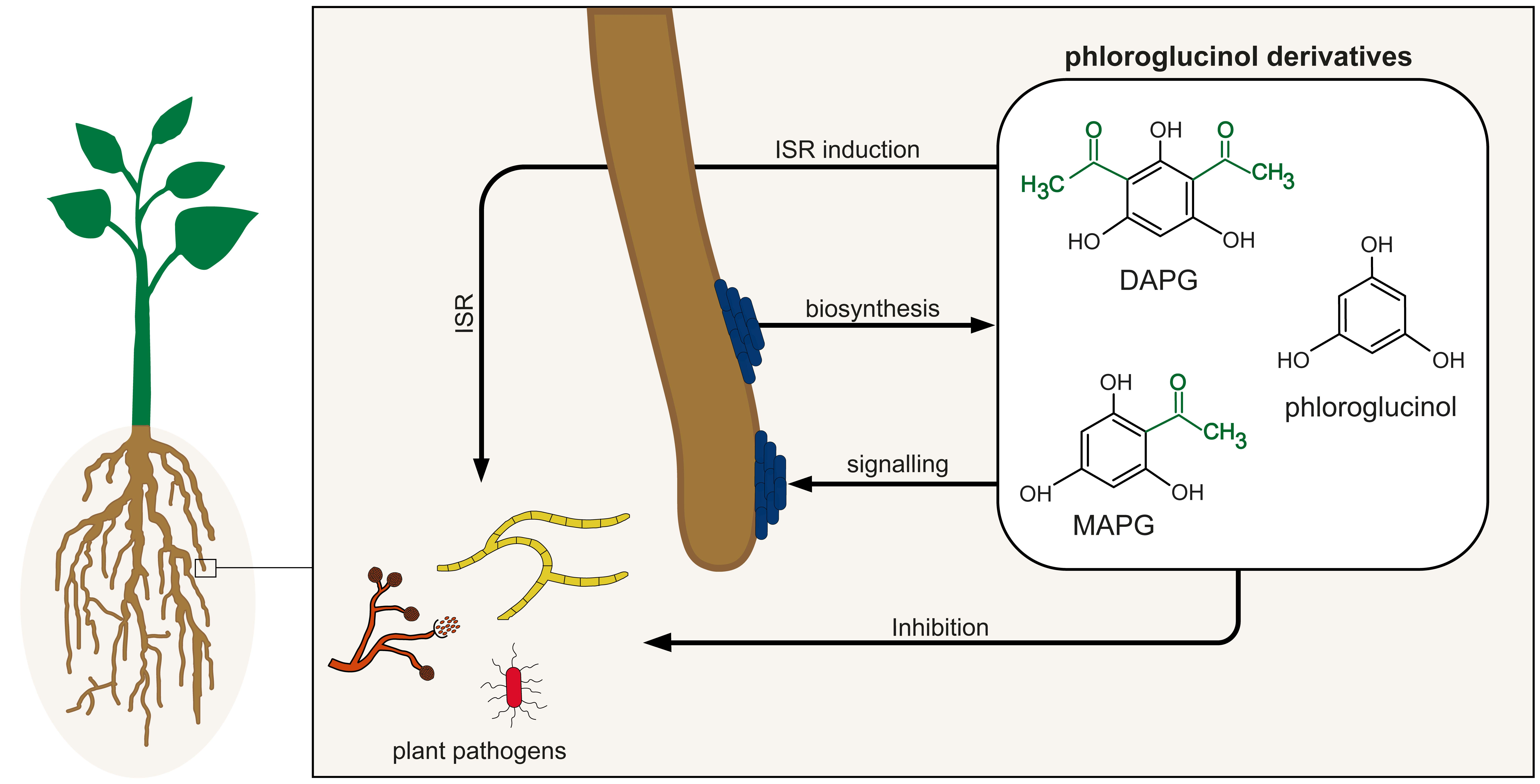
5.3. Phloroglucinol Derivatives as Signalling Molecules in the Rhizosphere
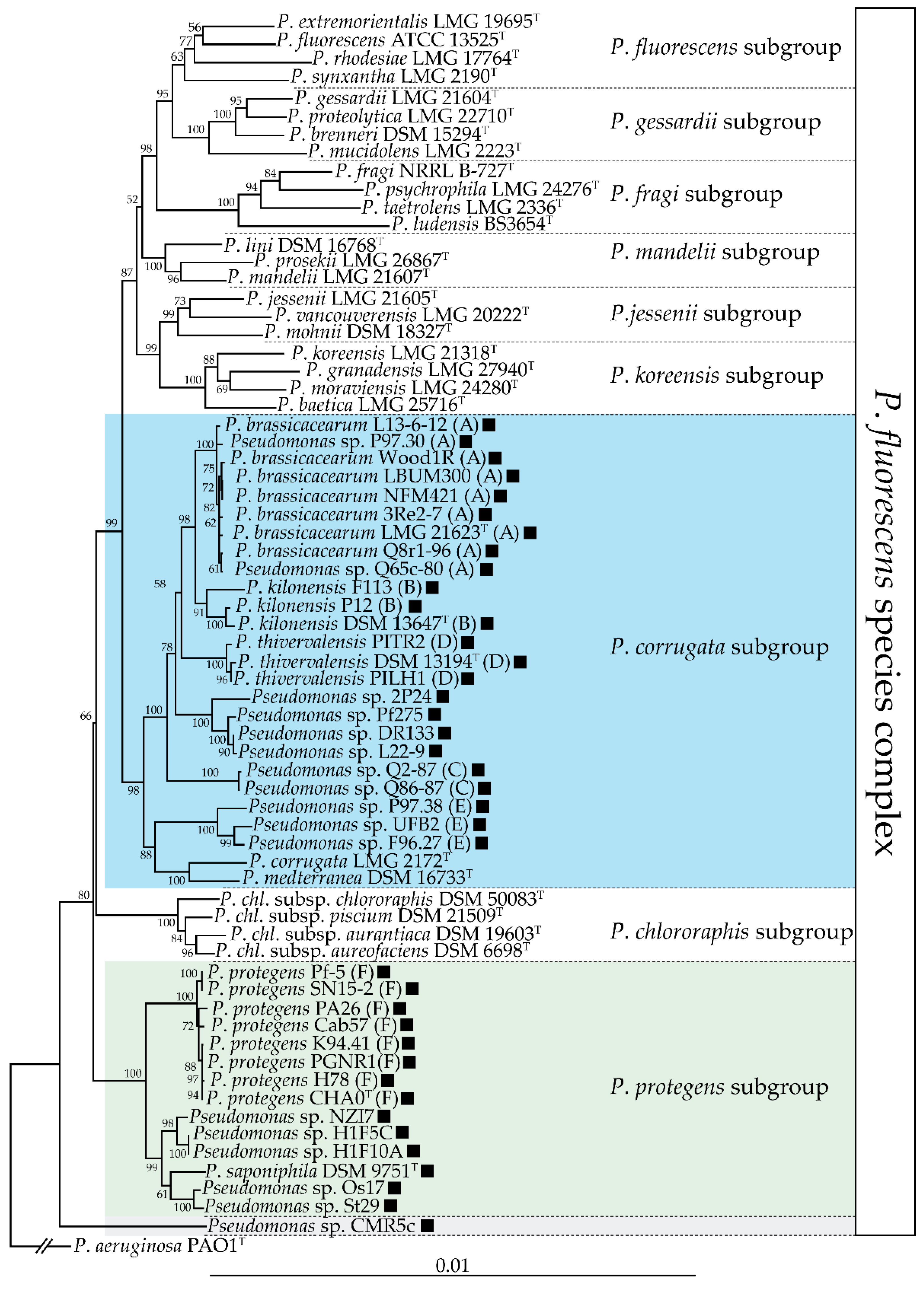
6. Genetic and Genomic Diversity of DAPG-Producing Pseudomonas spp.
7. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Bridi, H.; de Carvalho Meirelles, G.; von Poser, G.L. Structural diversity and biological activities of phloroglucinol derivatives from Hypericum species. Phytochemistry 2018, 155, 203–232. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Zhang, W.; Smid, S. Phlorotannins: A review on biosynthesis, chemistry and bioactivity. Food Biosci. 2020, 100832. [Google Scholar] [CrossRef]
- Weller, D.M.; Landa, B.; Mavrodi, O.; Schroeder, K.; De La Fuente, L.; Blouin Bankhead, S.; Allende Molar, R.; Bonsall, R.; Mavrodi, D.; Thomashow, L. Role of 2, 4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in the defense of plant roots. Plant Biol. 2007, 9, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Müller, T.; Behrendt, U.; Ruppel, S.; von der Waydbrink, G.; Müller, M.E. Fluorescent pseudomonads in the phyllosphere of wheat: Potential antagonists against fungal phytopathogens. Curr. Microbiol. 2016, 72, 383–389. [Google Scholar] [CrossRef]
- Myers, J.M.; Ramsey, J.P.; Blackman, A.L.; Nichols, A.E.; Minbiole, K.P.; Harris, R.N. Synergistic inhibition of the lethal fungal pathogen Batrachochytrium dendrobatidis: The combined effect of symbiotic bacterial metabolites and antimicrobial peptides of the frog Rana muscosa. J. Chem. Ecol. 2012, 38, 958–965. [Google Scholar] [CrossRef]
- Isnansetyo, A.; Cui, L.; Hiramatsu, K.; Kamei, Y. Antibacterial activity of 2, 4-diacetylphloroglucinol produced by Pseudomonas sp. AMSN isolated from a marine alga, against vancomycin-resistant Staphylococcus aureus. Int. J. Antimicrob. Agents 2003, 22, 545–547. [Google Scholar] [CrossRef]
- Kwak, Y.-S.; Weller, D.M. Take-all of wheat and natural disease suppression: A review. Plant Pathol. J. 2013, 29, 125. [Google Scholar] [CrossRef]
- Vincent, M.N.; Harrison, L.; Brackin, J.; Kovacevich, P.; Mukerji, P.; Weller, D.; Pierson, E. Genetic analysis of the antifungal activity of a soilborne Pseudomonas aureofaciens strain. Appl. Environ. Microbiol. 1991, 57, 2928–2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangera, M.G.; Thomashow, L.S. Characterization of a genomic locus required for synthesis of the antibiotic 2, 4-diacetylphloroglucinol by the biological control agent Pseudomonas fluorescens Q2-87. Mol. Plant Microbe Interact. 1996, 9, 83. [Google Scholar] [CrossRef]
- Fenton, A.; Stephens, P.; Crowley, J.; O’callaghan, M.; O’gara, F. Exploitation of gene (s) involved in 2, 4-diacetylphloroglucinol biosynthesis to confer a new biocontrol capability to a Pseudomonas strain. Appl. Environ. Microbiol. 1992, 58, 3873–3878. [Google Scholar] [CrossRef] [Green Version]
- Keel, C.; Schnider, U.; Maurhofer, M.; Voisard, C.; Laville, J.; Burger, U.; Wirthner, P.; Haas, D.; Défago, G. Suppression of Root Diseases by Pseudomonas fluorescens CHA0: Importance of the Bacterial Secondary Metabolite 2,4-diacetylphloroglucinol. Mol. Plant Microbe Interact. 1992, 5, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Bangera, M.G.; Thomashow, L.S. Identification and characterization of a gene cluster for synthesis of the polyketide antibiotic 2, 4-diacetylphloroglucinol from Pseudomonas fluorescens Q2-87. J. Bacteriol. 1999, 181, 3155–3163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnider-Keel, U.; Seematter, A.; Maurhofer, M.; Blumer, C.; Duffy, B.; Gigot-Bonnefoy, C.; Reimmann, C.; Notz, R.; Défago, G.; Haas, D. Autoinduction of 2, 4-diacetylphloroglucinol biosynthesis in the biocontrol agent Pseudomonas fluorescens CHA0 and repression by the bacterial metabolites salicylate and pyoluteorin. J. Bacteriol. 2000, 182, 1215–1225. [Google Scholar] [CrossRef] [Green Version]
- Moynihan, J.A.; Morrissey, J.P.; Coppoolse, E.R.; Stiekema, W.J.; O’Gara, F.; Boyd, E.F. Evolutionary history of the phl gene cluster in the plant-associated bacterium Pseudomonas fluorescens. Appl. Environ. Microbiol. 2009, 75, 2122–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, A.; Saitou, H.; Mori, T.; Matano, I.; Sugisaki, H.; Maruyama, K. Molecular and catalytic properties of monoacetylphloroglucinol acetyltransferase from Pseudomonas sp. YGJ3. Biosci. Biotechnol. Biochem. 2012, 76, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almario, J.; Bruto, M.; Vacheron, J.; Prigent-Combaret, C.; Moënne-Loccoz, Y.; Muller, D. Distribution of 2, 4-diacetylphloroglucinol biosynthetic genes among the Pseudomonas spp. reveals unexpected polyphyletism. Front. Microbiol. 2017, 8, 1218. [Google Scholar] [CrossRef] [Green Version]
- Achkar, J.; Xian, M.; Zhao, H.; Frost, J. Biosynthesis of phloroglucinol. J. Am. Chem. Soc. 2005, 127, 5332–5333. [Google Scholar] [CrossRef]
- Shimizu, Y.; Ogata, H.; Goto, S. Type III polyketide synthases: Functional classification and phylogenomics. ChemBioChem 2017, 18, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Zha, W.; Rubin-Pitel, S.B.; Zhao, H. Characterization of the substrate specificity of PhlD, a type III polyketide synthase from Pseudomonas fluorescens. J. Biol. Chem. 2006, 281, 32036–32047. [Google Scholar] [CrossRef]
- Shanahan, P.; Glennon, J.; Crowley, J.; Donnelly, D.; O’Gara, F. Liquid chromatographic assay of microbially derived phloroglucinol antibiotics for establishing the biosynthetic route to production, and the factors affecting their regulation. Anal. Chim. Acta 1993, 272, 271–277. [Google Scholar] [CrossRef]
- Pavkov-Keller, T.; Schmidt, N.G.; Żądło-Dobrowolska, A.; Kroutil, W.; Gruber, K. Structure and catalytic mechanism of a bacterial Friedel–Crafts acylase. ChemBioChem 2019, 20, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, N.G.; Żądło-Dobrowolska, A.; Ruppert, V.; Höflehner, C.; Wiltschi, B.; Kroutil, W. Molecular cloning, expression, and characterization of acyltransferase from Pseudomonas protegens. Appl. Microbiol. Biotechnol. 2018, 102, 6057–6068. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; Kazemi, M.; Żądło-Dobrowolska, A.; Kroutil, W.; Himo, F. Mechanism of Biocatalytic Friedel–Crafts Acylation by Acyltransferase from Pseudomonas protegens. ACS Catal. 2020, 10, 570–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, N.G.; Kroutil, W. Acyl donors and additives for the biocatalytic Friedel–Crafts acylation. Eur. J. Org. Chem. 2017, 2017, 5865–5871. [Google Scholar] [CrossRef]
- Żądło-Dobrowolska, A.; Schmidt, N.G.; Kroutil, W. Thioesters as Acyl Donors in Biocatalytic Friedel-Crafts-type Acylation Catalyzed by Acyltransferase from Pseudomonas protegens. ChemCatChem 2019, 11, 1064–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, N.G.; Pavkov-Keller, T.; Richter, N.; Wiltschi, B.; Gruber, K.; Kroutil, W. Biocatalytic Friedel–crafts acylation and fries reaction. Angew. Chem. Int. Ed. 2017, 56, 7615–7619. [Google Scholar] [CrossRef]
- Bottiglieri, M.; Keel, C. Characterization of PhlG, a hydrolase that specifically degrades the antifungal compound 2, 4-diacetylphloroglucinol in the biocontrol agent Pseudomonas fluorescens CHA0. Appl. Environ. Microbiol. 2006, 72, 418–427. [Google Scholar] [CrossRef] [Green Version]
- He, Y.-X.; Huang, L.; Xue, Y.; Fei, X.; Teng, Y.-B.; Rubin-Pitel, S.B.; Zhao, H.; Zhou, C.-Z. Crystal structure and computational analyses provide insights into the catalytic mechanism of 2, 4-diacetylphloroglucinol hydrolase PhlG from Pseudomonas fluorescens. J. Biol. Chem. 2010, 285, 4603–4611. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Sanz, D.; Meier-Kolthoff, J.P.; Göker, M.; Martín, M.; Rivilla, R.; Redondo-Nieto, M. Genomic and Genetic Diversity within the Pseudomonas fluorescens Complex. PLoS ONE 2016, 11, e0150183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnyk, R.A.; Hossain, S.S.; Haney, C.H. Convergent gain and loss of genomic islands drive lifestyle changes in plant-associated Pseudomonas. ISME J. 2019, 13, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Frapolli, M.; Défago, G.; Moënne-Loccoz, Y. Multilocus sequence analysis of biocontrol fluorescent Pseudomonas spp. producing the antifungal compound 2, 4-diacetylphloroglucinol. Environ. Microbiol. 2007, 9, 1939–1955. [Google Scholar] [CrossRef]
- Catara, V.; Sutra, L.; Morineau, A.; Achouak, W.; Christen, R.; Gardan, L. Phenotypic and genomic evidence for the revision of Pseudomonas corrugata and proposal of Pseudomonas mediterranea sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Trantas, E.A.; Licciardello, G.; Almeida, N.F.; Witek, K.; Strano, C.P.; Duxbury, Z.; Ververidis, F.; Goumas, D.E.; Jones, J.D.; Guttman, D.S. Comparative genomic analysis of multiple strains of two unusual plant pathogens: Pseudomonas corrugata and Pseudomonas mediterranea. Front. Microbiol. 2015, 6, 811. [Google Scholar] [CrossRef]
- Iavicoli, A.; Boutet, E.; Buchala, A.; Métraux, J.-P. Induced systemic resistance in Arabidopsis thaliana in response to root inoculation with Pseudomonas fluorescens CHA0. Mol. Plant Microbe Interact. 2003, 16, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.M.; Mavrodi, D.V.; van Pelt, J.A.; Pieterse, C.M.; van Loon, L.C.; Bakker, P.A. Induced systemic resistance in Arabidopsis thaliana against Pseudomonas syringae pv. tomato by 2, 4-diacetylphloroglucinol-producing Pseudomonas fluorescens. Phytopathology 2012, 102, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Chae, D.-H.; Kim, D.-R.; Cheong, M.S.; Lee, Y.B.; Kwak, Y.-S. Investigating the induced systemic resistance mechanism of 2, 4-Diacetylphloroglucinol (DAPG) using DAPG hydrolase-transgenic Arabidopsis. Plant Pathol. J. 2020, 36, 255. [Google Scholar] [CrossRef]
- Biessy, A.; Novinscak, A.; Blom, J.; Léger, G.; Thomashow, L.S.; Cazorla, F.M.; Josic, D.; Filion, M. Diversity of phytobeneficial traits revealed by whole-genome analysis of worldwide-isolated phenazine-producing Pseudomonas spp. Environ. Microbiol. 2019, 21, 437–455. [Google Scholar] [CrossRef] [Green Version]
- Frapolli, M.; Pothier, J.F.; Défago, G.; Moënne-Loccoz, Y. Evolutionary history of synthesis pathway genes for phloroglucinol and cyanide antimicrobials in plant-associated fluorescent pseudomonads. Mol. Phylogenet. Evol. 2012, 63, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Kidarsa, T.A.; Goebel, N.C.; Zabriskie, T.M.; Loper, J.E. Phloroglucinol mediates cross-talk between the pyoluteorin and 2, 4-diacetylphloroglucinol biosynthetic pathways in Pseudomonas fluorescens Pf-5. Mol. Microbiol. 2011, 81, 395–414. [Google Scholar] [CrossRef]
- Cuthbertson, L.; Nodwell, J.R. The TetR family of regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, A.; Morrissey, J.P.; Marquez, P.C.; Sheehan, M.M.; Delany, I.R.; O’Gara, F. Characterization of interactions between the transcriptional repressor PhlF and its binding site at the phlA promoter in Pseudomonas fluorescens F113. J. Bacteriol. 2002, 184, 3008–3016. [Google Scholar] [CrossRef] [Green Version]
- Delany, I.; Sheehan, M.M.; Fenton, A.; Bardin, S.; Aarons, S.; O’Gara, F. Regulation of production of the antifungal metabolite 2, 4-diacetylphloroglucinol in Pseudomonas fluorescens F113: Genetic analysis of phlF as a transcriptional repressor. Microbiology 2000, 146, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Yang, R.; Zhao, R.-X.; Han, J.-T.; Jia, W.-J.; Li, D.-Y.; Wang, Y.; Zhang, N.; Wu, Y.; Zhang, L.-Q. Transcriptional regulator PhlH modulates 2, 4-diacetylphloroglucinol biosynthesis in response to the biosynthetic intermediate and end product. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Lee, S.H.; Seeve, C.; Yu, J.M.; Pierson, L.S.; Pierson, E.A. Roles of the Gac-Rsm pathway in the regulation of phenazine biosynthesis in Pseudomonas chlororaphis 30-84. Microbiologyopen 2013, 2, 505–524. [Google Scholar] [CrossRef]
- Laville, J.; Voisard, C.; Keel, C.; Maurhofer, M.; Defago, G.; Haas, D. Global control in Pseudomonas fluorescens mediating antibiotic synthesis and suppression of black root rot of tobacco. Proc. Natl. Acad. Sci. USA 1992, 89, 1562–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimmann, C.; Valverde, C.; Kay, E.; Haas, D. Posttranscriptional repression of GacS/GacA-controlled genes by the RNA-binding protein RsmE acting together with RsmA in the biocontrol strain Pseudomonas fluorescens CHA0. J. Bacteriol. 2005, 187, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Latour, X. The Evanescent GacS Signal. Microorganisms 2020, 8, 1746. [Google Scholar] [CrossRef]
- Goodman, A.L.; Merighi, M.; Hyodo, M.; Ventre, I.; Filloux, A.; Lory, S. Direct interaction between sensor kinase proteins mediates acute and chronic disease phenotypes in a bacterial pathogen. Genes Dev. 2009, 23, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Humair, B.; González, N.; Mossialos, D.; Reimmann, C.; Haas, D. Temperature-responsive sensing regulates biocontrol factor expression in Pseudomonas fluorescens CHA0. ISME J. 2009, 3, 955–965. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Yamada, K.; Haas, D. ppGpp controlled by the Gac/Rsm regulatory pathway sustains biocontrol activity in Pseudomonas fluorescens CHA0. Mol. Plant Microbe Interact. 2012, 25, 1440–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workentine, M.L.; Chang, L.; Ceri, H.; Turner, R.J. The GacS–GacA two-component regulatory system of Pseudomonas fluorescens: A bacterial two-hybrid analysis. FEMS Microbiol. Lett. 2009, 292, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Kay, E.; Dubuis, C.; Haas, D. Three small RNAs jointly ensure secondary metabolism and biocontrol in Pseudomonas fluorescens CHA0. Proc. Natl. Acad. Sci. USA 2005, 102, 17136–17141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humair, B.; Wackwitz, B.; Haas, D. GacA-controlled activation of promoters for small RNA genes in Pseudomonas fluorescens. Appl. Environ. Microbiol. 2010, 76, 1497–1506. [Google Scholar] [CrossRef] [Green Version]
- Valverde, C.; Heeb, S.; Keel, C.; Haas, D. RsmY, a small regulatory RNA, is required in concert with RsmZ for GacA-dependent expression of biocontrol traits in Pseudomonas fluorescens CHA0. Mol. Microbiol. 2003, 50, 1361–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, M.; Lapouge, K.; Duss, O.; Oberstrass, F.C.; Jelesarov, I.; Haas, D.; Allain, F.H. Molecular basis of messenger RNA recognition by the specific bacterial repressing clamp RsmA/CsrA. Nat. Struct. Mol. Biol. 2007, 14, 807. [Google Scholar] [CrossRef]
- Lapouge, K.; Sineva, E.; Lindell, M.; Starke, K.; Baker, C.S.; Babitzke, P.; Haas, D. Mechanism of hcnA mRNA recognition in the Gac/Rsm signal transduction pathway of Pseudomonas fluorescens. Mol. Microbiol. 2007, 66, 341–356. [Google Scholar] [CrossRef]
- Duss, O.; Michel, E.; Diarra dit Konté, N.; Schubert, M.; Allain, F.H.-T. Molecular basis for the wide range of affinity found in Csr/Rsm protein–RNA recognition. Nucleic Acids Res. 2014, 42, 5332–5346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapouge, K.; Perozzo, R.; Iwaszkiewicz, J.; Bertelli, C.; Zoete, V.; Michielin, O.; Scapozza, L.; Haas, D. RNA pentaloop structures as effective targets of regulators belonging to the RsmA/CsrA protein family. RNA Biol. 2013, 10, 1030–1041. [Google Scholar] [CrossRef] [Green Version]
- Babitzke, P.; Romeo, T. CsrB sRNA family: Sequestration of RNA-binding regulatory proteins. Curr. Opin. Microbiol. 2007, 10, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Lapouge, K.; Schubert, M.; Allain, F.H.T.; Haas, D. Gac/Rsm signal transduction pathway of γ-proteobacteria: From RNA recognition to regulation of social behaviour. Mol. Microbiol. 2008, 67, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Kiefer, P.; Reimmann, C.; Keel, C.; Dubuis, C.; Rolli, J.; Vorholt, J.A.; Haas, D. Small RNA-dependent expression of secondary metabolism is controlled by Krebs cycle function in Pseudomonas fluorescens. J. Biol. Chem. 2009, 284, 34976–34985. [Google Scholar] [CrossRef] [Green Version]
- Heeb, S.; Blumer, C.; Haas, D. Regulatory RNA as mediator in GacA/RsmA-dependent global control of exoproduct formation in Pseudomonas fluorescens CHA0. J. Bacteriol. 2002, 184, 1046–1056. [Google Scholar] [CrossRef] [Green Version]
- Nowak-Thompson, B.; Gould, S.J.; Kraus, J.; Loper, J.E. Production of 2, 4-diacetylphloroglucinol by the biocontrol agent Pseudomonas fluorescens Pf-5. Can. J. Microbiol. 1994, 40, 1064–1066. [Google Scholar] [CrossRef]
- Haas, D.; Keel, C.; Laville, J.; Maurhofer, M.; Oberhänsli, T.; Schnider, U.; Voisard, C.; Wüthrich, B.; Defago, G. Secondary metabolites of Pseudomonas fluorescens strain CHA0 involved in the suppression of root diseases. In Advances in Molecular Genetics of Plant-Microbe Interactions; Hennecke, H., Verma, D.P.S., Eds.; Springer: Dordrecht, The Netherlands, 1991; Volume 1, pp. 450–456. [Google Scholar]
- Maurhofer, M.; Keel, C.; Haas, D.; Défago, G. Pyoluteorin production by Pseudomonas fluorescens strain CHA0 is involved in the suppression of Pythium damping-off of cress but not of cucumber. Eur. J. Plant Pathol. 1994, 100, 221–232. [Google Scholar] [CrossRef]
- Brodhagen, M.; Henkels, M.D.; Loper, J.E. Positive autoregulation and signaling properties of pyoluteorin, an antibiotic produced by the biological control organism Pseudomonas fluorescens Pf-5. Appl. Environ. Microbiol. 2004, 70, 1758–1766. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Philmus, B.; Chang, J.H.; Loper, J.E. Novel mechanism of metabolic co-regulation coordinates the biosynthesis of secondary metabolites in Pseudomonas protegens. Elife 2017, 6, e22835. [Google Scholar] [CrossRef]
- Shanahan, P.; O’Sullivan, D.J.; Simpson, P.; Glennon, J.D.; O’Gara, F. Isolation of 2, 4-diacetylphloroglucinol from a fluorescent pseudomonad and investigation of physiological parameters influencing its production. Appl. Environ. Microbiol. 1992, 58, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Duffy, B.K.; Défago, G. Environmental factors modulating antibiotic and siderophore biosynthesis by Pseudomonas fluorescens biocontrol strains. Appl. Environ. Microbiol. 1999, 65, 2429–2438. [Google Scholar] [CrossRef] [Green Version]
- Bacon, C.; Porter, J.; Norred, W.; Leslie, J. Production of fusaric acid by Fusarium species. Appl. Environ. Microbiol. 1996, 62, 4039–4043. [Google Scholar] [CrossRef] [Green Version]
- Notz, R.; Maurhofer, M.; Dubach, H.; Haas, D.; Défago, G. Fusaric acid-producing strains of Fusarium oxysporum alter 2, 4-diacetylphloroglucinol biosynthetic gene expression in Pseudomonas fluorescens CHA0 in vitro and in the rhizosphere of wheat. Appl. Environ. Microbiol. 2002, 68, 2229–2235. [Google Scholar] [CrossRef] [Green Version]
- Duffy, B.K.; Défago, G. Zinc improves biocontrol of Fusarium crown and root rot of tomato by Pseudomonas fluorescens and represses the production of pathogen metabolites inhibitory to bacterial antibiotic biosynthesis. Phytopathology 1997, 87, 1250–1257. [Google Scholar] [CrossRef] [Green Version]
- Jousset, A.; Rochat, L.; Lanoue, A.; Bonkowski, M.; Keel, C.; Scheu, S. Plants respond to pathogen infection by enhancing the antifungal gene expression of root-associated bacteria. Mol. Plant Microbe Interact. 2011, 24, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.Q.; Yan, X.; Zhang, M.Y.; Zhang, L.Q.; He, Y.X. Flavonoids repress production of antifungal 2, 4-DAPG but potentially facilitate root colonization of the rhizobacterium Pseudomonas fluorescens. Environ. Microbiol. 2020, 22, 5073–5089. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.J. Take-all of wheat. Physiol. Mol. Plant Pathol. 2003, 62, 73–86. [Google Scholar] [CrossRef]
- Freeman, J.; Ward, E. Gaeumannomyces graminis, the take-all fungus and its relatives. Mol. Plant Pathol. 2004, 5, 235–252. [Google Scholar] [CrossRef]
- Cook, R.J. Toward cropping systems that enhance productivity and sustainability. Proc. Natl. Acad. Sci. USA 2006, 103, 18389–18394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, D.M.; Raaijmakers, J.M.; Gardener, B.B.; Thomashow, L.S. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raaijmakers, J.M.; Weller, D.M. Natural plant protection by 2, 4-diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils. Mol. Plant Microbe Interact. 1998, 11, 144–152. [Google Scholar] [CrossRef] [Green Version]
- De Souza, J.T.; Weller, D.M.; Raaijmakers, J.M. Frequency, diversity, and activity of 2, 4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in Dutch take-all decline soils. Phytopathology 2003, 93, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raaijmakers, J.M.; Weller, D.M.; Thomashow, L.S. Frequency of antibiotic-producing Pseudomonas spp. in natural environments. Appl. Environ. Microbiol. 1997, 63, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Bonsall, R.F.; Weller, D.M. Effect of population density of Pseudomonas fluorescens on production of 2, 4-diacetylphloroglucinol in the rhizosphere of wheat. Phytopathology 1999, 89, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Kwak, Y.-S.; Bakker, P.A.; Glandorf, D.C.; Rice, J.T.; Paulitz, T.C.; Weller, D.M. Diversity, virulence, and 2, 4-diacetylphloroglucinol sensitivity of Gaeumannomyces graminis var. tritici isolates from Washington State. Phytopathology 2009, 99, 472–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stutz, E.; Défago, G.; Kern, H. Naturally occurring fluorescent Pseudomonads involved in suppression. Phytopathology 1986, 76, 181–185. [Google Scholar] [CrossRef]
- Almario, J.; Muller, D.; Défago, G.; Moënne-Loccoz, Y. Rhizosphere ecology and phytoprotection in soils naturally suppressive to Thielaviopsis black root rot of tobacco. Environ. Microbiol. 2014, 16, 1949–1960. [Google Scholar] [CrossRef] [Green Version]
- Ramette, A.; Moënne-Loccoz, Y.; Défago, G. Prevalence of fluorescent pseudomonads producing antifungal phloroglucinols and/or hydrogen cyanide in soils naturally suppressive or conducive to tobacco black root rot. FEMS Microbiol. Ecol. 2003, 44, 35–43. [Google Scholar] [CrossRef]
- Frapolli, M.; Défago, G.; Moënne-Loccoz, Y. Denaturing gradient gel electrophoretic analysis of dominant 2, 4-diacetylphloroglucinol biosynthetic phlD alleles in fluorescent Pseudomonas from soils suppressive or conducive to black root rot of tobacco. Soil Biol. Biochem. 2010, 42, 649–656. [Google Scholar] [CrossRef]
- Almario, J.; Moënne-Loccoz, Y.; Muller, D. Monitoring of the relation between 2, 4-diacetylphloroglucinol-producing Pseudomonas and Thielaviopsis basicola populations by real-time PCR in tobacco black root-rot suppressive and conducive soils. Soil Biol. Biochem. 2013, 57, 144–155. [Google Scholar] [CrossRef]
- Ramette, A.; Moënne-Loccoz, Y.; Défago, G. Genetic diversity and biocontrol potential of fluorescent pseudomonads producing phloroglucinols and hydrogen cyanide from Swiss soils naturally suppressive or conducive to Thielaviopsis basicola-mediated black root rot of tobacco. FEMS Microbiol. Ecol. 2006, 55, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keel, C.; Voisard, C.; Berling, C.-H.; Kahr, G.; Defago, G. Iron sufficiency, a prerequisite for the suppression of tobacco black root rot by Pseudomonas fluorescens strain CHA0 under gnotobiotic conditions. Phytopathology 1989, 79, 584–589. [Google Scholar] [CrossRef]
- Almario, J.; Prigent-Combaret, C.; Muller, D.; Moënne-Loccoz, Y. Effect of clay mineralogy on iron bioavailability and rhizosphere transcription of 2, 4-diacetylphloroglucinol biosynthetic genes in biocontrol Pseudomonas protegens. Mol. Plant Microbe Interact. 2013, 26, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Kyselková, M.; Kopecký, J.; Frapolli, M.; Défago, G.; Ságová-Marečková, M.; Grundmann, G.L.; Moënne-Loccoz, Y. Comparison of rhizobacterial community composition in soil suppressive or conducive to tobacco black root rot disease. ISME J. 2009, 3, 1127–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landa, B.B.; Mavrodi, O.V.; Raaijmakers, J.M.; Gardener, B.B.M.; Thomashow, L.S.; Weller, D.M. Differential ability of genotypes of 2, 4-diacetylphloroglucinol-producing Pseudomonas fluorescens strains to colonize the roots of pea plants. Appl. Environ. Microbiol. 2002, 68, 3226–3237. [Google Scholar] [CrossRef] [Green Version]
- Mazurier, S.; Corberand, T.; Lemanceau, P.; Raaijmakers, J.M. Phenazine antibiotics produced by fluorescent pseudomonads contribute to natural soil suppressiveness to Fusarium wilt. ISME J. 2009, 3, 977–991. [Google Scholar] [CrossRef]
- Cronin, D.; Moënne-Loccoz, Y.; Fenton, A.; Dunne, C.; Dowling, D.N.; O’Gara, F. Ecological interaction of a biocontrol Pseudomonas fluorescens strain producing 2, 4-diacetylphloroglucinol with the soft rot potato pathogen Erwinia carotovora subsp. atroseptica. FEMS Microbiol. Ecol. 1997, 23, 95–106. [Google Scholar] [CrossRef]
- Lanteigne, C.; Gadkar, V.J.; Wallon, T.; Novinscak, A.; Filion, M. Production of DAPG and HCN by Pseudomonas sp. LBUM300 contributes to the biological control of bacterial canker of tomato. Phytopathology 2012, 102, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Bonsall, R.F.; Weller, D.M.; Thomashow, L.S. Quantification of 2, 4-diacetylphloroglucinol produced by fluorescent Pseudomonas spp. in vitro and in the rhizosphere of wheat. Appl. Environ. Microbiol. 1997, 63, 951–955. [Google Scholar] [CrossRef] [Green Version]
- Okubara, P.A.; Bonsall, R.F. Accumulation of Pseudomonas-derived 2, 4-diacetylphloroglucinol on wheat seedling roots is influenced by host cultivar. Biol. Control 2008, 46, 322–331. [Google Scholar] [CrossRef]
- Kwak, Y.-S.; Bonsall, R.F.; Okubara, P.A.; Paulitz, T.C.; Thomashow, L.S.; Weller, D.M. Factors impacting the activity of 2, 4-diacetylphloroglucinol-producing Pseudomonas fluorescens against take-all of wheat. Soil Biol. Biochem. 2012, 54, 48–56. [Google Scholar] [CrossRef]
- Watt, M.; Hugenholtz, P.; White, R.; Vinall, K. Numbers and locations of native bacteria on field-grown wheat roots quantified by fluorescence in situ hybridization (FISH). Environ. Microbiol. 2006, 8, 871–884. [Google Scholar] [CrossRef] [PubMed]
- de Souza, J.T.; Arnould, C.; Deulvot, C.; Lemanceau, P.; Gianinazzi-Pearson, V.; Raaijmakers, J.M. Effect of 2, 4-diacetylphloroglucinol on Pythium: Cellular responses and variation in sensitivity among propagules and species. Phytopathology 2003, 93, 966–975. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Von Tiedemann, A. 2, 4-Diacetylphloroglucinol suppresses zoosporogenesis and impairs motility of Peronosporomycete zoospores. World J. Microbiol. Biotechnol. 2011, 27, 2071–2079. [Google Scholar] [CrossRef] [Green Version]
- Troppens, D.M.; Chu, M.; Holcombe, L.J.; Gleeson, O.; O’Gara, F.; Read, N.D.; Morrissey, J.P. The bacterial secondary metabolite 2, 4-diacetylphloroglucinol impairs mitochondrial function and affects calcium homeostasis in Neurospora crassa. Fungal Genet. Biol. 2013, 56, 135–146. [Google Scholar] [CrossRef]
- Gleeson, O.; O’Gara, F.; Morrissey, J.P. The Pseudomonas fluorescens secondary metabolite 2, 4 diacetylphloroglucinol impairs mitochondrial function in Saccharomyces cerevisiae. Antonie Leeuwenhoek 2010, 97, 261–273. [Google Scholar] [CrossRef]
- Troppens, D.M.; Dmitriev, R.I.; Papkovsky, D.B.; O’Gara, F.; Morrissey, J.P. Genome-wide investigation of cellular targets and mode of action of the antifungal bacterial metabolite 2, 4-diacetylphloroglucinol in Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 322–334. [Google Scholar] [CrossRef] [Green Version]
- Kwak, Y.-S.; Han, S.; Thomashow, L.S.; Rice, J.T.; Paulitz, T.C.; Kim, D.; Weller, D.M. Saccharomyces cerevisiae genome-wide mutant screen for sensitivity to 2, 4-diacetylphloroglucinol, an antibiotic produced by Pseudomonas fluorescens. Appl. Environ. Microbiol. 2011, 77, 1770–1776. [Google Scholar] [CrossRef] [Green Version]
- Julian, W.T.; Vasilchenko, A.V.; Shpindyuk, D.D.; Poshvina, D.V.; Vasilchenko, A.S. Bacterial-Derived Plant Protection Metabolite 2, 4-Diacetylphloroglucinol: Effects on Bacterial Cells at Inhibitory and Subinhibitory Concentrations. Biomolecules 2021, 11, 13. [Google Scholar] [CrossRef]
- Powers, M.J.; Sanabria-Valentín, E.; Bowers, A.A.; Shank, E.A. Inhibition of cell differentiation in Bacillus subtilis by Pseudomonas protegens. J. Bacteriol. 2015, 197, 2129–2138. [Google Scholar] [CrossRef]
- Brazelton, J.N.; Pfeufer, E.E.; Sweat, T.A.; Gardener, B.B.M.; Coenen, C. 2, 4-Diacetylphloroglucinol alters plant root development. Mol. Plant Microbe Interact. 2008, 21, 1349–1358. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Mavrodi, D.V.; Thomashow, L.S.; Weller, D.M. Differential response of wheat cultivars to Pseudomonas brassicacearum and take-all decline soil. Phytopathology 2018, 108, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Mavrodi, D.V.; Mavrodi, O.V.; Thomashow, L.S.; Weller, D.M. Exploring the Pathogenicity of Pseudomonas brassicacearum Q8r1-96 and Other Strains of the Pseudomonas fluorescens Complex on Tomato. Plant Dis. 2020, 104, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Yim, G.; Huimi Wang, H.; Davies Frs, J. Antibiotics as signalling molecules. Phil. Trans. R. Soc. B Biol. Sci. 2007, 362, 1195–1200. [Google Scholar] [CrossRef] [Green Version]
- Davies, J. Are antibiotics naturally antibiotics? J. Ind. Microbiol. Biotechnol. 2006, 33, 496–499. [Google Scholar] [CrossRef]
- Romero, D.; Traxler, M.F.; López, D.; Kolter, R. Antibiotics as signal molecules. Chem. Rev. 2011, 111, 5492–5505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, J.C.; Buchanan, A.; Vining, O.; Kidarsa, T.A.; Chang, J.H.; McPhail, K.L.; Loper, J.E. Phloroglucinol functions as an intracellular and intercellular chemical messenger influencing gene expression in Pseudomonas protegens. Environ. Microbiol. 2016, 18, 3296–3308. [Google Scholar] [CrossRef]
- Maurhofer, M.; Baehler, E.; Notz, R.; Martinez, V.; Keel, C. Cross talk between 2, 4-diacetylphloroglucinol-producing biocontrol pseudomonads on wheat roots. Appl. Environ. Microbiol. 2004, 70, 1990–1998. [Google Scholar] [CrossRef] [Green Version]
- Combes-Meynet, E.; Pothier, J.F.; Moënne-Loccoz, Y.; Prigent-Combaret, C. The Pseudomonas secondary metabolite 2, 4-diacetylphloroglucinol is a signal inducing rhizoplane expression of Azospirillum genes involved in plant-growth promotion. Mol. Plant Microbe Interact. 2011, 24, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Ramette, A.; Punjasamarnwong, P.; Zala, M.; Natsch, A.; Moënne-Loccoz, Y.; Défago, G. Cosmopolitan distribution of phlD-containing dicotyledonous crop-associated biocontrol pseudomonads of worldwide origin. FEMS Microbiol. Ecol. 2001, 37, 105–116. [Google Scholar] [CrossRef]
- Mavrodi, O.V.; McSpadden Gardener, B.B.; Mavrodi, D.V.; Bonsall, R.F.; Weller, D.M.; Thomashow, L.S. Genetic diversity of phlD from 2, 4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. Phytopathology 2001, 91, 35–43. [Google Scholar] [CrossRef] [Green Version]
- McSpadden Gardener, B.; Schroeder, K.; Kalloger, S.; Raaijmakers, J.; Thomashow, L.; Weller, D. Genotypic and Phenotypic Diversity of phlD-Containing Pseudomonas Strains Isolated from the Rhizosphere of Wheat. Appl. Environ. Microbiol. 2000, 66, 1939–1946. [Google Scholar] [CrossRef] [Green Version]
- Landa, B.B.; Mavrodi, O.V.; Schroeder, K.L.; Allende-Molar, R.; Weller, D.M. Enrichment and genotypic diversity of phlD-containing fluorescent Pseudomonas spp. in two soils after a century of wheat and flax monoculture. FEMS Microbiol. Ecol. 2006, 55, 351–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSpadden Gardener, B.B.; Mavrodi, D.V.; Thomashow, L.S.; Weller, D.M. A rapid polymerase chain reaction-based assay characterizing rhizosphere populations of 2, 4-diacetylphloroglucinol-producing bacteria. Phytopathology 2001, 91, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzola, M.; Funnell, D.L.; Raaijmakers, J. Wheat cultivar-specific selection of 2, 4-diacetylphloroglucinol-producing fluorescent Pseudomonas species from resident soil populations. Microb. Ecol. 2004, 48, 338–348. [Google Scholar] [CrossRef] [Green Version]
- McSpadden Gardener, B.B.; Gutierrez, L.J.; Joshi, R.; Edema, R.; Lutton, E. Distribution and biocontrol potential of phlD+ pseudomonads in corn and soybean fields. Phytopathology 2005, 95, 715–724. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, L.; Mavrodi, D.V.; Landa, B.B.; Thomashow, L.S.; Weller, D.M. phlD-based genetic diversity and detection of genotypes of 2, 4-diacetylphloroglucinol-producing Pseudomonas fluorescens. FEMS Microbiol. Ecol. 2006, 56, 64–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekar, J.; Prabavathy, V.R. Novel Phl-producing genotypes of finger millet rhizosphere associated pseudomonads and assessment of their functional and genetic diversity. FEMS Microbiol. Ecol. 2014, 89, 32–46. [Google Scholar] [CrossRef] [Green Version]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S. Enhanced annotations and features for comparing thousands of Pseudomonas genomes in the Pseudomonas genome database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, I.T.; Press, C.M.; Ravel, J.; Kobayashi, D.Y.; Myers, G.S.; Mavrodi, D.V.; DeBoy, R.T.; Seshadri, R.; Ren, Q.; Madupu, R. Complete genome sequence of the plant commensal Pseudomonas fluorescens Pf-5. Nat. Biotechnol. 2005, 23, 873–878. [Google Scholar] [CrossRef]
- Redondo-Nieto, M.; Barret, M.; Morrisey, J.P.; Germaine, K.; Martínez-Granero, F.; Barahona, E.; Navazo, A.; Sánchez-Contreras, M.; Moynihan, J.A.; Giddens, S.R. Genome sequence of the biocontrol strain Pseudomonas fluorescens F113. J. Bacteriol. 2012, 194, 1273–1274. [Google Scholar] [CrossRef]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W.; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K. Comparative genomics of plant-associated Pseudomonas spp.: Insights into diversity and inheritance of traits involved in multitrophic interactions. PLOS Genet. 2012, 8, e1002784. [Google Scholar] [CrossRef] [Green Version]
- Jousset, A.; Schuldes, J.; Keel, C.; Maurhofer, M.; Daniel, R.; Scheu, S.; Thuermer, A. Full-genome sequence of the plant growth-promoting bacterium Pseudomonas protegens CHA0. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Brendel, N.; Partida-Martinez, L.P.; Scherlach, K.; Hertweck, C. A cryptic PKS–NRPS gene locus in the plant commensal Pseudomonas fluorescens Pf-5 codes for the biosynthesis of an antimitotic rhizoxin complex. Org. Biomol. Chem. 2007, 5, 2211–2213. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.; Stockwell, V.O.; Henkels, M.D.; Nowak-Thompson, B.; Loper, J.E.; Gerwick, W.H. The genomisotopic approach: A systematic method to isolate products of orphan biosynthetic gene clusters. Chem. Biol. 2007, 14, 53–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezzonico, F.; Zala, M.; Keel, C.; Duffy, B.; Moënne-Loccoz, Y.; Défago, G. Is the ability of biocontrol fluorescent pseudomonads to produce the antifungal metabolite 2, 4-diacetylphloroglucinol really synonymous with higher plant protection? New Phytol. 2007, 173, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Laville, J.; Blumer, C.; Von Schroetter, C.; Gaia, V.; Défago, G.; Keel, C.; Haas, D. Characterization of the hcnABC gene cluster encoding hydrogen cyanide synthase and anaerobic regulation by ANR in the strictly aerobic biocontrol agent Pseudomonas fluorescens CHA0. J. Bacteriol. 1998, 180, 3187–3196. [Google Scholar] [CrossRef] [Green Version]
- Voisard, C.; Keel, C.; Haas, D.; Dèfago, G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. Embo J. 1989, 8, 351–358. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Weller, D.M. Exploiting Genotypic Diversity of 2, 4-Diacetylphloroglucinol-Producing Pseudomonas spp.: Characterization of Superior Root-Colonizing P. fluorescens Strain Q8r1-96. Appl. Environ. Microbiol. 2001, 67, 2545–2554. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, L.; Landa, B.B.; Weller, D.M. Host crop affects rhizosphere colonization and competitiveness of 2, 4-diacetylphloroglucinol-producing Pseudomonas fluorescens. Phytopathology 2006, 96, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, W.; Rubin-Pitel, S.B.; Shao, Z.; Zhao, H. Improving cellular malonyl-CoA level in Escherichia coli via metabolic engineering. Metab. Eng. 2009, 11, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Jiang, X.; Zhang, R.; Xian, M. Improved phloroglucinol production by metabolically engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2011, 91, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.; Lee, J.-K.; Zhao, H. Directed evolution of phloroglucinol synthase PhlD with increased stability for phloroglucinol production. Appl. Microbiol. Biotechnol. 2013, 97, 5861–5867. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biessy, A.; Filion, M. Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, and Functions. Metabolites 2021, 11, 182. https://doi.org/10.3390/metabo11030182
Biessy A, Filion M. Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, and Functions. Metabolites. 2021; 11(3):182. https://doi.org/10.3390/metabo11030182
Chicago/Turabian StyleBiessy, Adrien, and Martin Filion. 2021. "Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, and Functions" Metabolites 11, no. 3: 182. https://doi.org/10.3390/metabo11030182
APA StyleBiessy, A., & Filion, M. (2021). Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, and Functions. Metabolites, 11(3), 182. https://doi.org/10.3390/metabo11030182