
Metabolic Mechanism of Plant Defense against Rice Blast Induced by Probenazole

 , and
, and
Abstract
:1. Introduction
2. Results
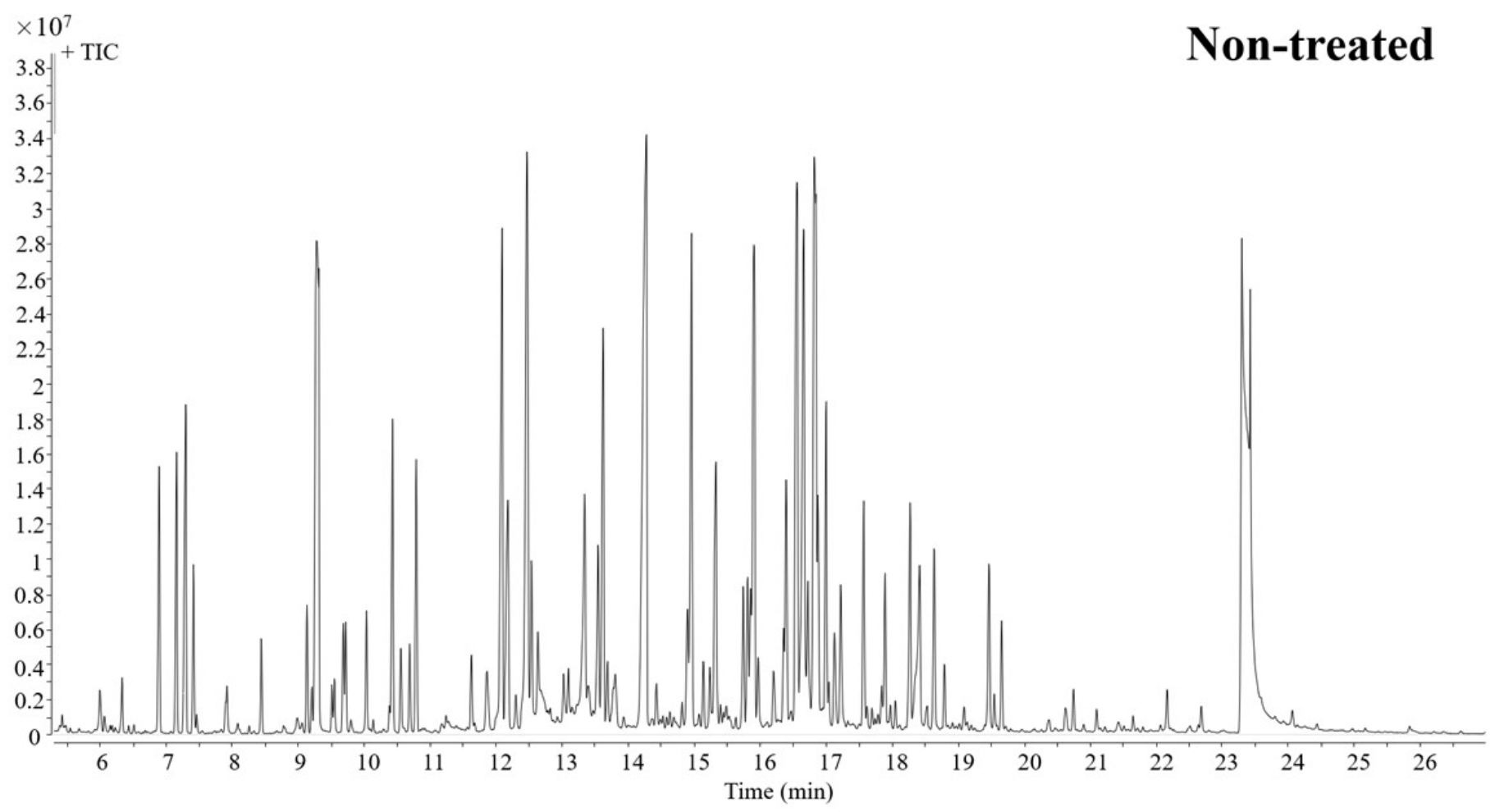
2.1. Effect of Probenazole on the Metabolome of Rice Seedlings
2.2. Effect of Probenazole on Differential Metabolites of Rice Seedlings
2.3. Effect of Probenazole on Metabolic Pathways of Rice Seedlings
3. Discussion
4. Materials and Methods
4.1. Chemical, Plant and Fungal Strain
4.2. Pretreatment
4.3. Gas Chromatography and Mass Spectrometry (GC-MS) Analysis
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oerke, E.C.; Dehne, H.W. Safeguarding Production-Losses in major crops and the role of crop protection. Crop. Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
- Yoshioka, K.; Nakashita, H.; Klessig, D.F.; Yamaguchi, I. Probenazole induces systemic acquired resistance in Arabidopsis with a novel type of action. Plant. J. 2001, 25, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Tada, Y.; Yokozeki, Y.; Akagi, H.; Hayashi, N.; Fujimura, T.; Ichikawa, N. Chemical induction of disease resistance in rice is correlated with the expression of a gene encoding a nucleotide binding site and leucine-rich repeats. Plant. Mol. Biol. 1999, 40, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Enyedi, A.J.; Yalpani, N.; Silverman, P.; Raskin, I. Localization, conjugation, and function of salicylic acid in tobacco during the hypersensitive reaction to tobacco mosaic virus. Proc. Natl. Acad. Sci. USA 1992, 89, 2480–2484. [Google Scholar] [CrossRef] [Green Version]
- Malamy, J.; Carr, J.P.; Klessig, D.F.; Raskin, I. Salicylic acid: A likely endogenous signal in the resistance response of tobacco to viral infection. Science 1990, 250, 1002–1004. [Google Scholar] [CrossRef] [Green Version]
- M´etraux, J.P.; Signer, H.; Ryals, J.; Ward, E.; Wyss-Benz, M. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science 1990, 250, 1004–1006. [Google Scholar] [CrossRef] [Green Version]
- Yalpani, N.; Silverman, P.; Wilson, T.M.A.; Kleier, D.A.; Raskin, I. Salicylic acid is a systemic signal and an inducer of pathogenesis-related proteins in virus-infected tobacco. Plant. Cell 1991, 3, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Silverman, P.; Sesker, M.; Kanter, D.; Schweizer, P.; Metraux, J.P.; Raskin, I. Salicylic acid in rice. Plant. Physiol. 1995, 108, 633–639. [Google Scholar] [CrossRef] [Green Version]
- Nakashita, H.; Yoshioka, K.; Yasuda, M.; Nitta, T.; Arai, Y.; Yoshida, S.; Yamaguchi, I. Probenazole induces systemic acquired resistance in tobacco through salicylic acid accumulation. Physiol. Mol. Plant. Pathol. 2002, 61, 197–203. [Google Scholar] [CrossRef]
- Watanabe, T.; Sekizawa, Y.; Shimura, M.; Suzuki, Y.; Matsumoto, K.; Iwata, M.; Mase, S. Effects of probenazole (Oryzemate) on rice plants with reference to controlling rice blast. J. Pestic. Sci. 1979, 4, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Shimura, M.; Iwata, M.; Tashiro, N.; Sekizawa, Y.; Suzuki, Y.; Mase, S.; Watanabe, T. Anti-conidial germination factors induced in the presence of probenazole and properties off our active substances. Agric. Biol. Chem. 1981, 45, 1431–1435. [Google Scholar]
- Yamaguchi, I. Activators for systemic acquired resistance. In Fungicidal Activity; Hutson, H., Miyamoto, J., Eds.; John Wiley & Sons Ltd.: London, UK, 1998; pp. 193–219. [Google Scholar]
- Ishiga, T.; Sakata, N.; Ugajin, T.; Ishiga, Y. Acibenzolar-S-methyl and probenazole activate stomatal based defense at different times to control bacterial blight of cabbage. J. Gen. Plant. Pathol. 2021, 87, 30–34. [Google Scholar] [CrossRef]
- Iwai, T.; Seo, S.; Mitsuhara, I.; Ohashi, Y. Probenazole-induced accumulation of salicylic acid confers resistance to Magnaporthe grisea in adult rice plants. Plant. Cell Physiol. 2007, 48, 915–924. [Google Scholar] [CrossRef] [Green Version]
- Phuong, L.T.; Zhao, L.; Fitrianti, A.N.; Matsui, H.; Noutoshi, Y.; Yamamoto, M.; Ichinose, Y.; Shiraishi, T.; Toyoda, K. The plant activator saccharin induces resistance to wheat powdery mildew by activating multiple defense-related genes. J. Gen. Plant. Pathol. 2020, 86, 107–113. [Google Scholar] [CrossRef]
- Heuberger, A.L.; Robison, F.M.; Lyons, S.M.A.; Broeckling, C.D.; Prenni, J.E. Evaluating plant immunity using mass spectrometry-based metabolomics workflows. Front. Plant. Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.Z.; Li, X.M.; Han, R.Q.; Cheng, X.K.; Xu, Y.J.; Huang, Z.Q.; Xu, B.; Liu, P.F.; Liu, X.L. Research of probenazole and sedaxane combined granule and the control efficacy on rice blast and sheath blight at seedling stage. Chin. J. Pesticide Sci. 2020, 22, 362–369. [Google Scholar] [CrossRef]
- Peluffo, L.; Lia, V.; Troglia, C.; Maringolo, C.; Norma, P.; Escande, A.; Esteban Hopp, H.; Lytovchenko, A.; Fernie, A.R.; Heinz, R.; et al. Metabolic profiles of sunflower genotypes with contrasting response to Sclerotinia sclerotiorum infection. Phytochemistry 2010, 71, 70–80. [Google Scholar] [CrossRef]
- Zhang, C.G.; Jing, H.M.; Zheng, H.L.; Zhao, Z.Q. Progress in Plant Resistance Induced by Salicylic Acid. Life Sci. Res. 2001, 5, 185–189. [Google Scholar] [CrossRef]
- Umemura, K.; Satou, J.; Iwata, M.; Uozumi, N.; Koga, J.; Kawano, T. Contribution of salicylic acid glucosyltransferase, OsSGT1, to chemically induced disease resistance in rice plants. Plant J. 2009, 57, 463–472. [Google Scholar] [CrossRef]
- Schwacke, R.; Grallath, S.; Breitkreuz, K.E.; Stransky, E.; Stransky, H.; Frommer, W.B.; Rentsch, D. LeProT1, a transporter for proline, glycine betaine, and γ-amino butyric acid in tomato pollen. Plant Cell 1999, 11, 377–391. [Google Scholar] [CrossRef] [Green Version]
- Erland, L.A.E.; Turi, C.E.; Saxena, P.K. Serotonin: An ancient molecule and an important regulator of plant processes. Biotechnol. Adv. 2016, 34, 1347–1361. [Google Scholar] [CrossRef]
- Hückelhoven, R. Cell wall-associated mechanisms of disease resistance and susceptibility. Annu. Rev. Phytopathol. 2008, 45, 101–127. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Xu, H.; Yi, H.; Yang, L.; Kong, Z.; Zhang, L.; Zhang, L.; Xue, S.; Ma, Z. Resistance to hemi-biotrophic F. graminearum infection is associated with coordinated and ordered expression of diverse defense signaling pathways. PLoS ONE 2011, 6, e19008. [Google Scholar] [CrossRef] [Green Version]
- Phuong, L.T.; Nur, A.; Mai, F.; Luan, T.; Matsui, H.; Noutoshi, Y. Antagonism between SA- and JA- signaling conditioned by saccharin in Arabidopsis thaliana renders resistance to a specific pathogen. J. Gen. Plant. Pathol. 2020, 86, 86–99. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annual Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Halim, V.A.; Vess, A.; Scheel, D.; Rosahl, S. The role of salicylic acid and jasmonic acid in pathogen defence. Plant. Biol. 2006, 8, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, Q.; Xie, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Abscisic acid and jasmonic acid are involved in drought priming-induced tolerance to drought in wheat. Crop. J. 2021, 9, 120–132. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The Crosstalks Between Jasmonic Acid and Other Plant Hormone Signaling Highlight the Involvement of Jasmonic Acid as a Core Component in Plant Response to Biotic and Abiotic Stresses. Front. Plant. Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Farmer, E.E.; Ryan, C.A. Octadecanoid precursors of jasmonic acid activate the synthesis of wound-inducible proteinase inhibitors. Plant Cell 1992, 4, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.; Beckmann, M.; Zubair, H.; Enot, D.P.; Caracuel-Rios, Z.; Overy, D.P.; Snowdon, S.; Talbot, N.J.; Draper, J. Metabolomic analysis reveals a common pattern of metabolic re-programming during invasion of three host plant species by Magnaporthe grisea. Plant J. 2009, 59, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Thines, E.; Weber, R.W.S.; Talbot, N.J. MAP kinase and protein kinase A-dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea. Plant Cell 2000, 12, 1703–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauzin, A.S.; Sulzenbacher, G.; Lafond, M.; Desseaux, V.; Reca, I.B.; Perrier, J.; Bellincampi, D.; Fourquet, P.; Lévêque, C.; Giardina, T. Functional characterization of a vacuolar invertase from Solanum lycopersicum: Post-translational regulation by N-glycosylation and a proteinaceous inhibitor. Biochimie 2014, 101, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Swarbrick, P.J.; Schulze-Lefert, P.; Scholes, J.D. Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant Cell Environ. 2006, 29, 1061–1076. [Google Scholar] [CrossRef]
- Shi, K.; Fu, L.J.; Zhang, S.; Li, X.; Liao, Y.W.K.; Xia, X.J.; Zhou, Y.H.; Wang, R.Q.; Chen, Z.X.; Yu, J.Q. Flexible change and cooperation between mitochondrial electron transport and cytosolic glycolysis as the basis for chilling tolerance in tomato plants. Planta 2013, 237, 589–601. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Vorwerk, S.; Somerville, S.; Somerville, C. The role of plant cell wall polysaccharide composition in disease resistance. Trends Plant Sci. 2004, 9, 203–209. [Google Scholar] [CrossRef]
- Brisson, L.F.; Tenhaken, R.; Lamb, C. Function of oxidative cross-linking of cell wall structural proteins in plant disease resistance. Plant Cell 1994, 6, 1703–1712. [Google Scholar] [CrossRef] [Green Version]
- Hardham, A.R.; Jones, D.A.; Takemoto, D. Cytoskeleton and cell wall function in penetration resistance. Curr. Opin. Plant Biol. 2007, 10, 342–348. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Y.; Gao, C. The ethylene response factor (ERF) genes from Tamarix hispida respond to salt, drought and ABA treatment. Trees 2014, 28, 317–327. [Google Scholar] [CrossRef]
- Liu, W.; Nju, K.; Wu, T.; Yang, Y.; Zhang, X.; Xu, X.; Wang, Y.; Han, Z. The ethylene response factor AtERF4 negatively regulates the iron deficiency response in Arabidopsis thaliana. PLoS ONE 2017, 12, e0186580. [Google Scholar]
- Liu, W.; Wu, T.; Li, Q.; Zhang, X.; Xu, X.; Li, T.; Han, Z.; Wang, Y. An ethylene response factor (MxERF4) functions as a repressor of Fe acquisition in Malus xiaojinensis. Sci. Rep. 2018, 8, 1068. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Zhang, L.; Rao, S.; Zhu, X.; Ye, L.; Chen, W.; Li, X. The relationship between the expression of ethylene-related genes and papaya fruit ripening disorder caused by chilling injury. PLoS ONE 2014, 9, e116002. [Google Scholar] [CrossRef]
- Yin, X.R.; Shi, Y.N.; Min, T.; Luo, Z.R.; Yao, Y.C.; Xu, Q.; Ferguson, I.; Chen, K.-S. Expression of ethylene response genes during persimmon fruit astringency removal. Planta 2012, 235, 895–906. [Google Scholar] [CrossRef]
- Tomotsugu, K.; Haruka, N.; Nobutaka, M.; Masaru, O.; Sakihito, K.; Masaru, O.T.; Fumihiko, S. A regulatory cascade involving class II ETHYLENE RESPONSE FACTOR transcriptional repressors operates in the progression of leaf senescence. Plant Physiol. 2013, 162, 991–1005. [Google Scholar]
- Van Loon, L.C.; Geraats, B.P.J.; Linthorst, H.J.M. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci. 2006, 11, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Raz, V.; Fluhr, R. Calcium requirement for ethylene- dependent responses. Plant. Cell 1992, 4, 1123–1130. [Google Scholar] [CrossRef]
- Lawton, K.A.; Potter, S.L.; Uknes, S.; Ryals, J. Acquired resistance signal transduction in Arabidopsis is ethylene independent. Plant Cell 1994, 6, 581–588. [Google Scholar] [CrossRef]
- Costa, F.; Peace, C.P.; Stella, S.; Serra, S.; Musacchi, S.; Bazzani, M.; Sansavini, S.; Van de Weg, W.E. QTL dynamics for fruit firmness and softening around an ethylene-dependent polygalacturonase gene in apple (Malus x domestica Borkh.). J. Exp. Bot. 2010, 61, 3029–3039. [Google Scholar] [CrossRef] [Green Version]
- Leslie, C.A.; Romani, R.J. Inhibition of Ethylene Biosynthesis by Salicylic Acid. Plant Physiol. 1988, 88, 833–837. [Google Scholar] [CrossRef] [Green Version]
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (GABA) and plant responses to stress. CRC. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Pérez, M.; Invers, O.; Ruiz, J.M.; Frederiksen, M.S.; Holmer, M. Physiological responses of the seagrass Posidonia oceanica to elevated organic matter content in sediments: An experimental assessment. J. Exp. Mar. Bio. Ecol. 2007, 344, 149–160. [Google Scholar] [CrossRef]
- Naylor, A.W.; Tolbert, N.E. Glutamic Acid Metabolism in Green and Etiolated Barley Leaves. Physiol. Plant. 1956, 9, 220–229. [Google Scholar] [CrossRef]
- Fischer, W.N.; André, B.; Rentsch, D.; Krolkiewicz, S.; Tegeder, M.; Breitkreuz, K.; Frommer, W.B. Amino acid transport in plants. Trends Plant. Sci. 1998, 3, 188–195. [Google Scholar] [CrossRef]
- Dai, T.; Chang, X.; Hu, Z.; Liang, L.; Sun, M.; Liu, P.; Liu, X. Untargeted Metabolomics Based on GC-MS and Chemometrics: A New Tool for the Early Diagnosis of Strawberry Anthracnose Caused by Colletotrichum theobromicola. Plant. Dis. 2019, 103, 2541–2547. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Match Status a | P b | Holm P c | Impact d |
|---|---|---|---|---|
| Ascorbate and aldarate metabolism | 1/4 | 0.016 | 0.196 | 0.500 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | 1/4 | 0.097 | 0.582 | 0.500 |
| Alanine, aspartate and glutamate metabolism | 3/14 | 0.000 | 0.002 | 0.424 |
| Phenylalanine metabolism | 1/10 | 0.226 | 0.945 | 0.357 |
| Glycine, serine and threonine metabolism | 1/11 | 0.048 | 0.443 | 0.270 |
| Arginine biosynthesis | 3/14 | 0.004 | 0.093 | 0.228 |
| Glyoxylate and dicarboxylate metabolism | 1/8 | 0.007 | 0.125 | 0.185 |
| Inositol phosphate metabolism | 1/15 | 0.173 | 0.807 | 0.129 |
| Pentose and glucuronate interconversions | 1/18 | 0.370 | 1.000 | 0.125 |
| Starch and sucrose metabolism | 1/9 | 0.073 | 0.559 | 0.123 |
| Tryptophan metabolism | 1/41 | 0.653 | 1.000 | 0.105 |
| Arginine and proline metabolism | 1/19 | 0.248 | 0.945 | 0.102 |
| Glycerolipid metabolism | 1/16 | 0.336 | 1.000 | 0.093 |
| Galactose metabolism | 1/9 | 0.028 | 0.298 | 0.092 |
| Glutathione metabolism | 1/28 | 0.513 | 1.000 | 0.089 |
| Citrate cycle (TCA cycle) | 1/10 | 0.088 | 0.570 | 0.077 |
| Glycolysis / Gluconeogenesis | 1/13 | 0.137 | 0.702 | 0.072 |
| Fructose and mannose metabolism | 1/20 | 0.401 | 1.000 | 0.051 |
| Pentose phosphate pathway | 3/22 | 0.016 | 0.196 | 0.047 |
| Phosphatidylinositol signaling system | 1/28 | 0.513 | 1.000 | 0.037 |
| Butanoate metabolism | 2/15 | 0.053 | 0.443 | 0.032 |
| Pyruvate metabolism | 1/22 | 0.431 | 1.000 | 0.031 |
| Fatty acid biosynthesis | 1/47 | 0.704 | 1.000 | 0.015 |
| Glycerophospholipid metabolism | 1/36 | 0.605 | 1.000 | 0.013 |
| Primary bile acid biosynthesis | 1/46 | 0.696 | 1.000 | 0.008 |
| Tyrosine metabolism | 1/42 | 0.662 | 1.000 | 0.007 |
| Amino sugar and nucleotide sugar metabolism | 2/37 | 0.238 | 0.945 | 0.000 |
| Aminoacyl-tRNA biosynthesis | 5/24 | 0.000 | 0.000 | 0.000 |
| beta-Alanine metabolism | 1/21 | 0.416 | 1.000 | 0.000 |
| Biosynthesis of unsaturated fatty acids | 1/18 | 0.229 | 0.945 | 0.000 |
| D-Glutamine and D-glutamate metabolism | 1/6 | 0.142 | 0.702 | 0.000 |
| Fatty acid degradation | 1/39 | 0.634 | 1.000 | 0.000 |
| Fatty acid elongation | 1/39 | 0.634 | 1.000 | 0.000 |
| Histidine metabolism | 1/16 | 0.336 | 1.000 | 0.000 |
| Lysine degradation | 1/25 | 0.474 | 1.000 | 0.000 |
| Nicotinate and nicotinamide metabolism | 1/15 | 0.319 | 1.000 | 0.000 |
| Nitrogen metabolism | 1/6 | 0.142 | 0.702 | 0.000 |
| Pantothenate and CoA biosynthesis | 2/19 | 0.081 | 0.564 | 0.000 |
| Porphyrin and chlorophyll metabolism | 1/30 | 0.538 | 1.000 | 0.000 |
| Propanoate metabolism | 1/23 | 0.446 | 1.000 | 0.000 |
| Purine metabolism | 1/65 | 0.816 | 1.000 | 0.000 |
| Pyrimidine metabolism | 1/39 | 0.634 | 1.000 | 0.000 |
| Selenocompound metabolism | 1/20 | 0.401 | 1.000 | 0.000 |
| Valine, leucine and isoleucine biosynthesis | 3/8 | 0.001 | 0.021 | 0.000 |
| Valine, leucine and isoleucine degradation | 1/20 | 0.267 | 0.973 | 0.000 |
| Pesticide | Dispose Group | Gram/m2 |
|---|---|---|
| None | A | 0 |
| 16% probenazole granules | B | 75.00 |
| 16% probenazole granules | C | 112.50 |
| 16% probenazole granules | D | 150.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Wang, G.; Zhang, B.; Dai, T.; Gu, A.; Li, X.; Cheng, X.; Liu, P.; Hao, J.; Liu, X. Metabolic Mechanism of Plant Defense against Rice Blast Induced by Probenazole. Metabolites 2021, 11, 246. https://doi.org/10.3390/metabo11040246
Wu Z, Wang G, Zhang B, Dai T, Gu A, Li X, Cheng X, Liu P, Hao J, Liu X. Metabolic Mechanism of Plant Defense against Rice Blast Induced by Probenazole. Metabolites. 2021; 11(4):246. https://doi.org/10.3390/metabo11040246
Chicago/Turabian StyleWu, Zhaochen, Guozhen Wang, Borui Zhang, Tan Dai, Anyu Gu, Xiaolin Li, Xingkai Cheng, Pengfei Liu, Jianjun Hao, and Xili Liu. 2021. "Metabolic Mechanism of Plant Defense against Rice Blast Induced by Probenazole" Metabolites 11, no. 4: 246. https://doi.org/10.3390/metabo11040246
APA StyleWu, Z., Wang, G., Zhang, B., Dai, T., Gu, A., Li, X., Cheng, X., Liu, P., Hao, J., & Liu, X. (2021). Metabolic Mechanism of Plant Defense against Rice Blast Induced by Probenazole. Metabolites, 11(4), 246. https://doi.org/10.3390/metabo11040246