
Oral Supplementation of an Alkylglycerol Mix Comprising Different Alkyl Chains Effectively Modulates Multiple Endogenous Plasmalogen Species in Mice

 , ,
, , 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
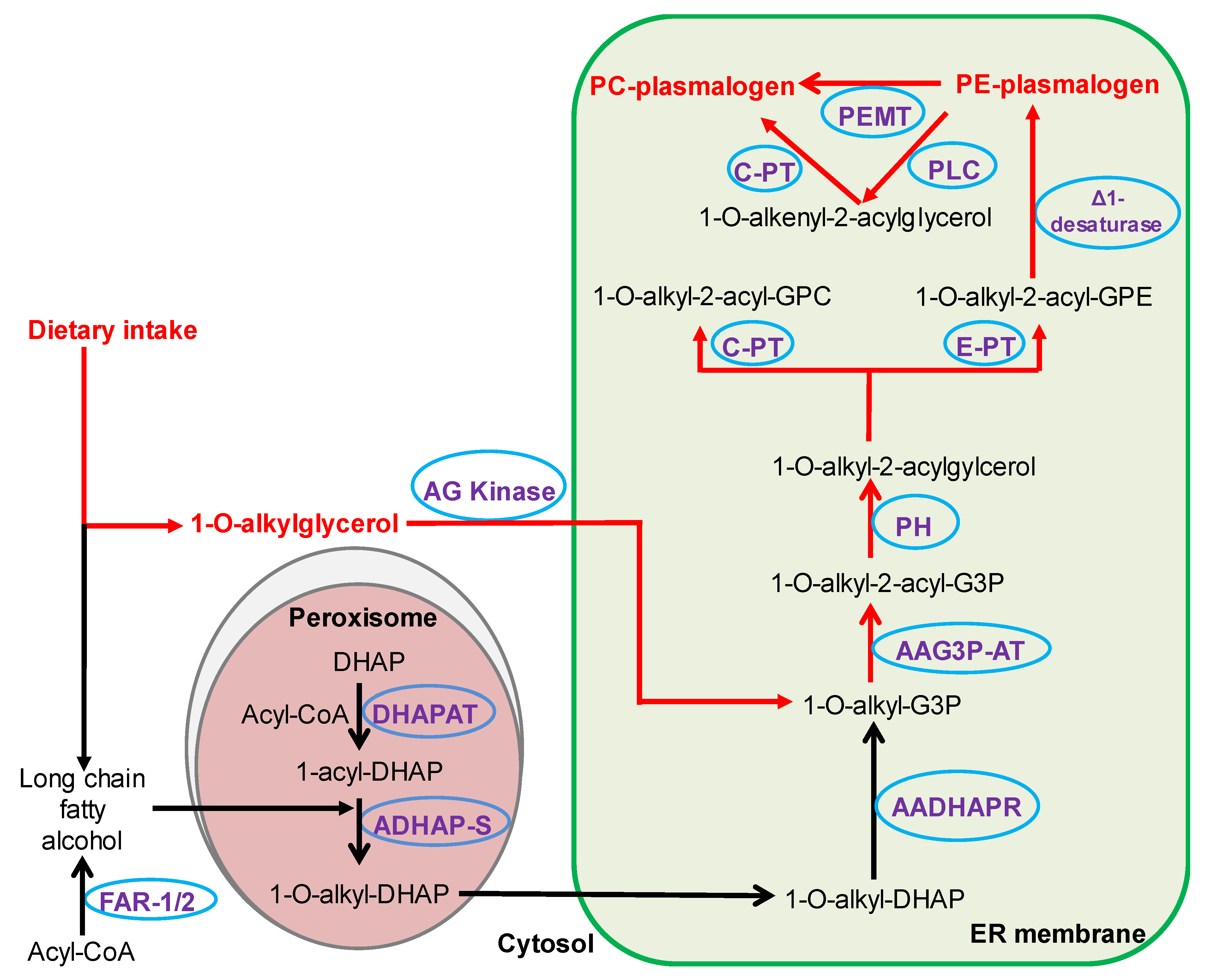
2.1. Effects of AKG Mix Treatment on Endogenous Alkyl Ether Phospholipids in Mice
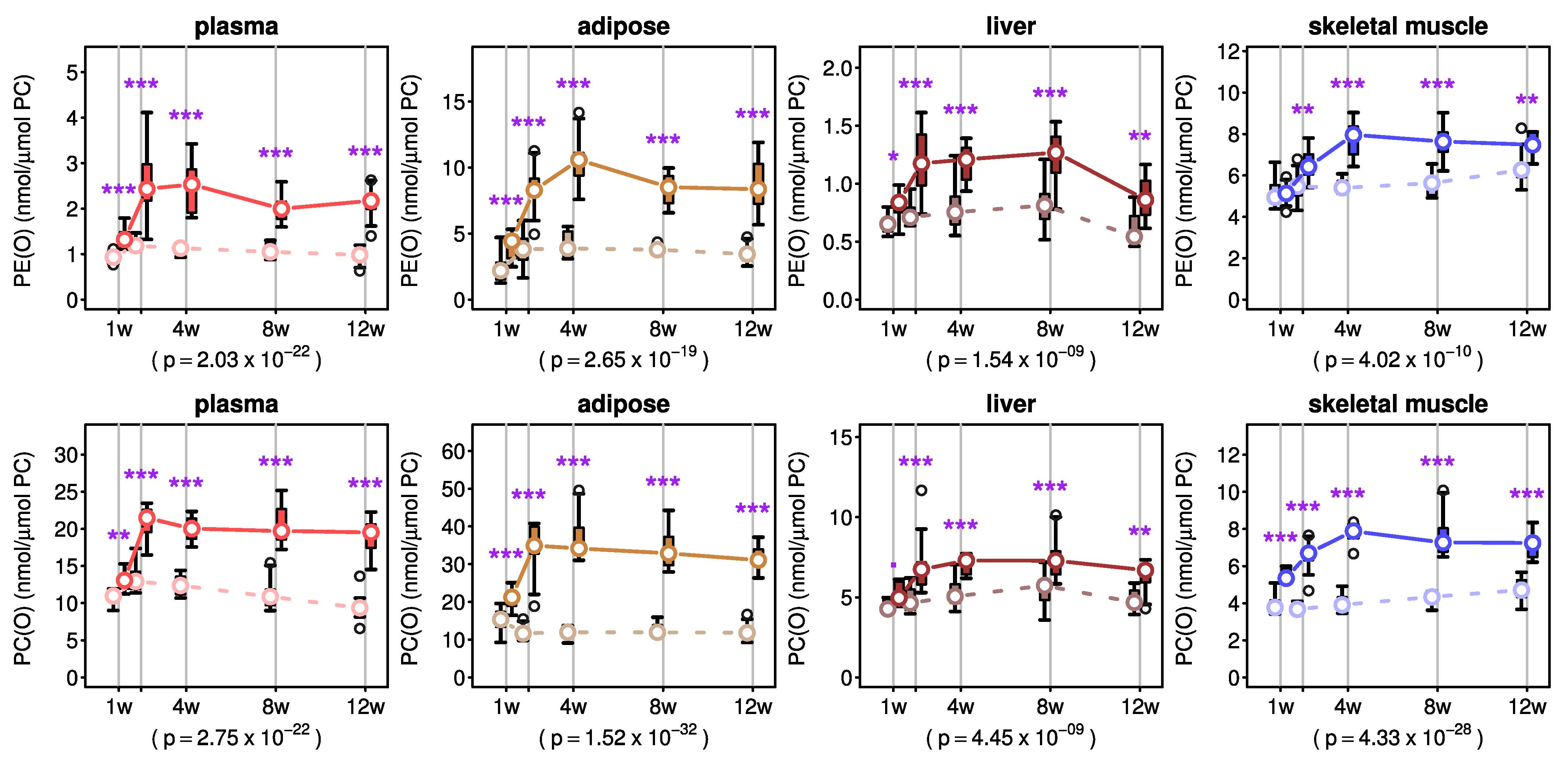
2.2. Effects of AKG Mix Treatment on Endogenous Plasmalogens in Mice
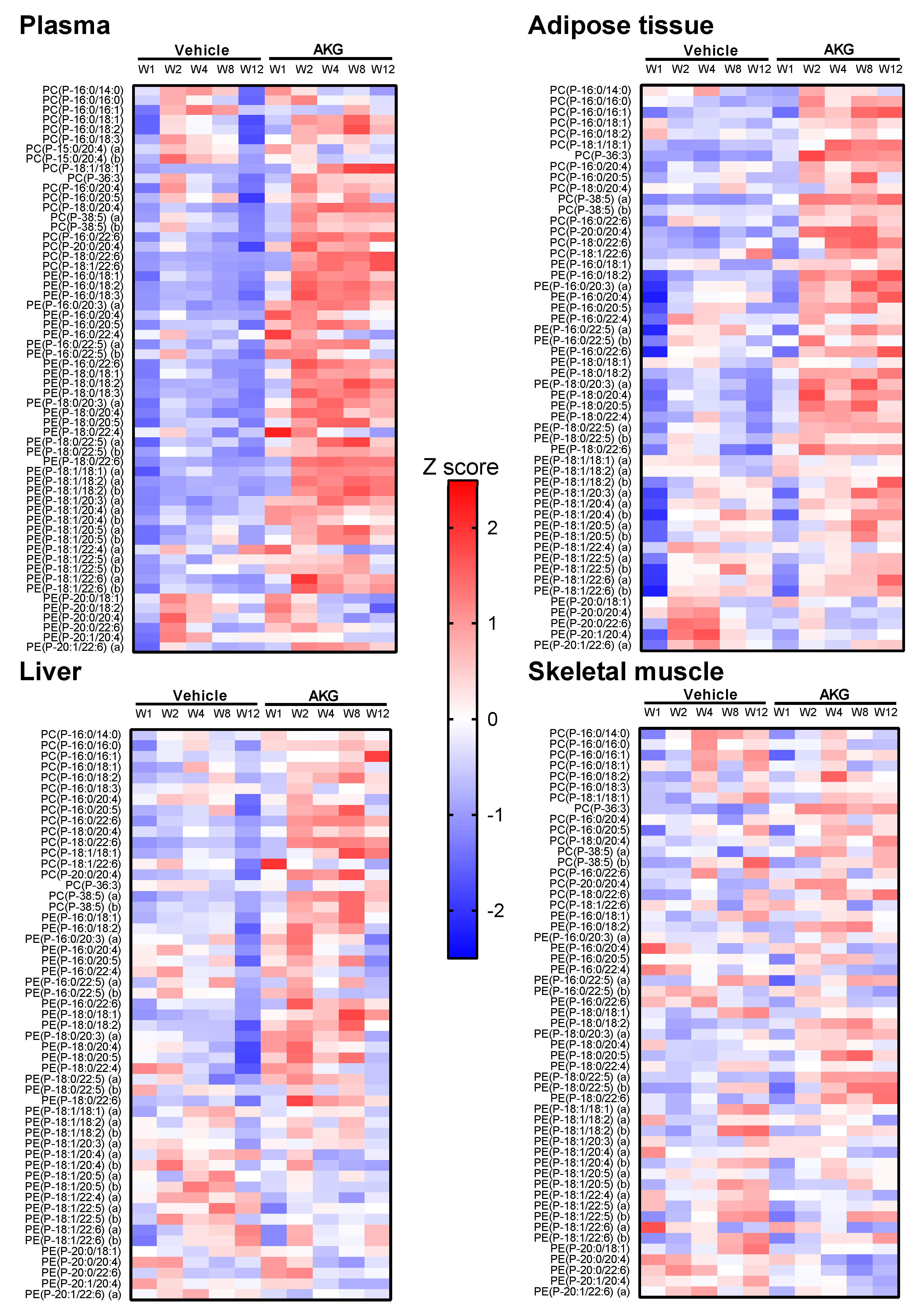
2.3. Effects of AKG Mix Treatment on Different Plasmalogen Species in Mice
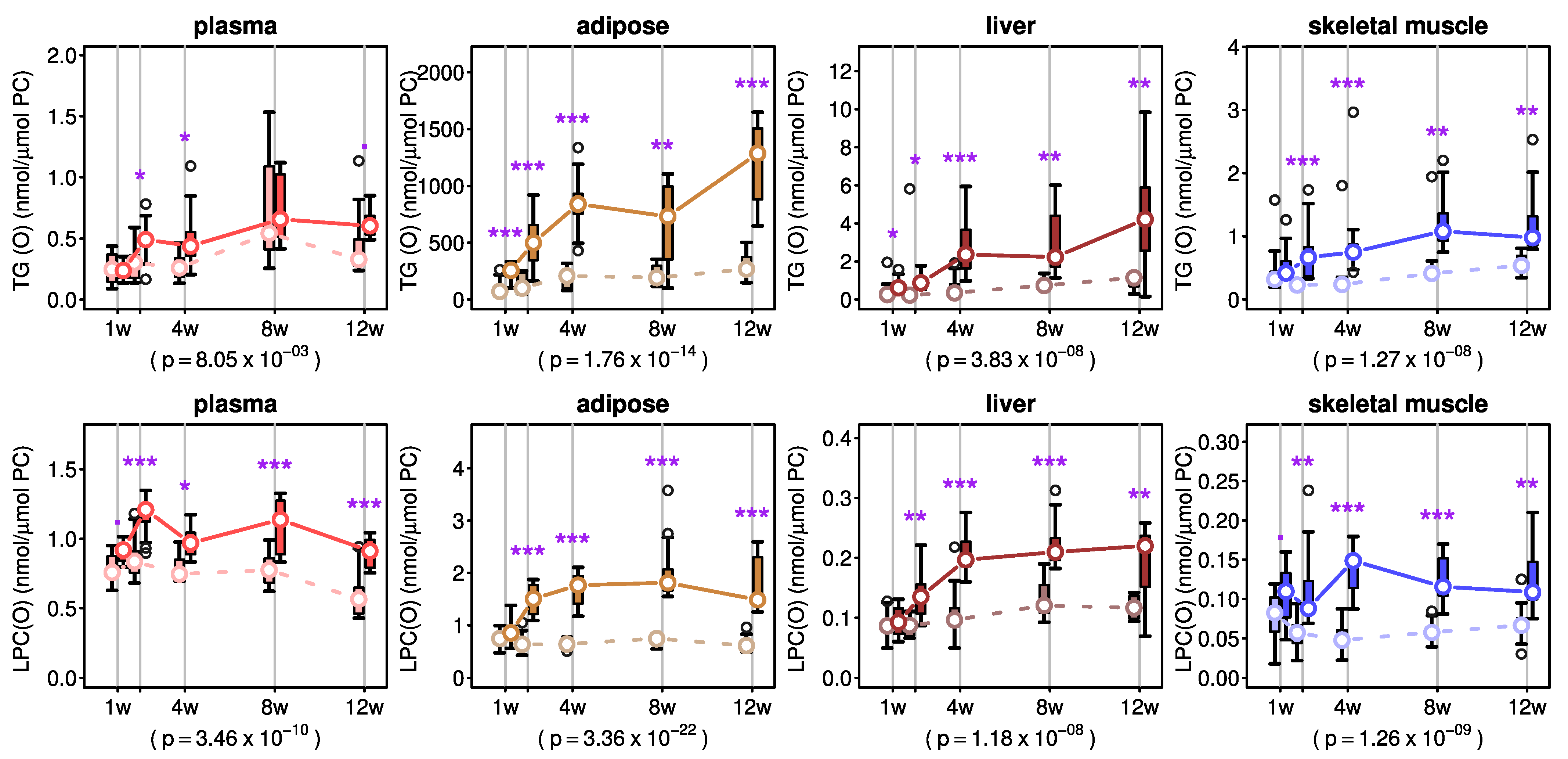
2.4. Effects of AKG Mix Treatment on Ether Triacylglycerols and Lyso-Ether Phospholipids in Mice
2.5. Effects of Alkylglycerol Mix Treatment on Sphingolipids, Lysophospholipids, Phospholipids and Neutral Lipids in Mice
3. Discussion
4. Materials and Methods
4.1. Preparation of the AKG Mix
4.2. Animal Experimentation
4.3. Tissue Homogenisation
4.4. Lipid Extraction
4.5. Liquid Chromatography and Electrospray Ionisation Tandem Mass Spectrometry
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagan, N.; Zoeller, R.A. Plasmalogens: Biosynthesis and functions. Prog. Lipid Res. 2001, 40, 199–229. [Google Scholar] [CrossRef]
- Lee, T.C. Biosynthesis and possible biological functions of plasmalogens. Biochim. Biophys. Acta 1998, 1394, 129–145. [Google Scholar] [CrossRef]
- Jurgens, G.; Fell, A.; Ledinski, G.; Chen, Q.; Paltauf, F. Delay of copper-catalyzed oxidation of low density lipoprotein by in vitro enrichment with choline or ethanolamine plasmalogens. Chem. Phys. Lipids 1995, 77, 25–31. [Google Scholar] [CrossRef]
- Maeba, R.; Ueta, N. Ethanolamine plasmalogens prevent the oxidation of cholesterol by reducing the oxidizability of cholesterol in phospholipid bilayers. J. Lipid Res. 2003, 44, 164–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honsho, M.; Abe, Y.; Fujiki, Y. Dysregulation of Plasmalogen Homeostasis Impairs Cholesterol Biosynthesis. J. Biol. Chem. 2015, 290, 28822–28833. [Google Scholar] [CrossRef] [Green Version]
- Munn, N.J.; Arnio, E.; Liu, D.; Zoeller, R.A.; Liscum, L. Deficiency in ethanolamine plasmalogen leads to altered cholesterol transport. J. Lipid Res. 2003, 44, 182–192. [Google Scholar] [CrossRef] [Green Version]
- Mandel, H.; Sharf, R.; Berant, M.; Wanders, R.J.; Vreken, P.; Aviram, M. Plasmalogen phospholipids are involved in HDL-mediated cholesterol efflux: Insights from investigations with plasmalogen-deficient cells. Biochem. Biophys. Res. Commun. 1998, 250, 369–373. [Google Scholar] [CrossRef]
- Rubio, J.M.; Astudillo, A.M.; Casas, J.; Balboa, M.A.; Balsinde, J. Regulation of Phagocytosis in Macrophages by Membrane Ethanolamine Plasmalogens. Front. Immunol. 2018, 9, 1723. [Google Scholar] [CrossRef] [PubMed]
- Ifuku, M.; Katafuchi, T.; Mawatari, S.; Noda, M.; Miake, K.; Sugiyama, M.; Fujino, T. Anti-inflammatory/anti-amyloidogenic effects of plasmalogens in lipopolysaccharide-induced neuroinflammation in adult mice. J. Neuroinflamm. 2012, 9, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguma, E.; Shinji Yamashita, S.; Kumagai, K.; Otoki, Y.; Yamamoto, A.; Eitsuka, T.; Nakagawa, K.; Miyazawa, T.; Kinoshita, M. Ethanolamine plasmalogen suppresses apoptosis in human intestinal tract cells in vitro by attenuating induced inflammatory stress. ACS Omega 2021, 6, 3140–3148. [Google Scholar] [CrossRef]
- Park, H.; He, A.; Tan, M.; Johnson, J.M.; Dean, J.M.; Pietka, T.A.; Chen, Y.; Zhang, X.; Hsu, F.F.; Razani, B.; et al. Peroxisome-derived lipids regulate adipose thermogenesis by mediating cold-induced mitochondrial fission. J. Clin. Investig. 2019, 129, 694–711. [Google Scholar] [CrossRef] [Green Version]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, S.J.; Dodt, G.; Raymond, G.V.; Braverman, N.E.; Moser, A.B.; Moser, H.W. Peroxisome biogenesis disorders. Biochim. Biophys. Acta 2006, 1763, 1733–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodenowe, D.B.; Cook, L.L.; Liu, J.; Lu, Y.; Jayasinghe, D.A.; Ahiahonu, P.W.; Heath, D.; Yamazaki, Y.; Flax, J.; Krenitsky, K.F.; et al. Peripheral ethanolamine plasmalogen deficiency: A logical causative factor in Alzheimer’s disease and dementia. J. Lipid Res. 2007, 48, 2485–2498. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, V.; Goodenowe, D.B. Plasmalogen deficiency and neuropathology in Alzheimer’s disease: Causation or coincidence? Alzheimer Dement. Transl. Res. Clin. Interv. 2019, 5, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Weir, J.M.; Wong, G.; Barlow, C.K.; Greeve, M.A.; Kowalczyk, A.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Jowett, J.B.; Shaw, J.; et al. Plasma lipid profiling in a large population-based cohort. J. Lipid Res. 2013, 54, 2898–2908. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.J.; Wong, G.; Barlow, C.K.; Weir, J.M.; Greeve, M.A.; MacIntosh, G.L.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Kowalczyk, A.; et al. Plasma lipid profiling shows similar associations with prediabetes and type 2 diabetes. PLoS ONE 2013, 8, e74341. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.J.; Wong, G.; Tsorotes, D.; Barlow, C.K.; Weir, J.M.; Christopher, M.J.; MacIntosh, G.L.; Goudey, B.; Stern, L.; Kowalczyk, A.; et al. Plasma lipidomic analysis of stable and unstable coronary artery disease. Arter. Thromb Vasc. Biol. 2011, 31, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Snyder, F. Alkylglycerol phosphotransferase. In Methods in Enzymology; Dennis, E.A., Vance, E.E., Eds.; Academic Press: Cambridge, MA, USA, 1992; Volume 209, pp. 211–215. [Google Scholar]
- Watschinger, K.; Werner, E.R. Orphan enzymes in ether lipid metabolism. Biochimie 2013, 95, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Lancaster, G.I.; Meikle, P.J. Plasmalogens: A potential therapeutic target for neurodegenerative and cardiometabolic disease. Prog. Lipid Res. 2019, 74, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Blank, M.L.; Snyder, F. Plasmanylethanolamine Δ1-desaturase. In Methods in Enzymology; Dennis, E.A., Vance, E.E., Eds.; Academic Press: Cambridge, MA, USA, 1992; Volume 209, pp. 390–396. [Google Scholar]
- Gallego-Garcia, A.; Monera-Girona, A.J.; Pajares-Martinez, E.; Bastida-Martinez, E.; Perez-Castano, R.; Iniesta, A.A.; Fontes, M.; Padmanabhan, S.; Elias-Arnanz, M. A bacterial light response reveals an orphan desaturase for human plasmalogen synthesis. Science 2019, 366, 128–132. [Google Scholar] [CrossRef]
- Wallner, S.; Schmitz, G. Plasmalogens the neglected regulatory and scavenging lipid species. Chem. Phys. Lipids 2011, 164, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Morita, S.Y.; Takeuchi, A.; Kitagawa, S. Functional analysis of two isoforms of phosphatidylethanolamine N-methyltransferase. Biochem. J. 2010, 432, 387–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.K.; Hajra, A.K. High incorporation of dietary 1-O-heptadecyl glycerol into tissue plasmalogens of young rats. FEBS Lett. 1988, 227, 187–190. [Google Scholar] [CrossRef] [Green Version]
- Blank, M.L.; Cress, E.A.; Smith, Z.L.; Snyder, F. Dietary supplementation with ether-linked lipids and tissue lipid composition. Lipids 1991, 26, 166–169. [Google Scholar] [CrossRef]
- Bergan, J.; Skotland, T.; Sylvanne, T.; Simolin, H.; Ekroos, K.; Sandvig, K. The ether lipid precursor hexadecylglycerol causes major changes in the lipidome of HEp-2 cells. PLoS ONE 2013, 8, e75904. [Google Scholar] [CrossRef] [Green Version]
- Rasmiena, A.A.; Barlow, C.K.; Stefanovic, N.; Huynh, K.; Tan, R.; Sharma, A.; Tull, D.; de Haan, J.B.; Meikle, P.J. Plasmalogen modulation attenuates atherosclerosis in ApoE- and ApoE/GPx1-deficient mice. Atherosclerosis 2015, 243, 598–608. [Google Scholar] [CrossRef]
- Tham, Y.K.; Huynh, K.; Mellett, N.A.; Henstridge, D.C.; Kiriazis, H.; Ooi, J.Y.Y.; Matsumoto, A.; Patterson, N.L.; Sadoshima, J.; Meikle, P.J.; et al. Distinct lipidomic profiles in models of physiological and pathological cardiac remodeling, and potential therapeutic strategies. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, L.; Smith, T.; Senanayake, V.; Mochizuki, A.; Miville-Godbout, E.; Goodenowe, D.; Di Paolo, T. Plasmalogen precursor analog treatment reduces levodopa-induced dyskinesias in parkinsonian monkeys. Behav. Brain Res. 2015, 286, 328–337. [Google Scholar] [CrossRef]
- Eichberg, J.; Gilbertson, J.R.; Karnovsky, M.L. Neutral plasmalogens analogous to the neutral triglycerides. J. Biol. Chem. 1961, 236, PC15–PC16. [Google Scholar] [CrossRef]
- Das, A.K.; Holmes, R.D.; Wilson, G.N.; Hajra, A.K. Dietary ether lipid incorporation into tissue plasmalogens of humans and rodents. Lipids 1992, 27, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Dilbaz, S.; Cossmann, J.; Hoang, A.C.; Diedrich, V.; Herwig, A.; Harauma, A.; Hoshi, Y.; Moriguchi, T.; Landgraf, K.; et al. Breast milk alkylglycerols sustain beige adipocytes through adipose tissue macrophages. J. Clin. Investig. 2019, 129, 2485–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, E.R.; Keller, M.A.; Sailer, S.; Lackner, K.; Koch, J.; Hermann, M.; Coassin, S.; Golderer, G.; Werner-Felmayer, G.; Zoeller, R.A.; et al. The TMEM189 gene encodes plasmanylethanolamine desaturase which introduces the characteristic vinyl ether double bond into plasmalogens. Proc. Natl. Acad. Sci. USA 2020, 117, 7792–7798. [Google Scholar] [CrossRef] [Green Version]
- Huynh, K.; Barlow, C.K.; Jayawardana, K.S.; Weir, J.M.; Mellett, N.A.; Cinel, M.; Magliano, D.J.; Shaw, J.E.; Drew, B.G.; Meikle, P.J. High-Throughput Plasma Lipidomics: Detailed Mapping of the Associations with Cardiometabolic Risk Factors. Cell Chem. Biol. 2019, 26, 71–84.e74. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paul, S.; Rasmiena, A.A.; Huynh, K.; Smith, A.A.T.; Mellett, N.A.; Jandeleit-Dahm, K.; Lancaster, G.I.; Meikle, P.J. Oral Supplementation of an Alkylglycerol Mix Comprising Different Alkyl Chains Effectively Modulates Multiple Endogenous Plasmalogen Species in Mice. Metabolites 2021, 11, 299. https://doi.org/10.3390/metabo11050299
Paul S, Rasmiena AA, Huynh K, Smith AAT, Mellett NA, Jandeleit-Dahm K, Lancaster GI, Meikle PJ. Oral Supplementation of an Alkylglycerol Mix Comprising Different Alkyl Chains Effectively Modulates Multiple Endogenous Plasmalogen Species in Mice. Metabolites. 2021; 11(5):299. https://doi.org/10.3390/metabo11050299
Chicago/Turabian StylePaul, Sudip, Aliki A. Rasmiena, Kevin Huynh, Adam Alexander T. Smith, Natalie A. Mellett, Karin Jandeleit-Dahm, Graeme I. Lancaster, and Peter J. Meikle. 2021. "Oral Supplementation of an Alkylglycerol Mix Comprising Different Alkyl Chains Effectively Modulates Multiple Endogenous Plasmalogen Species in Mice" Metabolites 11, no. 5: 299. https://doi.org/10.3390/metabo11050299
APA StylePaul, S., Rasmiena, A. A., Huynh, K., Smith, A. A. T., Mellett, N. A., Jandeleit-Dahm, K., Lancaster, G. I., & Meikle, P. J. (2021). Oral Supplementation of an Alkylglycerol Mix Comprising Different Alkyl Chains Effectively Modulates Multiple Endogenous Plasmalogen Species in Mice. Metabolites, 11(5), 299. https://doi.org/10.3390/metabo11050299