
The Induction of the Isoflavone Biosynthesis Pathway Is Associated with Resistance to Common Bacterial Blight in Phaseolus vulgaris L.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
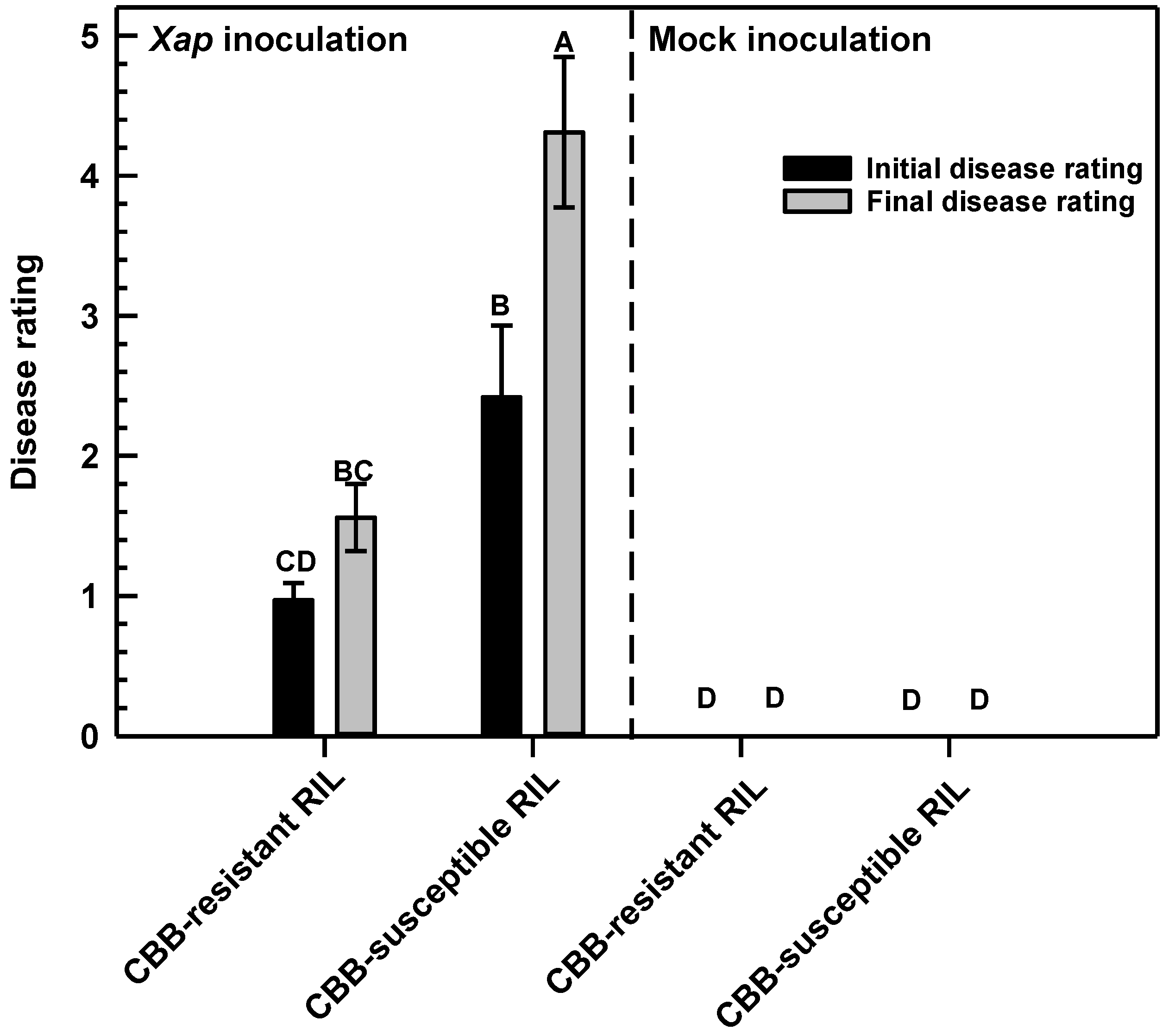
2.1. CBB Symptoms Were Evident in Leaves of the CBB-Susceptible P. vulgaris but Not CBB-Resistant Plants
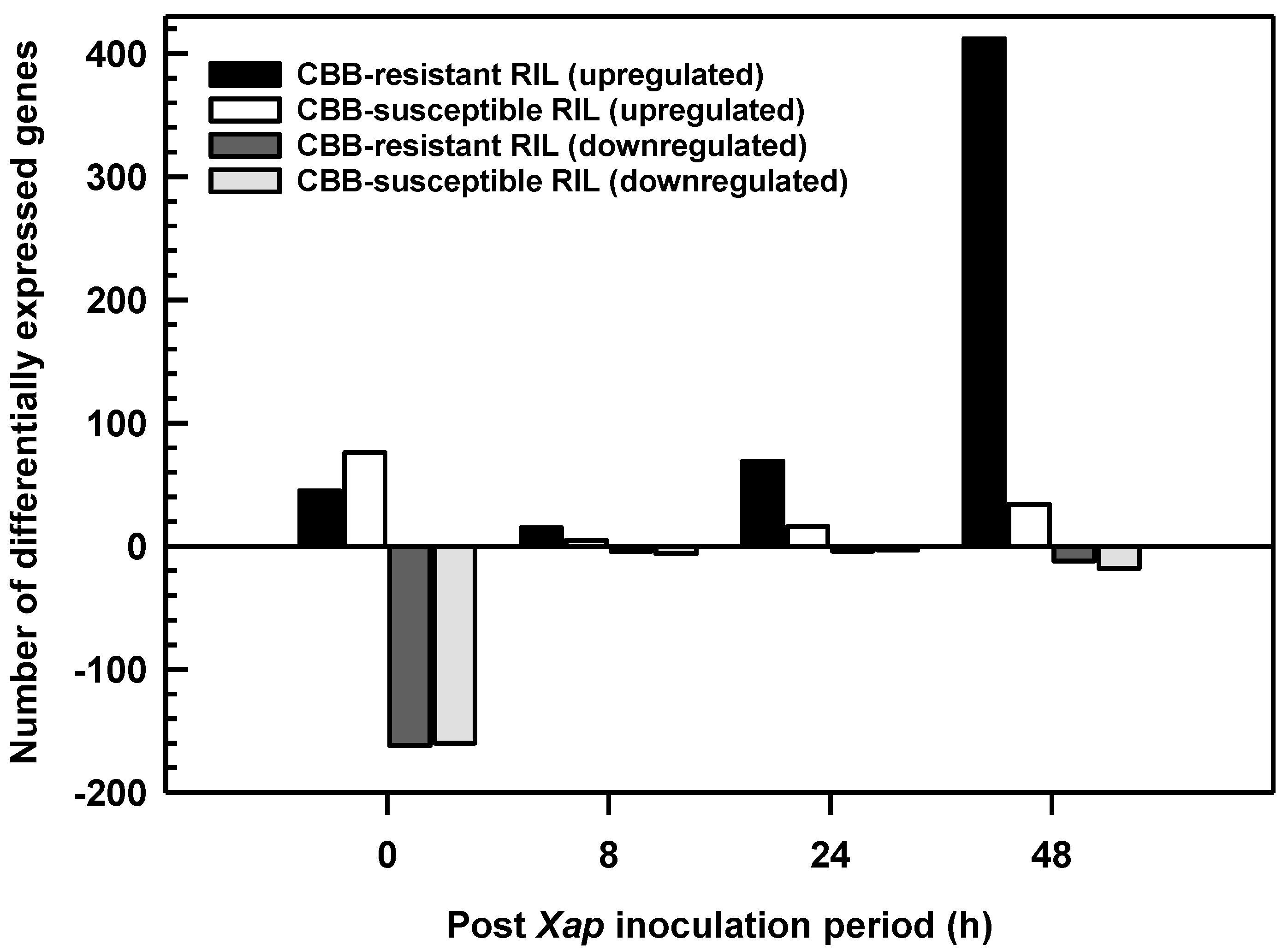
2.2. P. vulgaris RIL Leaf Transcriptomes Changed Following Treatment with X. axonopodis
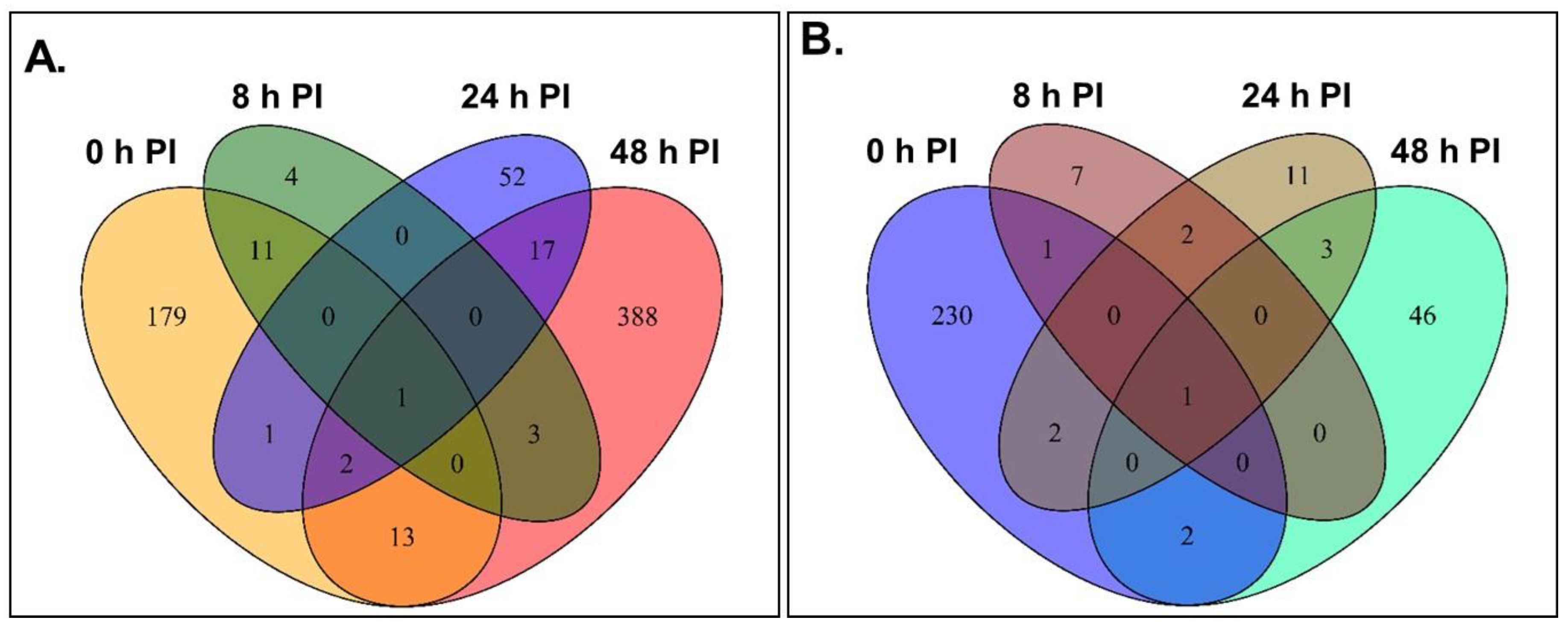
2.3. Differental Gene Expression Was Most Evident in the CBB-Resistant RIL at 48 h Following Inoculation with X. axonopodis
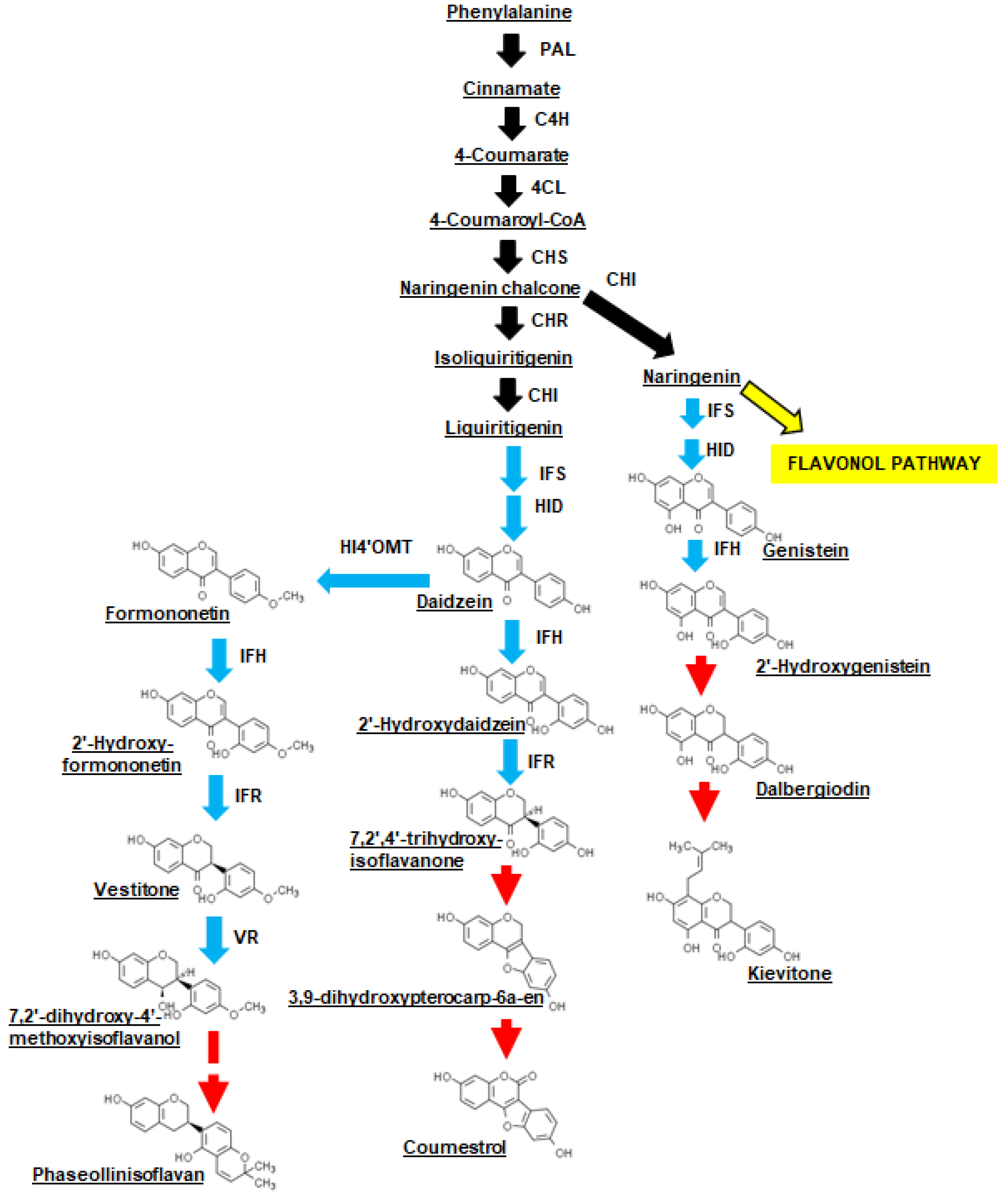
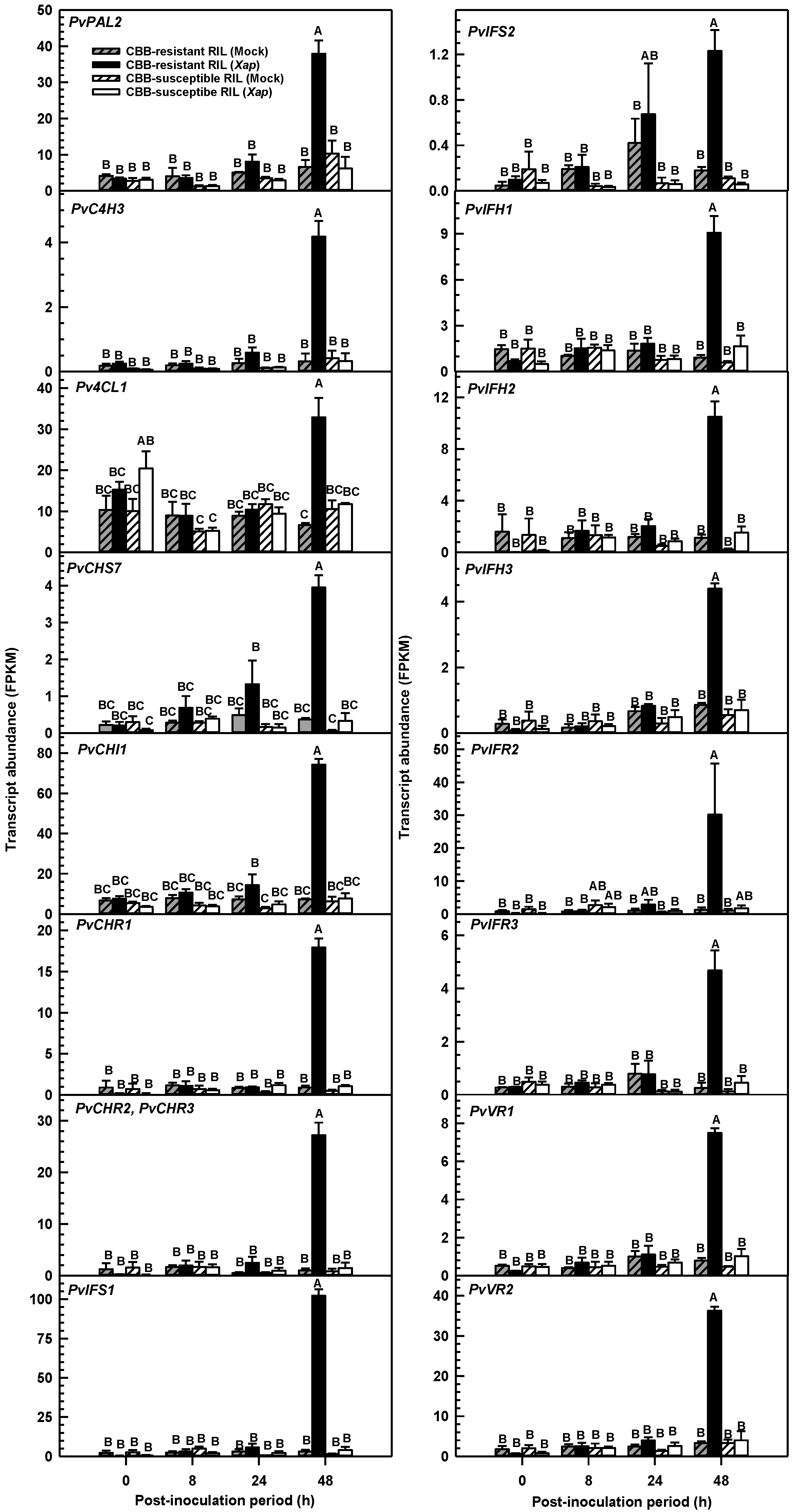
2.4. Isoflavone Biosynthesis Gene Transcripts Were More Abundant in the CBB-Resistant RIL at 48 h Following Inoculation with X. axonopodis
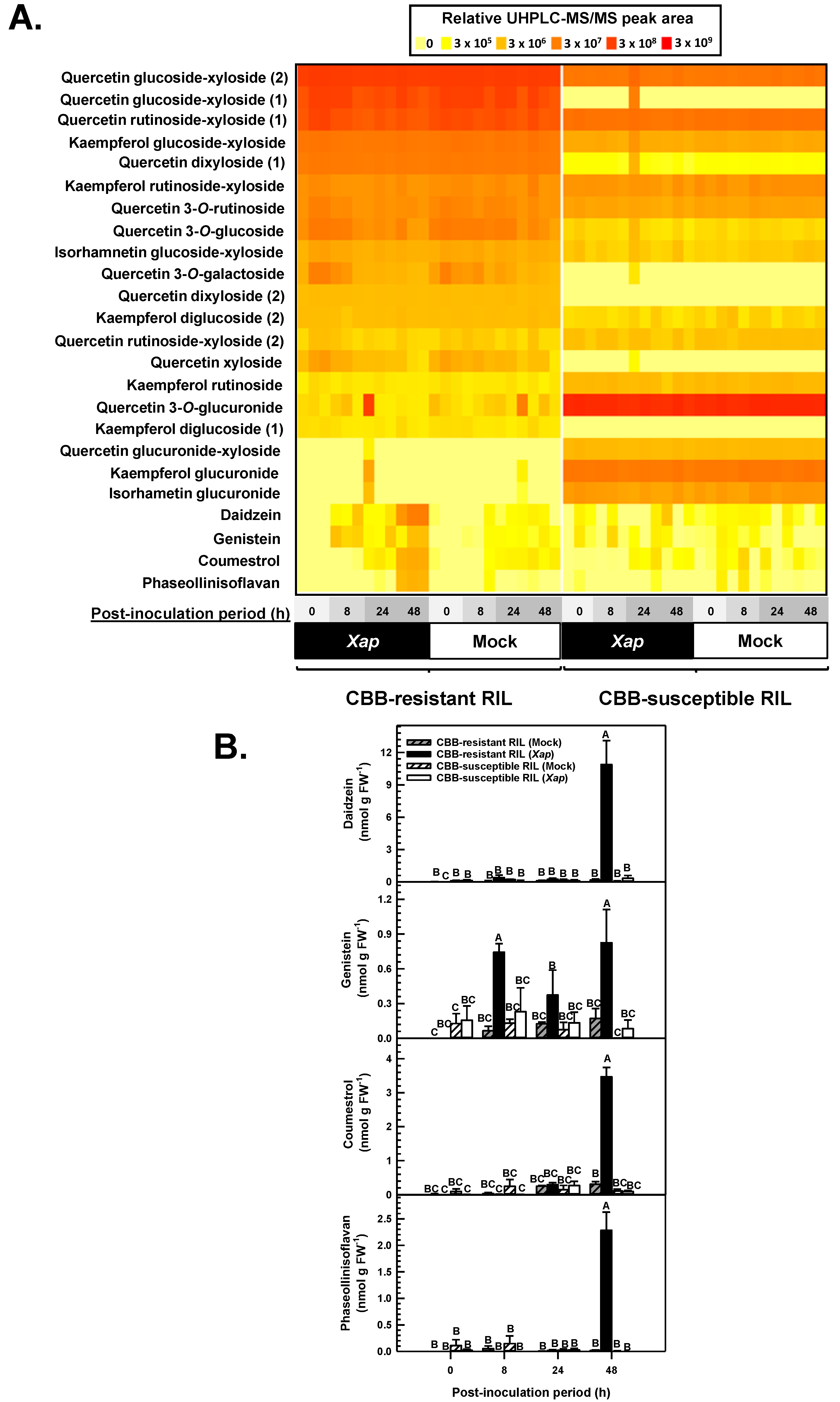
2.5. Isoflavones and Phytoalexins Accumulated in the Leaves of CBB-Resistant Plants Treated with X. axonopodis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Disease Inoculation
4.3. RNA Extraction and Sequencing
4.4. Transcript Estimation and Analysis
4.5. Metabolite Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Database. Food and Agriculture Organization of the United Nations, Statistics Division, Rome. 2013. Available online: http://fenix.fao.org/faostat/beta/en/#home (accessed on 16 February 2021).
- Perry, G.; Pauls, K.P. Common bacterial blight in Phaseolus vulgaris. In Agricultural Research Updates; Hendriks, B.P., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 239–264. [Google Scholar]
- Akhavan, A.; Bahar, M.; Askarian, H.; Lak, M.R.; Nazemi, A.; Zamani, Z. Bean common bacterial blight: Pathogen epiphytic life and effect of irrigation practices. SpringerPlus 2013, 2, 41. [Google Scholar] [CrossRef] [Green Version]
- Boersma, J.G.; Hou, A.; Gillard, C.L.; McRae, K.B.; Conner, R.L. Impact of common bacterial blight on the yield, seed weight and seed discoloration of different market classes of dry beans (Phaseolus vulgaris L.). Can. J. Plant Sci. 2015, 95, 703–710. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Miklas, P.N. Breeding common bean for resistance to common blight: A review. Crop Sci. 2015, 55, 971–984. [Google Scholar] [CrossRef]
- Singh, S.P.; Muñoz, C.G. Resistance to common bacterial blight among Phaseolus species and common bean improvement. Crop Sci. 1999, 39, 80–89. [Google Scholar] [CrossRef]
- Belete, T.; Bastas, K.K. Common bacterial blight (Xanthomonas axonopodis pv. phaseoli) of beans with special focus on Ethiopian condition. J. Plant Pathol. Microbiol. 2017, 8, 403. [Google Scholar]
- Tar’an, B.; Michaels, T.E.; Pauls, K.P. Mapping genetic factors affecting the reaction to Xanthomonas axonopodis pv. phaseoli in Phaseolus vulgaris L. under field conditions. Genome 2001, 44, 1046–1056. [Google Scholar] [CrossRef]
- Shi, C.; Yu, K.; Xie, W.; Perry, G.; Navabi, A.; Pauls, K.P.; Miklas, P.N.; Fourie, D. Development of candidate gene markers associated to common bacterial blight resistance in common bean. Theor. Appl. Genet. 2012, 125, 1525–1537. [Google Scholar] [CrossRef]
- Durham, K.M.; Xie, W.; Yu, K.; Pauls, K.P.; Lee, E.; Navabi, A. Interaction of common bacterial blight quantitative trait loci in a resistant inter-cross population of common bean. Plant Breed. 2013, 132, 658–666. [Google Scholar] [CrossRef]
- Perry, G.; DiNatale, C.; Xie, W.; Navabi, A.; Reinprecht, Y.; Crosby, W.; Yu, K.; Shi, C.; Pauls, K.P. A comparison of the molecular organization of genomic regions associated with resistance to common bacterial blight in two Phaseolus vulgaris genotypes. Front. Plant Sci. 2013, 4, 318. [Google Scholar] [CrossRef] [Green Version]
- Sepiol, C.J.; Yu, J.; Dhaubhadel, S. Genome-wide identification of Chalcone Reductase gene family in soybean: Insight into root-specific GmCHRs and Phytophthora sojae resistance. Front. Plant Sci. 2017, 8, 2073. [Google Scholar] [CrossRef] [Green Version]
- Naoumkina, M.; Farag, M.A.; Sumner, L.W.; Tang, Y.; Liu, C.J.; Dixon, R.A. Different mechanisms for phytoalexin induction by pathogen and wound signals in Medicago truncatula. Proc. Natl. Acad. Sci. USA 2007, 104, 17909–17915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliferis, K.A.; Faubert, D.; Jabaji, S. A metabolic profiling strategy for the dissection of plant defense against fungal pathogens. PLoS ONE 2014, 9, e111930. [Google Scholar] [CrossRef] [Green Version]
- Ishiga, Y.; Uppalapati, S.R.; Gill, U.S.; Huhman, D.; Tang, Y.; Mysore, K.S. Transcriptomic and metabolomic analyses identify a role for chlorophyll catabolism and phytoalexin during Medicago nonhost resistance against Asian soybean rust. Sci. Rep. 2015, 5, 13061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Hassan, S.; Kidd, B.N.; Garg, G.; Mathesius, U.; Singh, K.B.; Anderson, J.P. Ethylene signaling is important for isoflavonoid-mediated resistance to Rhizoctonia solani in roots of Medicago truncatula. Mol. Plant Microbe Interact. 2017, 30, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Gill, U.S.; Uppalapati, S.R.; Gallego-Giraldo, L.; Ishiga, Y.; Dixon, R.A.; Mysore, K.S. Metabolic flux towards the (iso)flavonoid pathway in lignin modified alfalfa lines induces resistance against Fusarium oxysporum f. sp. medicaginis. Plant Cell Environ. 2018, 41, 1997–2007. [Google Scholar]
- Chen, L.; Wu, Q.; He, W.; He, T.; Wu, Q.; Miao, Y. Combined de novo transcriptome and metabolome analysis of common bean response to Fusarium oxysporum f. sp. phaseoli infection. Int. J. Mol. Sci. 2019, 20, 6278. [Google Scholar] [CrossRef] [Green Version]
- Tohge, T.; Perez de Souza, L.; Fernie, A.R. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.L.; Gijzen, M.; Qutob, D.; Dixon, R.A. Molecular characterization of the enzyme catalyzing the aryl migration reaction of isoflavonoid biosynthesis in soybean. Arch. Biochem. Biophys. 1999, 367, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.; Yu, O.; Lau, S.M.C.; O’Keefe, D.P.; Odell, J.; Fader, G.; McGonigle, B. Identification and expression of isoflavone synthase, the key enzyme for biosynthesis of isoflavones in legumes. Nat. Biotechnol. 2000, 18, 208–212. [Google Scholar] [CrossRef]
- Wang, X.; He, X.; Lin, J.; Shao, H.; Chang, Z.; Dixon, R.A. Crystal structure of isoflavone reductase from alfalfa (Medicago sativa L.). J. Mol. Biol. 2006, 358, 1341–1352. [Google Scholar] [CrossRef]
- Uchida, K.; Akashi, T.; Aoki, T. The missing link in leguminous pterocarpan biosynthesis is a dirigent domain-containing protein with isoflavanol dehydratase activity. Plant Cell Physiol. 2017, 58, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Durango, D.; Pulgarin, N.; Echeverri, F.; Escobar, G.; Quiñones, W. Effect of salicylic acid and structurally related compounds in the accumulation of phytoalexins in cotyledons of common bean (Phaseolus vulgaris L.) cultivars. Molecules 2013, 18, 10609–10628. [Google Scholar] [CrossRef] [Green Version]
- Aisyah, S.; Gruppen, H.; Andini, S.; Bettonvil, M.; Severing, E.; Vincken, J.P. Variation in accumulation of isoflavonoids in Phaseoleae seedlings elicited by Rhizopus. Food Chem. 2016, 196, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.; Kang, Y.G.; Lee, T.; Kim, M.; Yoon, M.Y.; Lee, E.; Yang, X.; Kim, D.; Kim, Y.J.; Lee, T.R.; et al. Comprehensive RNA sequencing and co-expression network analysis to complete the biosynthetic pathway of coumestrol, a phytoestrogen. Sci. Rep. 2019, 9, 1934. [Google Scholar] [CrossRef]
- Fischer, D.; Ebenau-Jehle, C.; Grisebach, H. Purification and characterization of pterocarpan synthase from elicitor-challenged soybean cell cultures. Phytochemistry 1990, 29, 2879–2882. [Google Scholar] [CrossRef]
- Guo, L.; Dixon, R.A.; Paiva, N.L. Conversion of vestitone to medicarpin in alfalfa (Medicago sativa L.) is catalyzed by two independent enzymes. Identification, purification, and characterization of vestitone reductase and 7,2’-dihydroxy-4’-methoxyisoflavanol dehydratase. J. Biol. Chem. 1994, 269, 22372–22378. [Google Scholar] [CrossRef]
- Perez de Sousa, L.; Scossa, F.; Proost, S.; Bitocchi, E.; Papa, R.; Toghe, T.; Fernie, A.R. Multi-tissue integration of transcriptomic and specialized metabolite profiling provides tools for assessing the common bean (Phaseolus vulgaris) metabolome. Plant J. 2019, 97, 1132–1153. [Google Scholar] [CrossRef] [Green Version]
- Freixas-Coutin, J.A.; Munholland, S.; Silva, A.; Subedi, S.; Lukens, L.; Crosby, W.L.; Pauls, K.P.; Bozzo, G.G. Proanthocyanidin accumulation and transcriptional responses in the seed coat of cranberry beans (Phaseolus vulgaris L.) with different susceptibility to postharvest darkening. BMC Plant Biol. 2017, 17, 89. [Google Scholar] [CrossRef] [PubMed]
- Ripodas, C.; Dalla Via, V.; Aguilar, O.M.; Zanetti, M.E.; Blanco, F.A. Knock-down of a member of the isoflavone family impairs growth and nodulation in Phaseolus vulgaris. Plant Physiol. Biochem. 2013, 68, 81–89. [Google Scholar] [CrossRef]
- Foucher, J.; Ruh, M.; Préveaux, A.; Carrère, S.; Pelletier, S.; Briand, M.; Serre, R.F.; Jacques, M.A.; Chen, N.W.G. Common bean resistance to Xanthomonas is associated with upregulation of the salicylic acid pathway and downregulation of photosynthesis. BMC Genomics 2020, 21, 566. [Google Scholar]
- Xie, W.; Yu, K.; Pauls, K.P.; Navabi, A. Application of image analysis in studies of quantitative disease resistance, exemplified using common bacterial blight–common bean pathosystem. Phytopathology 2012, 102, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.G.; Park, B.K.; Yoo, C.H.; Jeon, E.; Oh, J.; Hwang, I. Characterization of the Xanthomonas axonopodis pv. glycines hrp pathogenicity island. J. Bacteriol. 2003, 185, 3155–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, M.; Beaver, J.S.; Porch, T.G. Dominant gene for common bean resistance to common bacterial blight caused by Xanthomonas axonopodis pv. phaseoli. Euphytica 2011, 179, 373–382. [Google Scholar] [CrossRef]
- Velasco, P.; Lema, M.; Francisco, M.; Soengas, P.; Cartea, M.E. In vivo and in vitro effects of secondary metabolites against Xanthomonas campestris pv. campestris. Molecules 2013, 18, 11131–11143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padder, B.A.; Kamfwa, K.; Awale, H.E.; Kelly, J.D. Transcriptome profiling of the Phaseolus vulgaris—Colletotrichum lindemuthianum pathosystem. PLoS ONE 2016, 11, e0165823. [Google Scholar] [CrossRef]
- Chen, J.; Pang, W.; Chen, B.; Zhang, C.; Piao, Z. Transcriptome analysis of Brassica rapa near-isogenic lines carrying clubroot-resistant and –susceptible alleles in response to Plasmodiophora brassicae during early infection. Front. Plant Sci. 2016, 6, 1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayo-Prieto, S.; Marra, R.; Vinale, F.; Rodríguez-González, Á.; Woo, S.L.; Lorito, M.; Gutiérrez, S.; Casquero, P.A. Effect of Trichoderma velutinum and Rhizoctonia solani on the metabolome of bean plants (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2019, 20, 549. [Google Scholar] [CrossRef] [Green Version]
- Botero, L.; Vizcaíno, S.; Quiñones, W.; Echeverri, F.; Gil, J.; Durango, D. Increased accumulation of isoflavonoids in common bean (Phaseolus vulgaris L.) tissues treated with 1-oxo-indane-4-carboxylic acid derivatives. Biotech. Rep. 2021, 29, e00601. [Google Scholar] [CrossRef]
- Chen, J.; Ullah, C.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Sclerotinia sclerotiorum circumvents flavonoid defenses by catabolizing flavonol glycosides and aglycones. Plant Physiol. 2019, 180, 1975–1987. [Google Scholar] [CrossRef] [Green Version]
- Dastmalchi, M.; Chapman, P.; Yu, J.; Austin, R.S.; Dhaubhadel, S. Transcriptomic evidence for the control of soybean root isoflavonoid content by regulation of overlapping phenylpropanoid pathways. BMC Genomics 2017, 18, 70. [Google Scholar] [CrossRef] [Green Version]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- Tonnessen, B.W.; Manoslava, P.; Lang, J.M.; Baraoidan, M.; Bordeos, A.; Mauleon, R.; Oard, J.; Hulbert, S.; Leung, H.; Leach, J.E. Rice phenylalanine ammonia-lyase gene OsPAL4 is associated with broad spectrum disease resistance. Plant Mol. Biol. 2015, 87, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Solekha, R.; Susanto, F.A.; Joko, T.; Nuringtyas, T.R.; Purwestri, Y.A. Phenylalanine ammonia lyase (PAL) contributes to the resistance of black rice against Xanthomonas oryzae pv. oryzae. J. Plant Pathol. 2020, 102, 359–365. [Google Scholar] [CrossRef]
- Dao, T.T.H.; Linthorst, H.J.M.; Verproote, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Zabala, G.; Zou, J.; Tuteja, J.; Gonzalez, D.O.; Clough, S.J.; Vodkin, L.O. Transcriptome changes in the phenylpropanoid pathway of Glycine max in response to Pseudomonas syringae infection. BMC Plant Biol. 2006, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Robison, F.M.; Turner, M.F.; Jahn, C.E.; Schwartz, H.F.; Prenni, J.E.; Brick, M.A.; Heuberger, A.L. Common bean varieties demonstrate differential physiological and metabolic responses to the pathogenic fungus Sclerotinia sclerotiorum. Plant Cell Environ. 2018, 41, 2141–2154. [Google Scholar] [PubMed]
- Durango, D.; Quiñones, W.; Torres, F.; Rosero, Y.; Gil, J.; Echeverri, F. Phytoalexin accumulation in Colombian bean varieties and aminosugars as elicitors. Molecules 2002, 7, 817–832. [Google Scholar] [CrossRef] [Green Version]
- He, X.Z.; Dixon, R.A. Genetic manipulation of isoflavone 7-O-methyltransferase enhances biosynthesis of 4’-O-methylated isoflavonoid phytoalexins and disease resistance in alfalfa. Plant Cell 2000, 12, 1689–1702. [Google Scholar]
- Shimada, N.; Akashi, T.; Aoki, T.; Ayabe, S.I. Induction of isoflavonoid pathway in the model legume Lotus japonicus: Molecular characterization of enzymes involved in phytoalexin biosynthesis. Plant Sci. 2000, 160, 37–47. [Google Scholar] [CrossRef]
- Cheng, Q.; Li, N.; Dong, L.; Zhang, D.; Fan, S.; Jiang, L.; Wang, X.; Xu, P.; Zhang, S. Overexpression of soybean isoflavone reductase (GmIFR) enhances resistance to Phytophthora sojae in soybean. Front. Plant Sci. 2015, 6, 1024. [Google Scholar] [CrossRef] [Green Version]
- López-Meyer, M.; Paiva, N.L. Immunolocalization of vestitone reductase and isoflavone reductase, two enzymes involved in the biosynthesis of the phytoalexin medicarpin. Physiol. Mol. Plant Pathol. 2002, 61, 15–30. [Google Scholar] [CrossRef]
- Uchida, K.; Aoki, T.; Suzuki, H.; Akashi, T. Molecular cloning and biochemical characterization of isoflav-3-ene synthase, a key enzyme for the biosynthesis of (+)-pisatin and coumestrol. Plant Biotechnol. 2020, 37, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Thamil Arasan, S.K.; Park, J.I.; Ahmed, N.U.; Jung, H.J.; Hur, Y.; Kang, K.K.; Lim, Y.P.; Nou, I.S. Characterization and expression analysis of dirigent family genes related to stresses in Brassica. Plant Physiol. Biochem. 2013, 67, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Seneviratne, H.K.; Dalisay, D.S.; Kim, K.W.; Moinuddin, S.G.A.; Yang, H.; Hartshorn, C.M.; Davin, L.B.; Lewis, N.G. Non-host disease resistance response in pea (Pisum sativum) pods: Biochemical function of DRR206 and phytoalexin pathway localization. Phytochemistry 2015, 113, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhao, M.; Liu, T.; Dong, L.; Cheng, Q.; Wu, J.; Wang, L.; Chen, X.; Zhang, C.; Lu, W.; et al. A novel soybean dirigent gene gmdir22 contributes to promotion of lignan biosynthesis and enhances resistance to Phytophthora sojae. Front. Plant Sci. 2017, 8, 1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.Y.; Choi, G.J.; Son, S.W.; Jang, K.S.; Lim, H.K.; Lee, S.O.; Sung, N.D.; Cho, K.Y.; Kim, J.C. Isolation and antifungal activity of lignans from Myristica fragrans against various plant pathogenic fungi. Pest Manag. Sci. 2007, 63, 935–940. [Google Scholar] [CrossRef]
- Jahan, M.A.; Harris, B.; Lowery, M.; Infante, A.M.; Percifield, R.J.; Kovinich, N. Glyceollin transcription factor GmMYB29A2 regulates soybean resistance to Phytophthora sojae. Plant Physiol. 2020, 183, 530–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahan, M.A.; Harris, B.; Lowery, M.; Coburn, K.; Infante, A.M.; Percifield, R.J.; Ammer, A.G.; Kovinich, N. The NAC family transcription factor GmNAC42-1 regulates biosynthesis of the anticancer and neuroprotective glyceollins in soybean. BMC Genomics 2019, 20, 149. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Dhanvantari, B.N. Transfer of common blight (Xanthomonas campestris pv phaseoli) resistance from Phaseolus coccineus Lam. to P. vulgaris L. through interspecific hybridization. Can. J. Plant. Sci. 1987, 67, 685–695. [Google Scholar] [CrossRef]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli physiology in Luria-Bertani broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [Green Version]
- Andrus, C.F. A method of testing beans for resistance to bacterial blight. Phytopathology 1948, 38, 757–759. [Google Scholar]
- Yu, K.; Park, S.J.; Poysa, V. Marker-assisted selection of common beans for resistance to common bacterial blight: Efficacy and economics. Plant. Breed. 2000, 119, 411–415. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Goff, L.; Trapnell, C.; Kelley, D. Cummerbund: Analysis, Exploration, Manipulation, and Visualization of Cufflinks High-Throughput Sequencing Data. R Package Version 2.28.0. Available online: https://www.bioconductor.org/packages/release/bioc/html/cummeRbund.html (accessed on 12 July 2019).
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [Green Version]
- Roepke, J.; Bozzo, G.G. Arabidopsis thaliana β-glucosidase BGLU15 attacks flavonol 3-O-β-glucoside-7-O-α-rhamnosides. Phytochemistry 2015, 109, 14–24. [Google Scholar] [CrossRef]
- Available online: https://www.mzcloud.org/ (accessed on 22 April 2021).
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cox, L.D.; Munholland, S.; Mats, L.; Zhu, H.; Crosby, W.L.; Lukens, L.; Pauls, K.P.; Bozzo, G.G. The Induction of the Isoflavone Biosynthesis Pathway Is Associated with Resistance to Common Bacterial Blight in Phaseolus vulgaris L. Metabolites 2021, 11, 433. https://doi.org/10.3390/metabo11070433
Cox LD, Munholland S, Mats L, Zhu H, Crosby WL, Lukens L, Pauls KP, Bozzo GG. The Induction of the Isoflavone Biosynthesis Pathway Is Associated with Resistance to Common Bacterial Blight in Phaseolus vulgaris L. Metabolites. 2021; 11(7):433. https://doi.org/10.3390/metabo11070433
Chicago/Turabian StyleCox, Laura D., Seth Munholland, Lili Mats, Honghui Zhu, William L. Crosby, Lewis Lukens, Karl Peter Pauls, and Gale G. Bozzo. 2021. "The Induction of the Isoflavone Biosynthesis Pathway Is Associated with Resistance to Common Bacterial Blight in Phaseolus vulgaris L." Metabolites 11, no. 7: 433. https://doi.org/10.3390/metabo11070433
APA StyleCox, L. D., Munholland, S., Mats, L., Zhu, H., Crosby, W. L., Lukens, L., Pauls, K. P., & Bozzo, G. G. (2021). The Induction of the Isoflavone Biosynthesis Pathway Is Associated with Resistance to Common Bacterial Blight in Phaseolus vulgaris L. Metabolites, 11(7), 433. https://doi.org/10.3390/metabo11070433