
A Metabolomic Study of the Analgesic Effect of Lappaconitine Hydrobromide (LAH) on Inflammatory Pain
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Paw Withdrawal Latency
2.3. Hematoxylin and Eosin (H&E) Staining
- (1)
- Dewaxing: Xylene (10023418, Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) I for 20 min; Xylene II for 20 min; 100% ethanol (100092683, Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) I for 5 min; 100% ethanol II for 5 min; 75% ethanol for 5 min; rinsing with tap water.
- (2)
- Staining sections with hematoxylin solution for 3–5 min and rinsing with tap water. Then, we treat the section with hematoxylin differentiation solution and then rinse with tap water. We then treat the section with hematoxylin Scott’s Tap Water bluing and rinse with tap water (HE dye solution set, G1003, Wuhan Servicebio Technology Co., Ltd., Wuhan, Hubei, China).
- (3)
- Treating with 85% ethanol for 5 min; 95% ethanol for 5 min. Finally, we stained the sections with eosin dye for 5 min.
- (4)
- Dehydrating, as follows: 100% ethanol I for 5 min; 100% ethanol II for 5 min; 100% ethanol III for 5 min; Xylene I for 5 min; Xylene II for 5 min. Finally, we seal it with neutral gum (10004160, Sinopharm Chemical Reagent Co., Ltd., Shanghai, China).
- (5)
- Observing with a microscope (NIKON ECLIPSE E100, Nikon-Essilor Co., Ltd., Tokyo, Japan) for the purposes of inspection, image acquisition (NIKON DS-U3, Nikon-Essilor Co., Ltd., Tokyo, Japan), and analysis. The nucleus is blue, while the cytoplasm is red.
2.4. Preparation of Sample for LC-MS/MS
2.5. LC-MS/MS Analysis
2.6. Data Analysis
3. Results
3.1. Paw Withdrawal Latency
3.2. H&E Staining
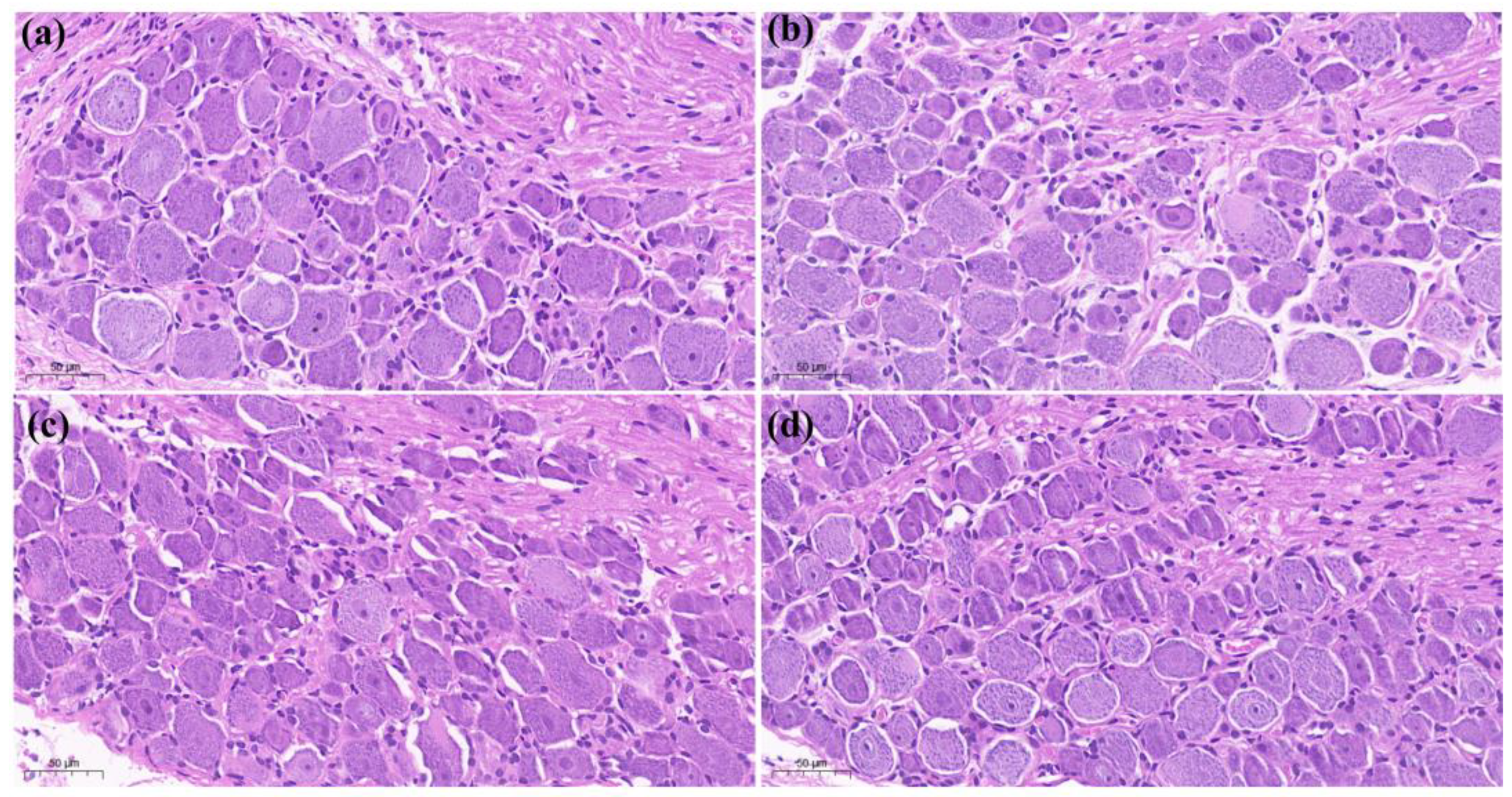
 ”), with a thin spiny layer and visible enlargement. In the treatment group CFA + 4 mg/kg LAH (c) and treatment group CFA + 8 mg/kg LAH (d), the small vesicles decreased, the spiny layer thickened, and the swelling reaction was alleviated. The results showed that the inflammatory swelling of the sole of the foot in rats could be significantly reduced by LAH (Figure 3).
”), with a thin spiny layer and visible enlargement. In the treatment group CFA + 4 mg/kg LAH (c) and treatment group CFA + 8 mg/kg LAH (d), the small vesicles decreased, the spiny layer thickened, and the swelling reaction was alleviated. The results showed that the inflammatory swelling of the sole of the foot in rats could be significantly reduced by LAH (Figure 3).3.3. Differential Analysis of Metabolites
3.3.1. Orthogonal Partial Least-Squares Discriminant Analysis
3.3.2. Univariate Analysis—Fold Change Analysis
3.4. Differential Metabolic Pathway Enrichment Analysis
4. Discussion
5. Conclusions
- (1)
- Lappaconitine has a preferential analgesic effect on CFA-induced inflammatory pain behavior.
- (2)
- CFA-induced inflammatory pain mainly triggered changes in the pathways of retinol metabolism, glycerophospholipid metabolism, and alanine, aspartate, and glutamate metabolism in the DRG of rats; the alpha-linolenic acid metabolism and retinol metabolism were significantly altered after LAH treatment.
- (3)
- These results suggest that LAH could effectively reduce inflammatory pain and might achieve this through regulating the lipid metabolism in rat DRG.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Raja, S.N.; Carr, D.B.; Cohen, M.L.; Finnerup, N.B.; Flor, H.; Gibson, S.B.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Yongjun, Z.; Tingjie, Z.; Xiaoqiu, Y.; Zhiying, F.; Feng, Q.; Guangke, X.; Jinfeng, L.; Fachuan, N.; Xiaohong, J.; Yanqing, L. A survey of chronic pain in China. Libyan J. Med. 2020, 15, 1730550. [Google Scholar] [CrossRef] [PubMed]
- Yong, R.J.; Mullins, P.M.; Bhattacharyya, N. Prevalence of chronic pain among adults in the United States. Pain 2022, 163, e328–e332. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Liu, C.; Gong, G.-H.; Quan, Z.-S. Synthesis, in vitro and in vivo biological evaluation of novel lappaconitine derivatives as potential anti-inflammatory agents. Acta Pharm. Sinica B 2020, 10, 628–645. [Google Scholar] [CrossRef]
- Lu, S.; Wang, S.J. Clinical control study of lappaconitine vs morphine for postoperative epidural analgesia. Chin. J. New Drugs Clin. Remedies 2004, 23, 497–499. [Google Scholar]
- Ameri, A. The effects of Aconitum alkaloids on the central nervous system. Prog Neurobiol 1998, 56, 211–235. [Google Scholar] [CrossRef]
- Teng, G.; Zhang, X.; Zhang, C.; Chen, L.; Sun, W.; Qiu, T.; Zhang, J. Lappaconitine trifluoroacetate contained polyvinyl alcohol nanofibrous membranes: Characterization, biological activities and transdermal application. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 108, 110515. [Google Scholar] [CrossRef]
- Kaixiang, W. Clinical study of Lappaconitine for postoperative patient-controlled subcutaneous analgesia on plasma β-endorphin. China Med. Herald 2010, 7, 17–19. [Google Scholar]
- Ono, M.; Satoh, T. Pharmacological studies of lappaconitine. Analgesic activities. Arzneimittel-Forschung 1988, 38, 892–895. [Google Scholar]
- Wright, S.N. Irreversible block of human heart (hH1) sodium channels by the plant alkaloid lappaconitine. Mol. Pharmacol. 2001, 59, 183–192. [Google Scholar] [CrossRef]
- Ren, M.; Yu, Q.; Shi, C.; Luo, J.-B. Anticancer Activities of C18-, C19-, C20-, and Bis-Diterpenoid Alkaloids Derived from Genus Aconitum. Mol. J. Synth. Chem. Nat. Prod. Chem. 2017, 22, 267. [Google Scholar] [CrossRef]
- Ou, S.; Zhao, Y.-D.; Xiao, Z.; Wen, H.-Z.; Cui, J.; Ruan, H.-Z. Effect of lappaconitine on neuropathic pain mediated by P2X3 receptor in rat dorsal root ganglion. Neurochem. Int. 2011, 58, 564–573. [Google Scholar] [CrossRef]
- Guo, X.; Tang, X.C. Effects of reserpine and 5-HT on analgesia induced by lappaconitine and N-deacetyllappaconitine. Zhongguo Yao Li Xue Bao Acta Pharmacol. Sin. 1990, 11, 14–18. [Google Scholar]
- Ono, M.; Satoh, T. Pharmacological studies on lappaconitine: Possible interaction with endogenous noradrenergic and serotonergic pathways to induce antinociception. Jpn. J. Pharmacol. 1992, 58, 251–257. [Google Scholar] [CrossRef]
- Sun, M.-L.; Ao, J.-P.; Wang, Y.-R.; Huang, Q.; Li, T.-F.; Li, X.-Y.; Wang, Y. Lappaconitine, a C18-diterpenoid alkaloid, exhibits antihypersensitivity in chronic pain through stimulation of spinal dynorphin A expression. Psychopharmacology 2018, 235, 2559–2571. [Google Scholar] [CrossRef] [PubMed]
- Berta, T.; Qadri, Y.; Tan, P.H.; Ji, R.R. Targeting dorsal root ganglia and primary sensory neurons for the treatment of chronic pain. Expert Opin. Ther. Targets 2017, 21, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Wang, S.; Shao, F.; Fang, J.; Xu, Y.-L.; Wang, W.; Sun, H.; Liu, X.; Du, J.; Fang, J. Electroacupuncture Stimulation Alleviates CFA-Induced Inflammatory Pain Via Suppressing P2X3 Expression. Int. J. Mol. Sci. 2019, 20, 3248. [Google Scholar] [CrossRef]
- Pan, H.-L.; Wu, Z.-Z.; Zhou, H.-Y.; Chen, S.; Zhang, H.-M.; Li, D.-P. Modulation of pain transmission by G-protein-coupled receptors. Pharmacol. Ther. 2008, 117, 141–161. [Google Scholar] [CrossRef]
- Liu, J.A.; Yu, J.; Cheung, C.W. Immune Actions on the Peripheral Nervous System in Pain. Int J Mol Sci 2021, 22, 1448. [Google Scholar] [CrossRef]
- Baral, P.; Udit, S.; Chiu, I.M. Pain and immunity: Implications for host defence. Nat. Rev. Immunol. 2019, 19, 433–447. [Google Scholar]
- Lu, Y.; Cao, D.-L.; Ma, L.-J.; Gao, Y.-J. TRAF6 Contributes to CFA-Induced Spinal Microglial Activation and Chronic Inflammatory Pain in Mice. Cell. Mol. Neurobiol. 2021, 42, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.-C.; Liu, T.; Gao, Y.-J. Chemokines in chronic pain: Cellular and molecular mechanisms and therapeutic potential. Pharmacol. Ther. 2020, 212, 107581. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Kim, M.; Hwang, S.W. Molecular mechanisms underlying the actions of arachidonic acid-derived prostaglandins on peripheral nociception. J. Neuroinflamm. 2020, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Patti, G.J.; Yanes, Ó.; Shriver, L.P.; Courade, J.-P.; Tautenhahn, R.; Manchester, M.; Siuzdak, G. Metabolomics implicates altered sphingolipids in chronic pain of neuropathic origin. Nat. Chem. Biol. 2012, 8, 232–234. [Google Scholar] [CrossRef]
- Goodacre, R.; Vaidyanathan, S.; Dunn, W.B.; Harrigan, G.G.; Kell, D.B. Metabolomics by numbers: Acquiring and understanding global metabolite data. Trends Biotechnol. 2004, 22, 245–252. [Google Scholar] [CrossRef]
- Teckchandani, S.; Nagana Gowda, G.A.; Raftery, D.; Curatolo, M. Metabolomics in chronic pain research. Eur. J. Pain 2020, 25, 313–326. [Google Scholar] [CrossRef]
- Adams, S.B.; Setton, L.A.; Kensicki, E.; Bolognesi, M.P.; Toth, A.P.; Nettles, D.L. Global metabolic profiling of human osteoarthritic synovium. Osteoarthr. Cartil. 2012, 20, 64–67. [Google Scholar] [CrossRef]
- Xiao, Z.-Y.; Zhang, Z.; Huang, S.; Lon, J.R.; Xie, S. Metabolic Profiling of Serum for Osteoarthritis Biomarkers. Dis. Markers 2022, 2022, 1800812. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Yan, G.-L.; Wang, P.; Wang, X.-J. Metabolomics for Biomarker Discovery: Moving to the Clinic. BioMed Res. Int. 2015, 2015, 354671. [Google Scholar] [CrossRef]
- Pretorius, C.J.; Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Metabolomics for Biomarker Discovery: Key Signatory Metabolic Profiles for the Identification and Discrimination of Oat Cultivars. Metabolites 2021, 11, 165. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Osthues, T.; Sisignano, M. Oxidized Lipids in Persistent Pain States. Front. Pharmacol. 2019, 10, 1147. [Google Scholar] [CrossRef] [PubMed]
- Bochkov, V.N.; Oskolkova, O.; Birukov, K.G.; Levonen, A.-L.; Binder, C.J.; Stöckl, J. Generation and biological activities of oxidized phospholipids. Antioxid. Redox Signal. 2010, 12, 1009–1059. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, A. Prostaglandin E2 and pain—An update. Biol. Pharm. Bull. 2011, 34, 1170–1173. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, A.M.; Akopian, A.N.; Ruparel, N.B.; Diogenes, A.R.; Weintraub, S.T.; Uhlson, C.L.; Murphy, R.C.; Hargreaves, K.M. Heat generates oxidized linoleic acid metabolites that activate TRPV1 and produce pain in rodents. J. Clin. Investig. 2010, 120, 1617–1626. [Google Scholar] [CrossRef]
- Alsalem, M.; Wong, A.; Millns, P.; Arya, P.H.; Chan, M.S.; Bennett, A.; Barrett, D.A.; Chapman, V.; Kendall, D.A. The contribution of the endogenous TRPV1 ligands 9-HODE and 13-HODE to nociceptive processing and their role in peripheral inflammatory pain mechanisms. Br. J. Pharmacol. 2013, 168, 1961–1974. [Google Scholar] [CrossRef]
- Hiesinger, K.; Wagner, K.M.; Hammock, B.D.; Proschak, E.; Hwang, S.H. Development of multitarget agents possessing soluble epoxide hydrolase inhibitory activity. Prostaglandins Other Lipid Mediat 2019, 140, 31–39. [Google Scholar] [CrossRef]
- Inceoglu, B.; Jinks, S.L.; Schmelzer, K.R.; Waite, T.; Kim, I.H.; Hammock, B.D. Inhibition of soluble epoxide hydrolase reduces LPS-induced thermal hyperalgesia and mechanical allodynia in a rat model of inflammatory pain. Life Sci. 2006, 79, 2311–2319. [Google Scholar] [CrossRef]
- Nakamoto, K.; Tokuyama, S. The role of brain fatty acids in pain. Pain Res. 2018, 33, 193–202. [Google Scholar] [CrossRef]
- Goldberg, R.J.; Katz, J.D. A meta-analysis of the analgesic effects of omega-3 polyunsaturated fatty acid supplementation for inflammatory joint pain. Pain 2007, 129, 210–223. [Google Scholar] [CrossRef]
- Matta, J.A.; Miyares, R.L.; Ahern, G.P. TRPV1 is a novel target for omega-3 polyunsaturated fatty acids. J. Physiol. 2007, 578, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Gronert, K.; Devchand, P.R.; Moussignac, R.-L.; Serhan, C.N. Novel Docosatrienes and 17S-Resolvins Generated from Docosahexaenoic Acid in Murine Brain, Human Blood, and Glial Cells. J. Biol. Chem. 2003, 278, 14677–14687. [Google Scholar] [CrossRef] [PubMed]
- Bazan, N.G. Neuroprotectin D1 (NPD1): A DHA-derived mediator that protects brain and retina against cell injury-induced oxidative stress. Brain Pathol. 2005, 15, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Lü, N.; Xu, Z.Z.; Liu, T.; Serhan, C.N.; Ji, R.R. Resolving TRPV1- and TNF-α-mediated spinal cord synaptic plasticity and inflammatory pain with neuroprotectin D1. J. Neurosci. 2011, 31, 15072–15085. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [Green Version]
- Nagy, K.; Tiuca, I. Importance of Fatty Acids in Physiopathology of Human Body. In Fatty Acids; IntechOpen: London, UK, 2017. [Google Scholar]
- Ma, J.-J.; Han, B.-C.; Yang, Y.; Peng, J.-P. Retinoic acid synthesis and metabolism are concurrent in the mouse uterus during peri-implantation. Cell Tissue Res. 2012, 350, 525–537. [Google Scholar] [CrossRef]
- Lengqvist, J.; Mata De Urquiza, A.; Bergman, A.C.; Willson, T.M.; Sjövall, J.; Perlmann, T.; Griffiths, W.J. Polyunsaturated fatty acids including docosahexaenoic and arachidonic acid bind to the retinoid X receptor alpha ligand-binding domain. Mol. Cell Proteom. 2004, 3, 692–703. [Google Scholar] [CrossRef]
- Yang, F.-C.; Xu, F.; Wang, T.; Chen, G. Roles of vitamin A in the regulation of fatty acid synthesis. World J. Clin. Cases 2021, 9, 4506–4519. [Google Scholar] [CrossRef]
- Keller, H.; Dreyer, C.; Medin, J.A.; Mahfoudi, A.; Ozato, K.; Wahli, W. Fatty acids and retinoids control lipid metabolism through activation of peroxisome proliferator-activated receptor-retinoid X receptor heterodimers. Proc. Natl. Acad. Sci. USA 1993, 90, 2160–2164. [Google Scholar] [CrossRef]
- Yin, S.; Luo, J.; Qian, A.-H.; Du, J.; Yang, Q.; Zhou, S.; Yu, W.; Du, G.; Clark, R.B.; Walters, E.T.; et al. Retinoids activate the irritant receptor TRPV1 and produce sensory hypersensitivity. J. Clin. Investig. 2013, 123, 3941–3951. [Google Scholar] [CrossRef]
- Alique, M.; Lucio, F.J.; Herrero, J.F. Vitamin A active metabolite, all-trans retinoic acid, induces spinal cord sensitization. II. Effects after intrathecal administration. Br. J. Pharmacol. 2006, 149, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Luo, J.; Qian, A.; Yu, W.; Hu, H. LE135, a retinoid acid receptor antagonist, produces pain through direct activation of TRP channels. Br. J. Pharmacol. 2014, 171, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Núñez, V.; Alameda, D.; Rico, D.; Mota, R.A.; Gonzalo, P.; Cedenilla, M.; Fischer, T.; Boscá, L.; Glass, C.K.; Arroyo, A.G.; et al. Retinoid X receptor α controls innate inflammatory responses through the up-regulation of chemokine expression. Proc. Natl. Acad. Sci. USA 2010, 107, 10626–10631. [Google Scholar] [CrossRef] [PubMed]
- Kolseth, I.B.M.; Ågren, J.; Sundvold-gjerstad, V.; Lyngstadaas, S.P.; Wang, J.E.; Dahle, M.K. 9-cis Retinoic Acid Inhibits Inflammatory Responses of Adherent Monocytes and Increases Their Ability to Induce Classical Monocyte Migration. J. Innate Immun. 2011, 4, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Amann, P.M.; Eichmüller, S.B.; Schmidt, J.; Bazhin, A.V. Regulation of gene expression by retinoids. Curr. Med. Chem. 2011, 18, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.K.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Freitag, C.M.; Miller, R.J. Peroxisome proliferator-activated receptor agonists modulate neuropathic pain: A link to chemokines? Front. Cell. Neurosci. 2014, 8, 238. [Google Scholar] [CrossRef]
- Jiang, C.; Ting, A.T.; Seed, B. PPAR-γ agonists inhibit production of monocyte inflammatory cytokines. Nature 1998, 391, 82–86. [Google Scholar] [CrossRef]

 ”.
”.
”.
”.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Saline | CFA + Saline | CFA + 4 mg/kg LAH | CFA + 8 mg/kg LAH |
|---|---|---|---|---|
| Baseline | 37.31 ± 0.59 | 37.22 ± 0.59 | 37.33 ± 0.68 | 36.59 ± 0.55 |
| Day 1 | 38.51 ± 0.48 | 10.52 ± 0.32 * | 13.90 ± 0.31 *# | 18.53 ± 0.32 *#& |
| Day 2 | 37.21 ± 0.37 | 13.32 ± 0.51 * | 19.35 ± 0.49 *# | 25.21 ± 0.64 *#& |
| Day 3 | 37.87 ± 0.57 | 14.59 ± 0.47 * | 22.76 ± 0.43 *# | 30.93 ± 0.54 *#& |
| Day 4 | 37.46 ± 0.72 | 18.56 ± 0.62 * | 24.83 ± 0.51 *# | 31.04 ± 0.59 *#& |
| Day 5 | 38.04 ± 0.63 | 23.86 ± 0.58 * | 28.44 ± 0.35 *# | 32.11 ± 0.43 *#& |
| Day 6 | 37.69 ± 0.61 | 25.34 ± 0.69 * | 28.31 ± 0.49 *# | 35.06 ± 0.56 *#& |
| Day 7 | 37.37 ± 0.63 | 24.88 ± 0.77 * | 30.28 ± 0.63 *# | 35.55 ± 0.30 #& |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, X.; Li, Z.; Mao, Y.; Liu, Z.; Liu, X.; Zhu, X.; Zhang, J. A Metabolomic Study of the Analgesic Effect of Lappaconitine Hydrobromide (LAH) on Inflammatory Pain. Metabolites 2022, 12, 923. https://doi.org/10.3390/metabo12100923
Li X, Wang X, Li Z, Mao Y, Liu Z, Liu X, Zhu X, Zhang J. A Metabolomic Study of the Analgesic Effect of Lappaconitine Hydrobromide (LAH) on Inflammatory Pain. Metabolites. 2022; 12(10):923. https://doi.org/10.3390/metabo12100923
Chicago/Turabian StyleLi, Xu, Xueqi Wang, Zhengdou Li, Ying Mao, Zhao Liu, Xiaoxiao Liu, Xinliang Zhu, and Ji Zhang. 2022. "A Metabolomic Study of the Analgesic Effect of Lappaconitine Hydrobromide (LAH) on Inflammatory Pain" Metabolites 12, no. 10: 923. https://doi.org/10.3390/metabo12100923
APA StyleLi, X., Wang, X., Li, Z., Mao, Y., Liu, Z., Liu, X., Zhu, X., & Zhang, J. (2022). A Metabolomic Study of the Analgesic Effect of Lappaconitine Hydrobromide (LAH) on Inflammatory Pain. Metabolites, 12(10), 923. https://doi.org/10.3390/metabo12100923