
Morinda officinalis Polysaccharides Ameliorates Bone Growth by Attenuating Oxidative Stress and Regulating the Gut Microbiota in Thiram-Induced Tibial Dyschondroplasia Chickens
 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics
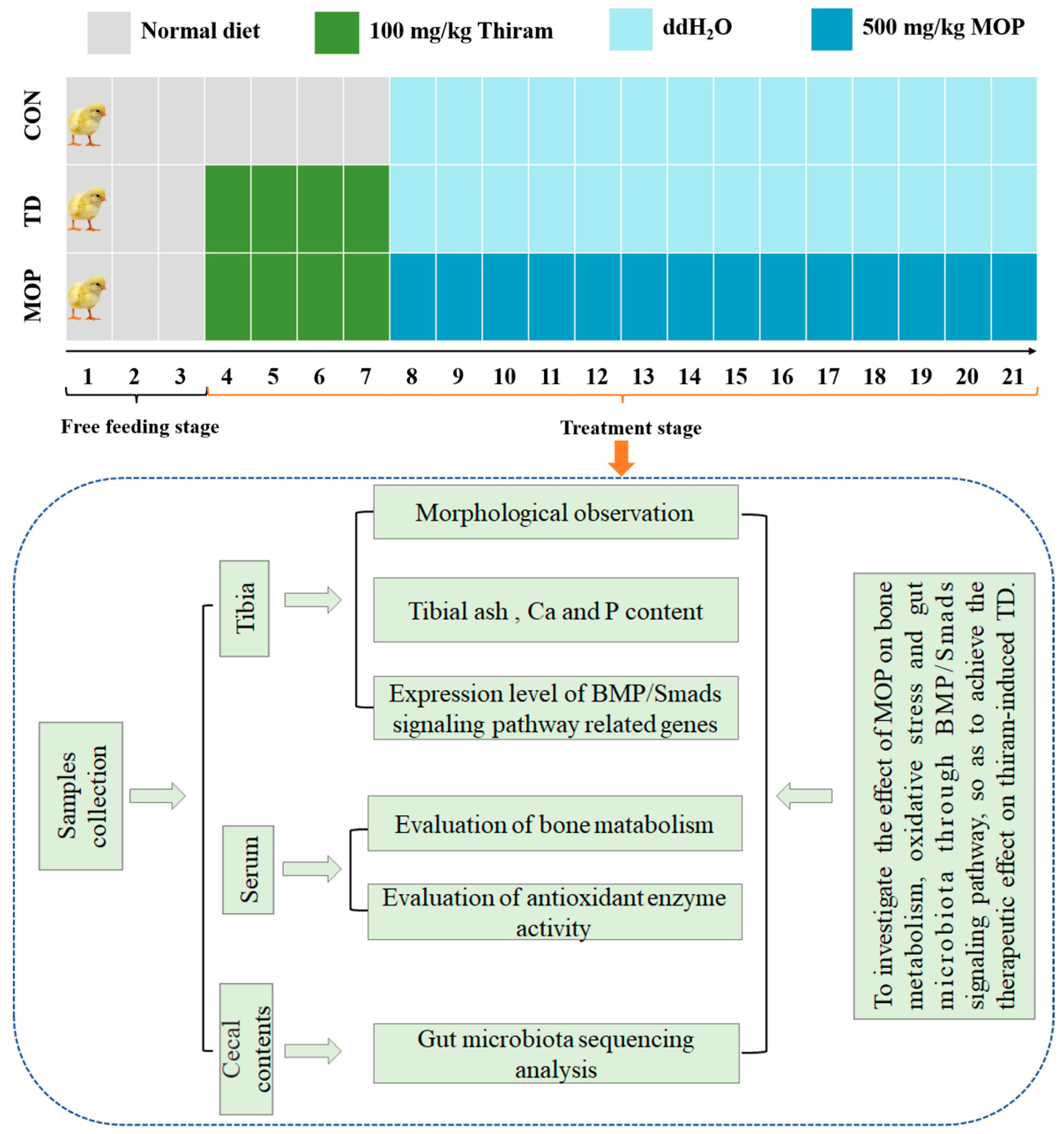
2.2. Chemicals, Animals, and Experimental Protocol
2.3. Determination of Tibia Indicators
2.4. Determination of Tibia Ash and Calcium and Phosphorus Content
2.5. Determination of Plasma Biochemical Indicators
2.6. Histopathological Analysis
2.7. RNA Isolation and RT-qPCR
2.8. Gut Microbiota Sequencing Analysis
2.9. Statistical Analysis
3. Results
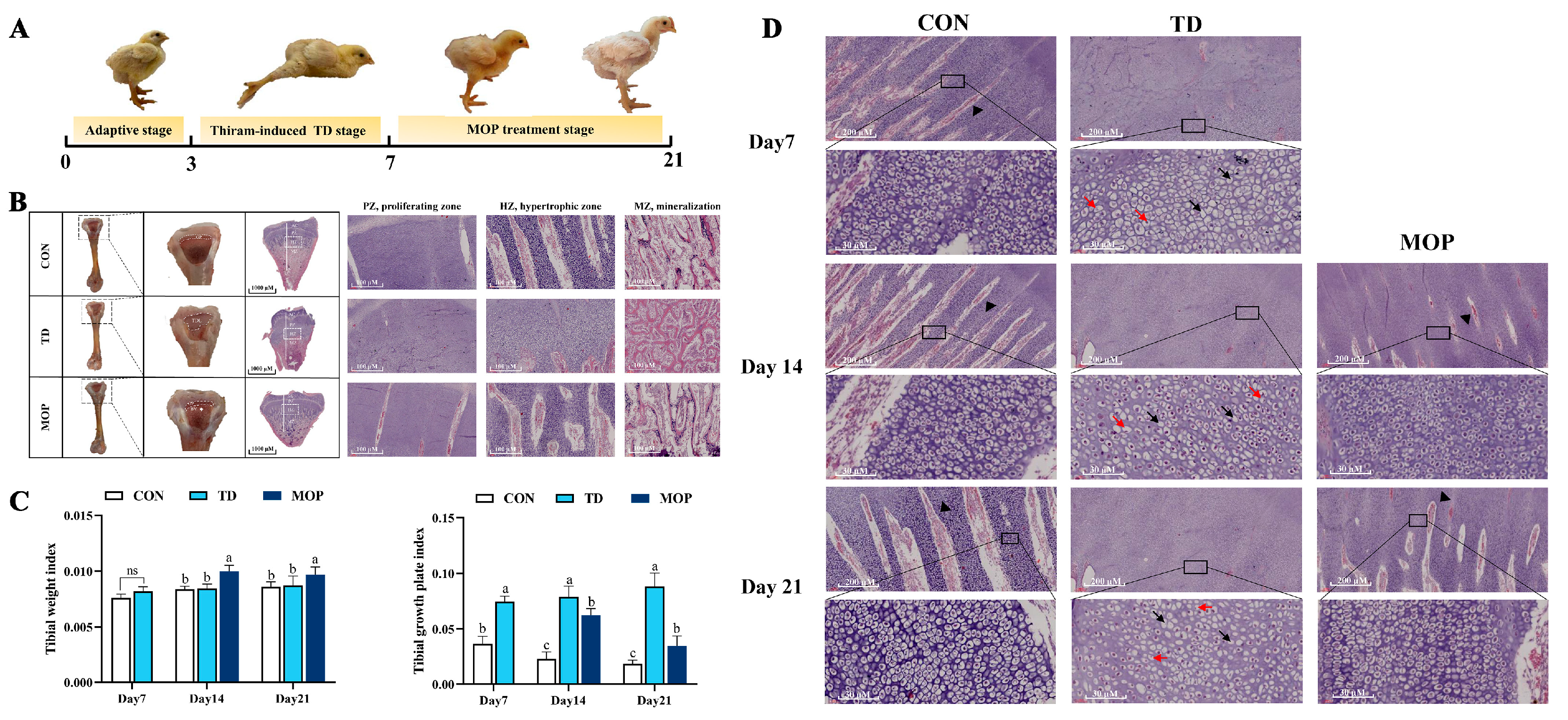
3.1. Effect of MOP on Clinical Symptoms, Tibial Parameters, Histopathology of TGPs in TD Broilers
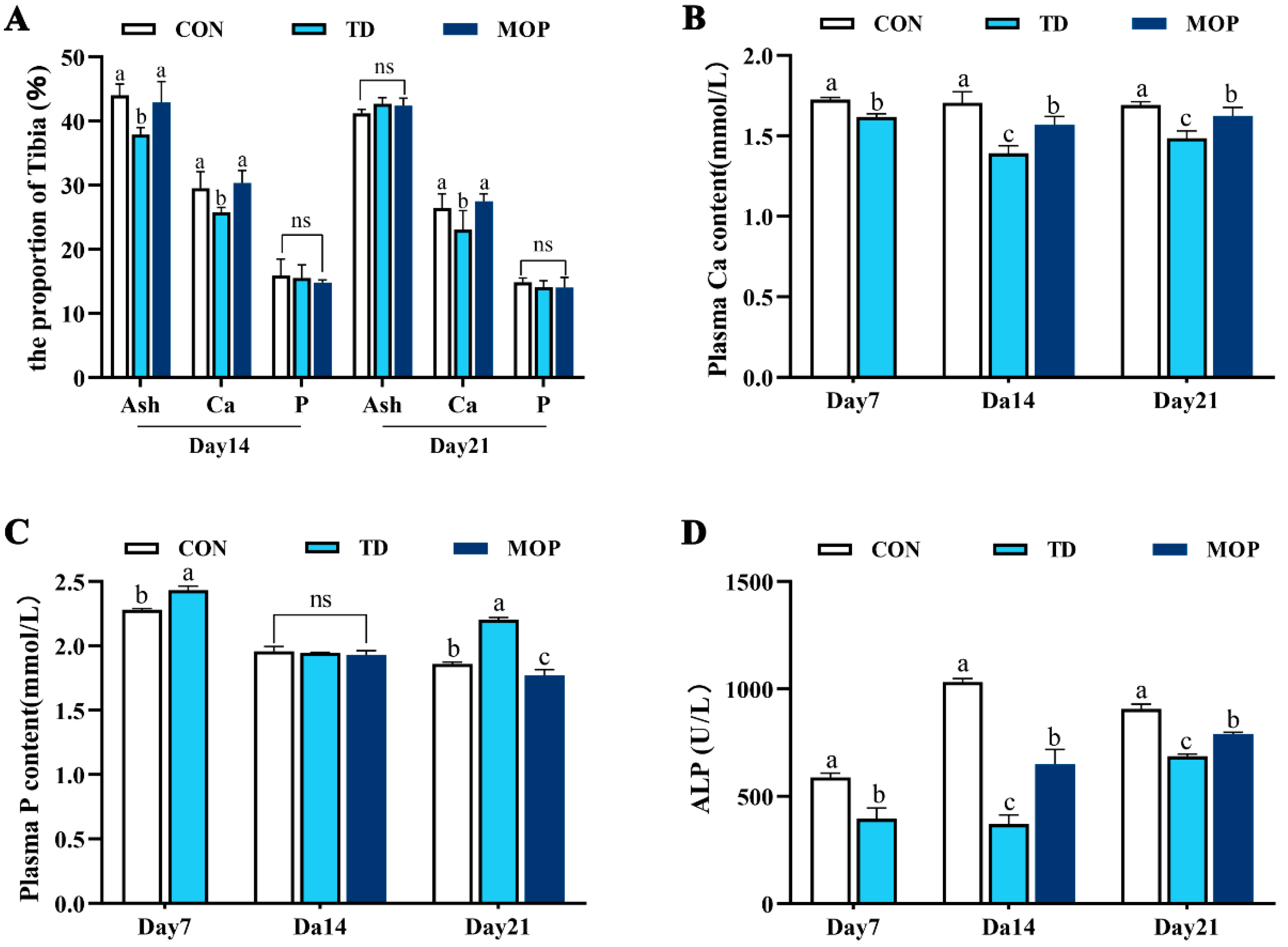
3.2. Effect of MOP on Calcium, Phosphorus, and Ash Content in Tibia and Plasma of TD Broilers
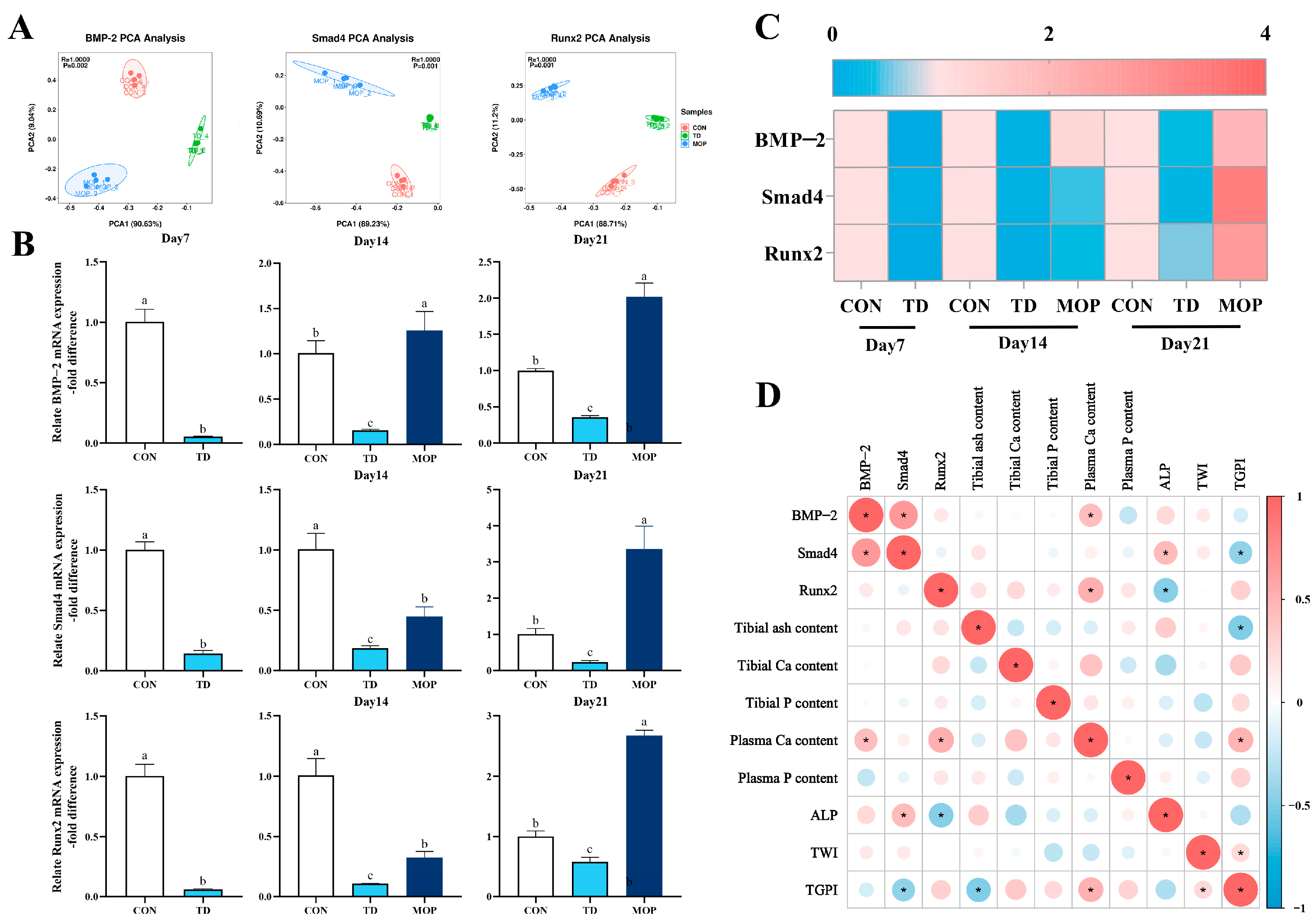
3.3. Effect of MOP on the Expression of BMP/Smads Signaling Pathway in TD Broilers
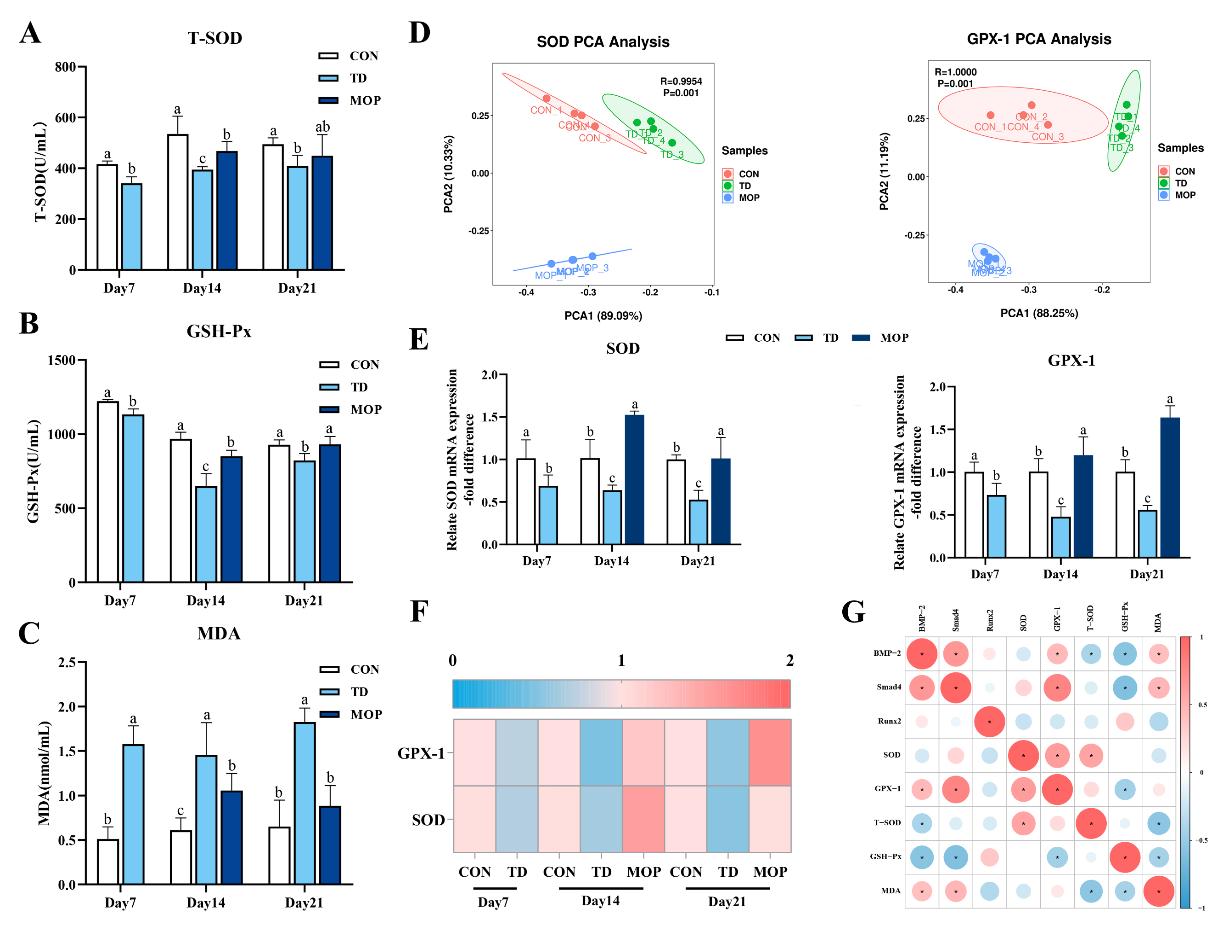
3.4. Effect of MOP on Antioxidant Activity in TD Broilers
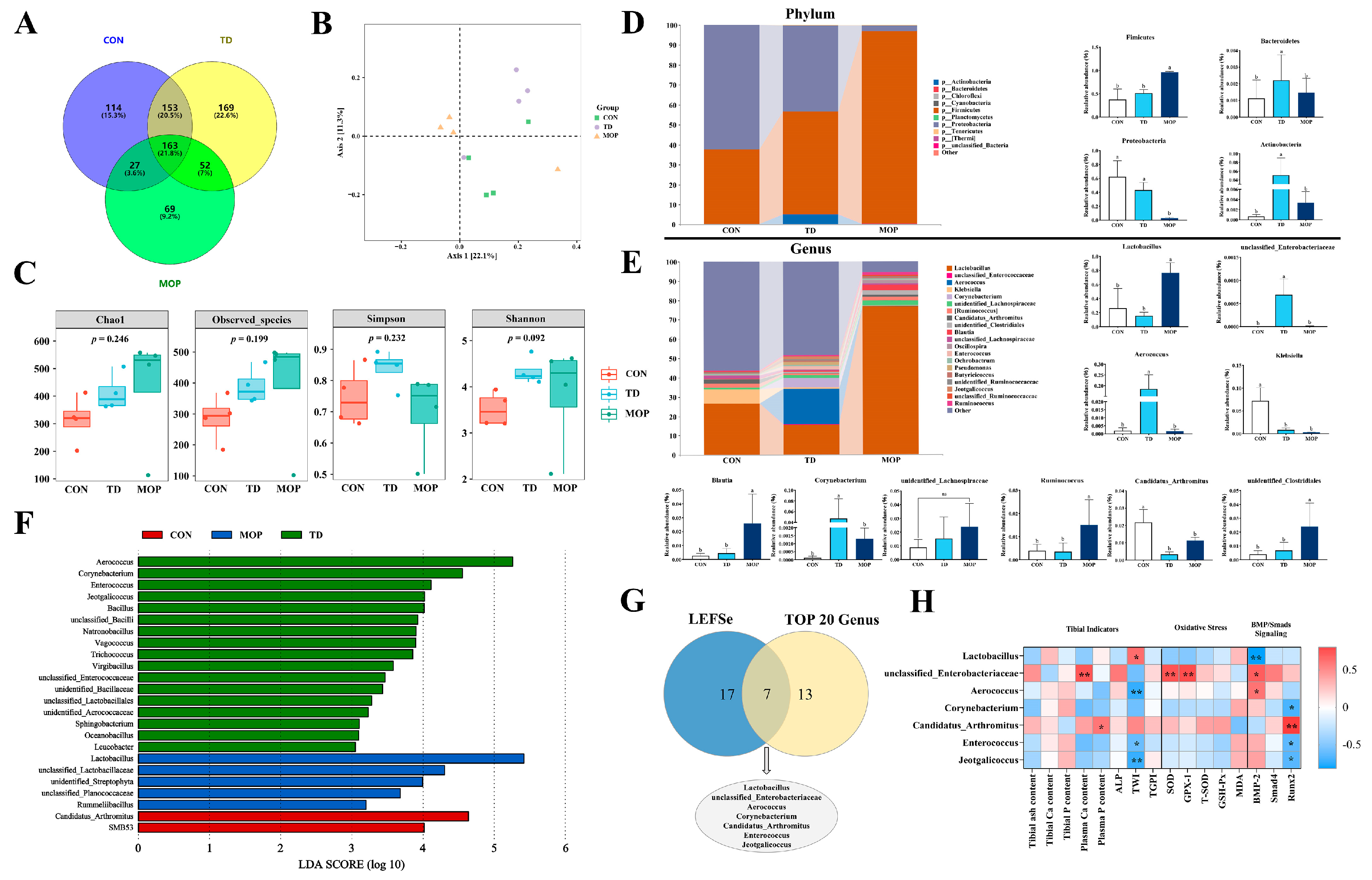
3.5. Effect of MOP on Gut Microbiota of TD Broilers
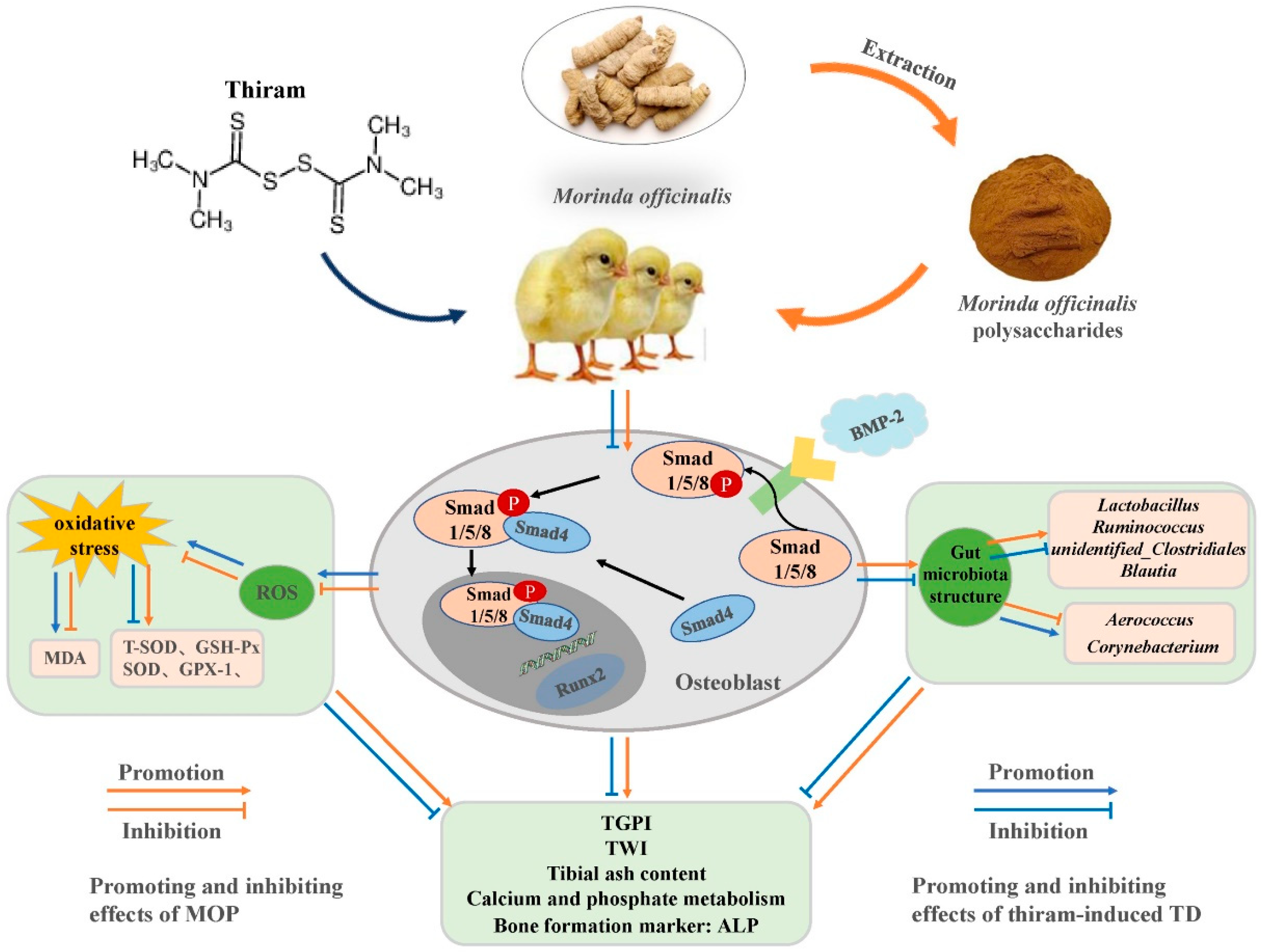
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pines, M.; Knopov, V.; Genina, O.; Hurwitz, S.; Faerman, A.; Gerstenfeld, L.C.; Leach, R.M. Development of avian tibial dyschondroplasia: Gene expression and protein synthesis. Calcif. Tissue Res. 1998, 63, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.C.; Zhang, L.H.; Rehman, M.U.; Iqbal, M.K.; Lan, Y.F.; Mehmood, K.; Zhang, H.; Qiu, G.; Nabi, F.; Yao, W.Y.; et al. High altitude hypoxia as a factor that promotes tibial growth plate development in broiler chickens. PLoS ONE 2017, 12, e0173698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waqas, M.; Wang, Y.; Li, A.; Qamar, H.; Yao, W.Y.; Tong, X.L.; Zhang, J.L.; Iqbal, M.; Mehmood, K.; Li, J.K. Osthole: A Coumarin Derivative Assuage Thiram-Induced Tibial Dyschondroplasia by Regulating BMP-2 and RUNX-2 Expressions in Chickens. Antioxidants 2019, 8, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Q.Q.; Kong, A.A.; Tao, K.S.; Zheng, S.H.; Tong, C.; Wang, X.B.; Tong, Z.X.; Rehman, M.U.; Huang, S.C. Characterization of growth performance, meat quality and serum biochemical parameters in chickens suffering from tibial dyschondroplasia. Livest. Sci. 2020, 233, 103956. [Google Scholar] [CrossRef]
- Huang, S.C.; Cao, Q.Q.; Cao, Y.B.; Yang, Y.R.; Xu, T.T.; Yue, K.; Liu, F.; Tong, Z.X.; Wang, X.B. Morinda officinalis polysaccharides improve meat quality by reducing oxidative damage in chickens suffering from tibial dyschondroplasia. Food Chem. 2021, 344, 128688. [Google Scholar] [CrossRef] [PubMed]
- Genin, O.; Hasdai, A.; Shinder, D.; Pines, M. The effect of inhibition of heat-shock proteins on thiram-induced tibial dyschondroplasia. Poult. Sci. 2012, 91, 1619–1626. [Google Scholar] [CrossRef]
- Huang, S.C.; Kong, A.A.; Cao, Q.Q.; Tong, Z.X.; Wang, X.B. The role of blood vessels in broiler chickens with tibial dyschondroplasia. Poult. Sci. 2019, 98, 6527–6532. [Google Scholar] [CrossRef]
- Shahi, M.; Peymani, A.; Sahmani, M. Regulation of Bone Metabolism. Rep. Biochem. Mol. Biol. 2017, 5, 73–82. [Google Scholar]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. RANKL biology: Bone metabolism, the immune system, and beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef] [Green Version]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef]
- Yang, X.M.; Mou, D.G.; Yu, Q.Y.; Zhang, J.M.; Xiong, Y.; Zhang, Z.M.; Xing, S. Nerve growth factor promotes osteogenic differentiation of MC3T3-E1 cells via BMP-2/Smads pathway. Ann. Anat. 2021, 239, 151819. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Tripurani, S.K.; James, R.; Pangas, S.A. Minimal fertility defects in mice deficient in oocyte-expressed Smad4. Biol. Reprod. 2012, 86, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Tu, X.; Choi, K.; Akiyama, H.; Mishina, Y.J.; Long, F.X. BMP–Smad4 signaling is required for precartilaginous mesenchymal condensation independent of Sox9 in the mouse. Dev. Biol. 2015, 400, 132–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Wu, Q.Q.; Liao, H.H.; Fan, D.; Tang, Q.Z. Bcl6 Suppresses Cardiac Fibroblast Activation and Function via Directly Binding to Smad4. Curr. Med. Sci. 2019, 39, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Javed, A.; Bae, J.S.; Afzal, F.; Gutierrez, S.; Pratap, J.; Zaidi, S.K.; Lou, Y.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Structural coupling of Smad and Runx2 for execution of the BMP2 osteogenic signal. J. Biol. Chem. 2008, 283, 8412–8422. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.Q.; Cai, H.A.; Wei, L.L.; Lan, R.X. Chitosan oligosaccharides alleviate acute heat stress-induced oxidative damage by activating ERK1/2-mediated HO-1 and GSH-Px gene expression in breast muscle of broilers. Poult. Sci. 2021, 101, 101515. [Google Scholar] [CrossRef]
- Sjogren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The gut microbiota regulates bone mass in mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, K.; Zhang, H.; Yao, W.Y.; Jiang, X.; Waqas, M.; Li, A.Y.; Wang, Y.P.; Lei, L.; Zhang, L.H.; Qamar, H.; et al. Protective effect of Astragaloside IV to inhibit thiram-induced tibial dyschondroplasia. Environ. Sci. Pollut. Res. Int. 2019, 26, 16210–16219. [Google Scholar] [CrossRef]
- Guo, L.X.; Wang, H.Y.; Liu, X.D.; Zheng, J.Y.; Tang, Q.; Wang, X.N.; Liu, J.Q.; Yin, H.Q.; Miao, B.; Liang, Y.L.; et al. Saponins from Clematis mandshurica Rupr. regulates gut microbiota and its metabolites during alleviation of collagen-induced arthritis in rats. Pharmacol. Res. 2019, 149, 104459. [Google Scholar] [CrossRef]
- Jia, X.; Jia, L.; MO, L.; Yuan, S.; Zheng, X.; He, J.; Chen, V.; Guo, Q.; Zheng, L.; Yuan, Q.; et al. Berberine Ameliorates Periodontal Bone Loss by Regulating Gut Microbiota. J. Dent. Res. 2019, 98, 107–116. [Google Scholar] [CrossRef]
- Li, B.; Liu, M.Y.; Wang, Y.; Gong, S.Q.; Yao, W.F.; Li, W.F.; Gao, H.; Wei, M.J. Puerarin improves the bone micro-environment to inhibit OVX-induced osteoporosis via modulating SCFAs released by the gut microbiota and repairing intestinal mucosal integrity. Biomed. Pharmacother. 2020, 132, 110923. [Google Scholar] [CrossRef] [PubMed]
- Isomura, H.; Fujie, K.; Shibata, K.; Inoue, N.; Iizuka, T.; Takebe, G.; Takahashi, K.; Nishihira, J.; Izumi, H.; Sakamoto, W. Bone metabolism and oxidative stress in postmenopausal rats with iron overload. Toxicology 2004, 197, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhao, L.; Liu, J.; Liu, A.L.; Zeng, W.S.; Luo, S.Q.; Bai, X.C. Hydrogen Peroxide Induces G2 Cell Cycle Arrest and Inhibits Cell Proliferation in Osteoblasts. Anat. Rec. 2010, 292, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Rivadeneira, F.; Mäkitie, O. Osteoporosis and Bone Mass Disorders: From Gene Pathways to Treatments. Trends Endocrinol. Metab. 2016, 27, 262–281. [Google Scholar] [CrossRef] [PubMed]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef]
- Kalyanaraman, H.; Ramdani, G.; Joshua, J.; Schall, N.; Boss, G.R.; Cory, E.; Sah, R.L.; Casteel, D.E.; Pilz, R.B. A Novel, Direct No Donor Regulates Osteoblast and Osteoclast Functions and Increases Bone Mass in Ovariectomized Mice. J. Bone Miner. Res. 2017, 32, 46–59. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of Mammals and Their Gut Microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; Filippis, A.D.; Xiao, H.; Quiles, J.L.; Xiao, J.B.; et al. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 24. [Google Scholar] [CrossRef]
- Lu, L.Y.; Chen, X.X.; Liu, Y.; Yu, X.J. Gut microbiota and bone metabolism. FASEB J. 2021, 35, e21740. [Google Scholar] [CrossRef]
- De Sire, A.; de Sire, R.; Curci, C.; Castiglione, F.; Wahli, W. Role of Dietary Supplements and Probiotics in Modulating Microbiota and Bone Health: The Gut-Bone Axis. Cells 2022, 11, 743. [Google Scholar] [CrossRef]
- Lian, W.S.; Wang, F.S.; Chen, Y.S.; Tsai, M.H.; Chao, H.R.; Jahr, H.; Wu, R.W.; Ko, J.Y. Gut Microbiota Ecosystem Governance of Host Inflammation, Mitochondrial Respiration and Skeletal Homeostasis. Biomedicines 2022, 10, 860. [Google Scholar] [CrossRef] [PubMed]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R. Probiotic L. reuteri treatment prevents bone loss in a menopausal ovariectomized mouse model. J. Cell Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.T.; Yue, K.; Zhang, C.D.; Tong, X.S.; Lin, L.X.; Cao, Q.Q.; Huang, S.C. Probiotics Treatment of Leg Diseases in Broiler Chickens: A Review. Probiotics Antimicrob. Proteins 2022, 14, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Wang, J.; Mehmood, K.; Chang, Y.F.; Tang, Z.; Li, Y. Treatment of tibial dyschondroplasia with traditional Chinese medicines: “Lesson and future directions”. Poult. Sci. 2020, 99, 6422–6433. [Google Scholar] [CrossRef]
- Zhang, J.H.; Xin, H.L.; Xu, Y.M.; Shen, Y.; He, Y.Q.; Lin, B.; Song, H.T.; Yang, H.Y.; Qin, L.P.; Zhang, Q.Y.; et al. Morinda officinalis How—A comprehensive review of traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2018, 213, 230–255. [Google Scholar] [CrossRef]
- Huang, S.C.; Rehman, M.U.; Lan, Y.F.; Qiu, G.; Zhang, H.; Iqbal, M.K.; Luo, H.Q.; Mehmood, K.; Zhang, L.H.; Li, J.K. Tibial dyschondroplasia is highly associated with suppression of tibial angiogenesis through regulating the HIF-1α/VEGF/VEGFR signaling pathway in chickens. Sci. Rep. UK 2017, 7, 9089. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.T.; Zheng, J.J.; Jin, W.X.; Li, L.; Lin, L.X.; Shaukat, A.; Zhang, C.D.; Cao, Q.Q.; Ashraf, M.; Huang, S.C. Total Flavonoids of Rhizoma Drynariae Ameliorate Bone Growth in Experimentally Induced Tibial Dyschondroplasia in Chickens via Regulation of OPG/RANKL Axis. Front. Pharmacol. 2022, 13, 881057. [Google Scholar] [CrossRef]
- Waqas, M.; Qamar, H.; Zhang, J.L.; Yao, W.Y.; Li, A.Y.; Wang, Y.P.; Iqbal, M.; Mehmood, K.; Jiang, X.; Li, J.K. Puerarin enhance vascular proliferation and halt apoptosis in thiram-induced avian tibial dyschondroplasia by regulating HIF-1α, TIMP-3 and BCL-2 expressions. Ecotox. Environ. Safe 2020, 190, 110126. [Google Scholar] [CrossRef]
- Tong, X.S.; Yu, G.S.; Liu, Q.Y.; Zhang, X.Q.; Bian, J.C.; Liu, Z.P.; Gu, J.H. Puerarin alleviates cadmium-induced oxidative damage to bone by reducing autophagy in rats. Environ. Toxicol. 2022, 37, 720–729. [Google Scholar] [CrossRef]
- Huang, S.C.; Zhang, L.H.; Zhang, J.L.; Rehman, M.U.; Tong, X.L.; Qiu, G.; Jiang, X.; Iqbal, M.; Shahzad, M.; Shen, Y.Q.; et al. Role and regulation of growth plate vascularization during coupling with osteogenesis in tibial dyschondroplasia of chickens. Sci. Rep. UK 2018, 8, 3680. [Google Scholar]
- Zhang, H.; Mehmood, K.; Jiang, X.; Yao, W.Y.; Iqbal, M.; Waqas, M.; Rehman, M.U.; Li, A.Y.; Shen, Y.Q.; Li, J.K. Effect of tetramethyl thiuram disulfide (thiram) in relation to tibial dyschondroplasia in chickens. Environ. Sci. Pollut. Res. Int. 2018, 25, 28264–28274. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Kong, A.A.; Fu, P.F.; Cao, Q.Q.; Tao, K.S.; Liu, D.Y.; Wang, X.B.; Tong, Z.X.; Rehman, M.U.; Huang, S.C. Lactobacillus rhamnosus JYLR-005 Prevents Thiram-Induced Tibial Dyschondroplasia by Enhancing Bone-Related Growth Performance in Chickens. Probiotics Antimicrob. Proteins 2021, 13, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Bromfield, J.I.; Hoffman, L.C.; Horyanto, D.; Soumeh, E.A. Enhancing Growth Performance, Organ Development, Meat Quality, and Bone Mineralisation of Broiler Chickens through Multi-Enzyme Super-Dosing in Reduced Energy Diets. Animals 2021, 11, 2791. [Google Scholar] [CrossRef] [PubMed]
- Saha, M.K.; Agrawal, P.; Saha, S.G.; Vishwanathan, V.; Pathak, V.; Saiprasad, S.V.; Dhariwal, P.; Dave, M. Evaluation of Correlation between Salivary Calcium, Alkaline Phosphatase and Osteoporosis- A Prospective, Comparative and Observational Study. J. Clin. Diagn. Res. 2017, 11, 63–66. [Google Scholar]
- Sun, W.P.; Li, M.Y.; Zhang, Y.; Huang, Y.J.; Zhan, Q.Z.; Ren, Y.Y.; Dong, H.; Chen, J.N.; Li, Z.G.; Fan, C.; et al. Total flavonoids of rhizoma drynariae ameliorates bone formation and mineralization in BMP-Smad signaling pathway induced large tibial defect rats. Biomed. Pharmacother. 2021, 138, 111480. [Google Scholar] [CrossRef]
- Liu, M.J.; Xiao, G.G.; Rong, P.J.; Dong, J.Z.; Zhang, Z.G.; Zhao, H.Y.; Teng, J.G.; Zhao, H.X.; Pan, J.H.; Li, Y.; et al. Semen Astragali Complanati- and Rhizoma Cibotii-enhanced bone formation in osteoporosis rats. BMC Complement. Altern. Med. 2013, 13, 141. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.Y.; Huang, D.; Shen, X.; Qin, N.B.; Jiang, K.M.; Zhang, D.W.; Zhang, Q. Identification and characterization of a polysaccharide from the roots of Morinda officinalis, as an inducer of bone formation by up-regulation of target gene expression. Int. J. Biol. Macromol. 2019, 133, 446–456. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B.; Wijnen, A.J.; Stein, J.L.; Montecino, M.; Javed, A.; Zaidi, S.K.; Young, D.W.; Choi, J.Y.; Pockwinse, S.M. Runx2 control of organization, assembly and activity of the regulatory machinery for skeletal gene expression. Oncogenesis 2004, 23, 4315–4329. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.W.; Fan, D.Y.; Yuan, W.Q. CMTM3 suppresses bone formation and osteogenic differentiation of mesenchymal stem cells through inhibiting Erk1/2 and RUNX2 pathways. Genes Dis. 2021, 8, 882–890. [Google Scholar] [CrossRef]
- Zhou, Q.Z.; Zhu, L.; Zhang, D.; Li, N.; Li, Q.; Dai, P.P.; Mao, Y.X.; Li, X.M.; Ma, J.F.; Huang, S.B. Oxidative Stress-Related Biomarkers in Postmenopausal Osteoporosis: A Systematic Review and Meta-Analyses. Dis. Markers 2016, 2016, 7067984. [Google Scholar] [CrossRef] [Green Version]
- Zahan, O.M.; Serban, O.; Gherman, C.; Fodor, D. The evaluation of oxidative stress in osteoarthritis. Med. Pharm. Rep. 2020, 93, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Mccord, J.M.; Edeas, M.A. SOD, oxidative stress and human pathologies: A brief history and a future vision. Biomed. Pharmacother. 2005, 59, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Mikail, M.A.; Ahmed, I.A.; Hazali, N.; Abdul Rasad, M.S.B.; Abdul Ghani, R.; Hashim, R.; Arief, S.J.; Md Isa, M.L.; Draman, S. Comparison of the effects of three different Baccaurea angulata whole fruit juice doses on plasma, aorta and liver MDA levels, antioxidant enzymes and total antioxidant capacity. Eur. J. Nutr. 2018, 57, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Cha, X.X.; Han, S.H.; Jian, J.Y.; Zhang, S.Y.; Yu, S.; Fu, D.J.; Yao, M.; Zhang, L.R.; Feng, G.Q. Inulin with a low degree of polymerization protects human umbilical vein endothelial cells from hypoxia/reoxygenation-induced injury. Carbohydr. Polym. 2019, 216, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.L.; Liu, L.L.; Chen, Y.X.; Ma, H.; Li, M.; Qu, W.S.; Yin, J.Y.; Zhang, X.; Gao, Y.T.; Shan, J.J.; et al. The chemical character of polysaccharides from processed Morindae officinalis and their effects on anti-liver damage. Int. J. Biol. Macromol. 2019, 141, 410–421. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Fang, L.H.; Liu, X.P.; Li, R.; Nie, T.T.; Gao, C.H. Effects of LncRNA SNHG15 on Osteogenic/Adipogenic Differentiation of Bone Marrow Mesenchymal Stem Cells Under Oxidative Stress by TGFβ/Smad Signaling Pathway. J. Biomater. Tissue Eng. 2019, 9, 1273–1278. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, M.; Park, M.H. Diphlorethohydroxycamalol isolated from Ishige okamurae prevents H2O2-induced oxidative damage via BMP2/Runx2 signaling in osteoblastic MC3T3-E1 cells. Fitoterapia 2021, 152, 104921. [Google Scholar] [CrossRef]
- Yeoman, C.J.; Chia, N.; Jeraldo, P.; Sipos, M.; Goldenfeld, N.D.; White, B.A. The microbiome of the chicken gastrointestinal tract. Anim. Health Res. Rev. 2012, 13, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.C.; Xue, J.; Song, H.; Du, Q. Regulatory effects of traditional Chinese medicine on intestinal flora. J. Chin. Pharm. Sci. 2020, 29, 161–175. [Google Scholar]
- Kim, J.-S.; Park, J.-E.; Lee, K.C.; Choi, S.-H.; Oh, B.S.; Yu, S.Y.; Eom, M.K.; Kang, S.W.; Han, K.-I.; Suh, M.K.; et al. Blautia faecicola sp. nov., isolated from faeces from a healthy human. Int. J. Syst. Evol. Microbiol. 2020, 70, 2059–2065. [Google Scholar] [CrossRef]
- Pal, D.; Naskar, M.; Bera, A.; Mukhopadhyay, B. Chemical synthesis of the pentasaccharide repeating unit of the O-specific polysaccharide from Ruminococcus gnavus. Carbohyd. Res. 2021, 507, 108384. [Google Scholar] [CrossRef] [PubMed]
- Schnupf, P.; Gaboriau-Routhiau, V.; Cerf-Bensussan, N. Host interactions with Segmented Filamentous Bacteria: An unusual trade-off that drives the post-natal maturation of the gut immune system. Seminin. Immunol. 2013, 25, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Roux, V.; Drancourt, M.; Stein, A.; Riegel, P.; Raoult, D.; La Scola, B. Corynebacterium Species Isolated from Bone and Joint Infections Identified by 16S rRNA Gene Sequence Analysis. J. Clin. Microbiol. 2004, 42, 2231–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.Y.; Chan, Y.C.; Wong, C.Y. Corynebacterium striatum as an emerging pathogen. J. Infect. Dev. Ctries 2010, 12, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M. Aerococcus: An increasingly acknowledged human pathogen. Clin. Microbiol. Infect. 2016, 22, 22–27. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, % | Content |
|---|---|
| Corn | 57.0 |
| Soybean meal | 31.5 |
| Corn gluten meal | 3.4 |
| Soybean oil | 3.1 |
| Limestone | 1.2 |
| Dicalcium phosphate | 2.0 |
| L-Lysine | 0.3 |
| DL-Methionine | 0.2 |
| Sodium chloride | 0.3 |
| Premix 1 | 1.0 |
| Total | 100.0 |
| Calculated nutrient levels | |
| Apparent metabolizable energy, MJ/kg | 12.6 |
| Crude protein, % | 21.3 |
| Calcium, % | 1.0 |
| Total phosphorus, % | 0.7 |
| Available phosphorus, % | 0.5 |
| Lysine, % | 1.2 |
| Methionine, % | 0.5 |
| Methionine + cystine, % | 0.9 |
| Analyzed nutrient levels 2 | |
| Gross energy, MJ/kg | 15.5 |
| Crude protein, % | 20.8 |
| Calcium, % | 1.1 |
| Total phosphorus, % | 0.7 |
| Lysine, % | 1.2 |
| Methionine, % | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Xu, T.; Lin, L.; Shaukat, A.; Tong, X.; Yue, K.; Cao, Q.; Zhang, C.; Liu, F.; Huang, S. Morinda officinalis Polysaccharides Ameliorates Bone Growth by Attenuating Oxidative Stress and Regulating the Gut Microbiota in Thiram-Induced Tibial Dyschondroplasia Chickens. Metabolites 2022, 12, 958. https://doi.org/10.3390/metabo12100958
Zhang C, Xu T, Lin L, Shaukat A, Tong X, Yue K, Cao Q, Zhang C, Liu F, Huang S. Morinda officinalis Polysaccharides Ameliorates Bone Growth by Attenuating Oxidative Stress and Regulating the Gut Microbiota in Thiram-Induced Tibial Dyschondroplasia Chickens. Metabolites. 2022; 12(10):958. https://doi.org/10.3390/metabo12100958
Chicago/Turabian StyleZhang, Chaodong, Tingting Xu, Luxi Lin, Aftab Shaukat, Xishuai Tong, Ke Yue, Qinqin Cao, Cai Zhang, Fang Liu, and Shucheng Huang. 2022. "Morinda officinalis Polysaccharides Ameliorates Bone Growth by Attenuating Oxidative Stress and Regulating the Gut Microbiota in Thiram-Induced Tibial Dyschondroplasia Chickens" Metabolites 12, no. 10: 958. https://doi.org/10.3390/metabo12100958
APA StyleZhang, C., Xu, T., Lin, L., Shaukat, A., Tong, X., Yue, K., Cao, Q., Zhang, C., Liu, F., & Huang, S. (2022). Morinda officinalis Polysaccharides Ameliorates Bone Growth by Attenuating Oxidative Stress and Regulating the Gut Microbiota in Thiram-Induced Tibial Dyschondroplasia Chickens. Metabolites, 12(10), 958. https://doi.org/10.3390/metabo12100958