
Harmonization of Rapid Evaporative Ionization Mass Spectrometry Workflows across Four Sites and Testing Using Reference Material and Local Food-Grade Meats

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
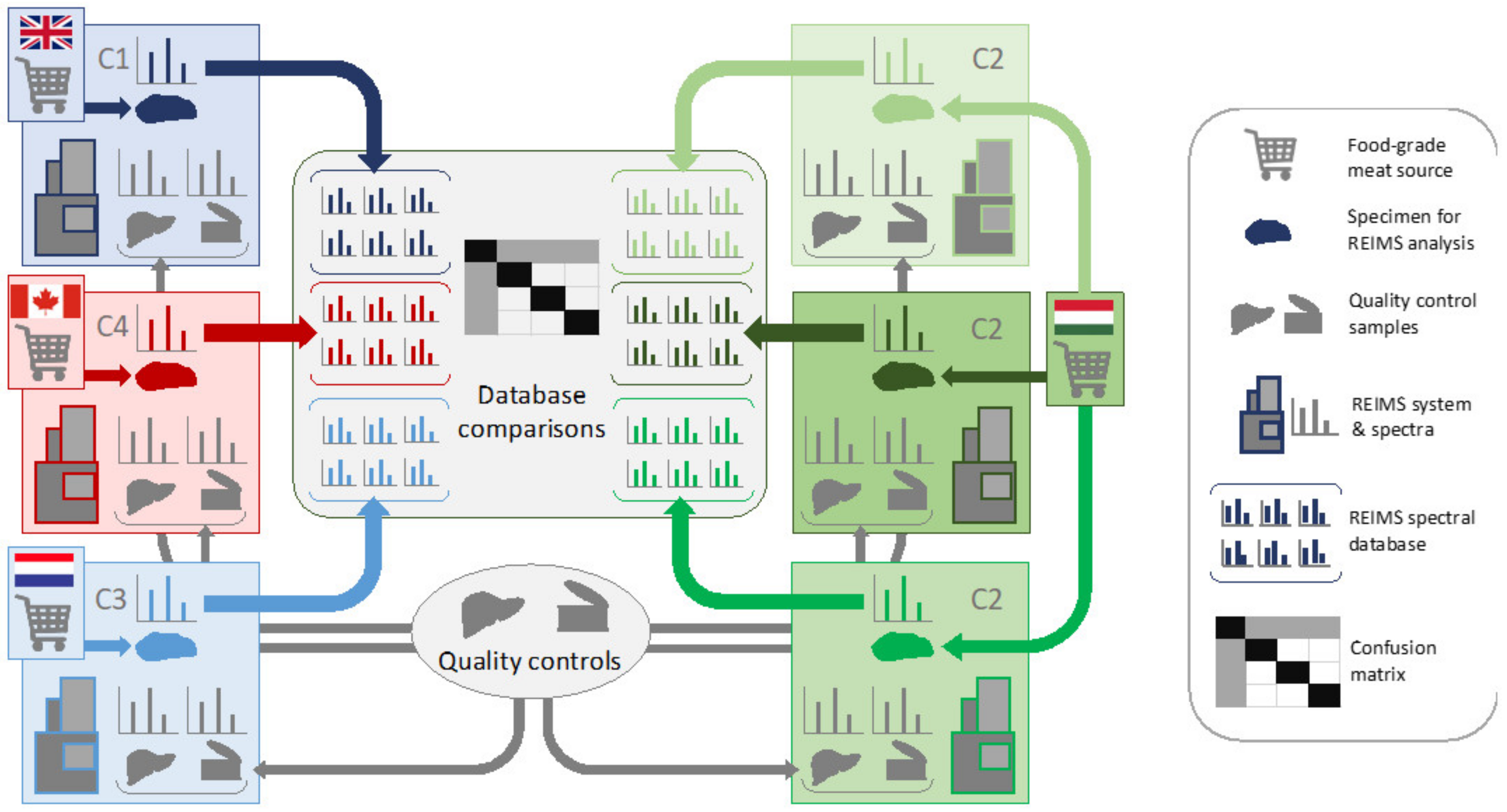
2.1. Samples and Logistics
2.2. Sampling with Diathermic Knife
2.3. REIMS—Q-TOF Instrumentation
2.4. Data Analysis
3. Results
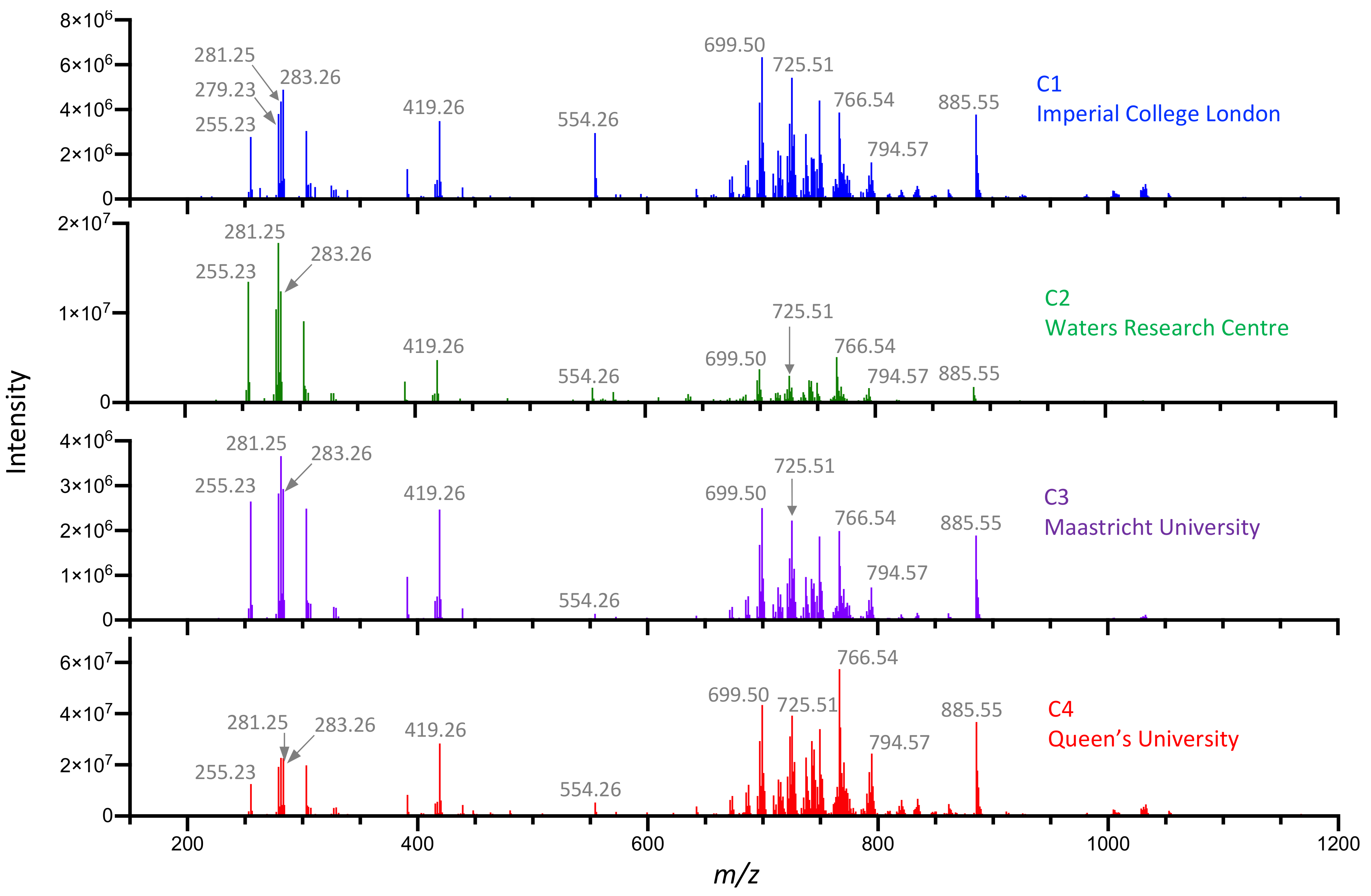
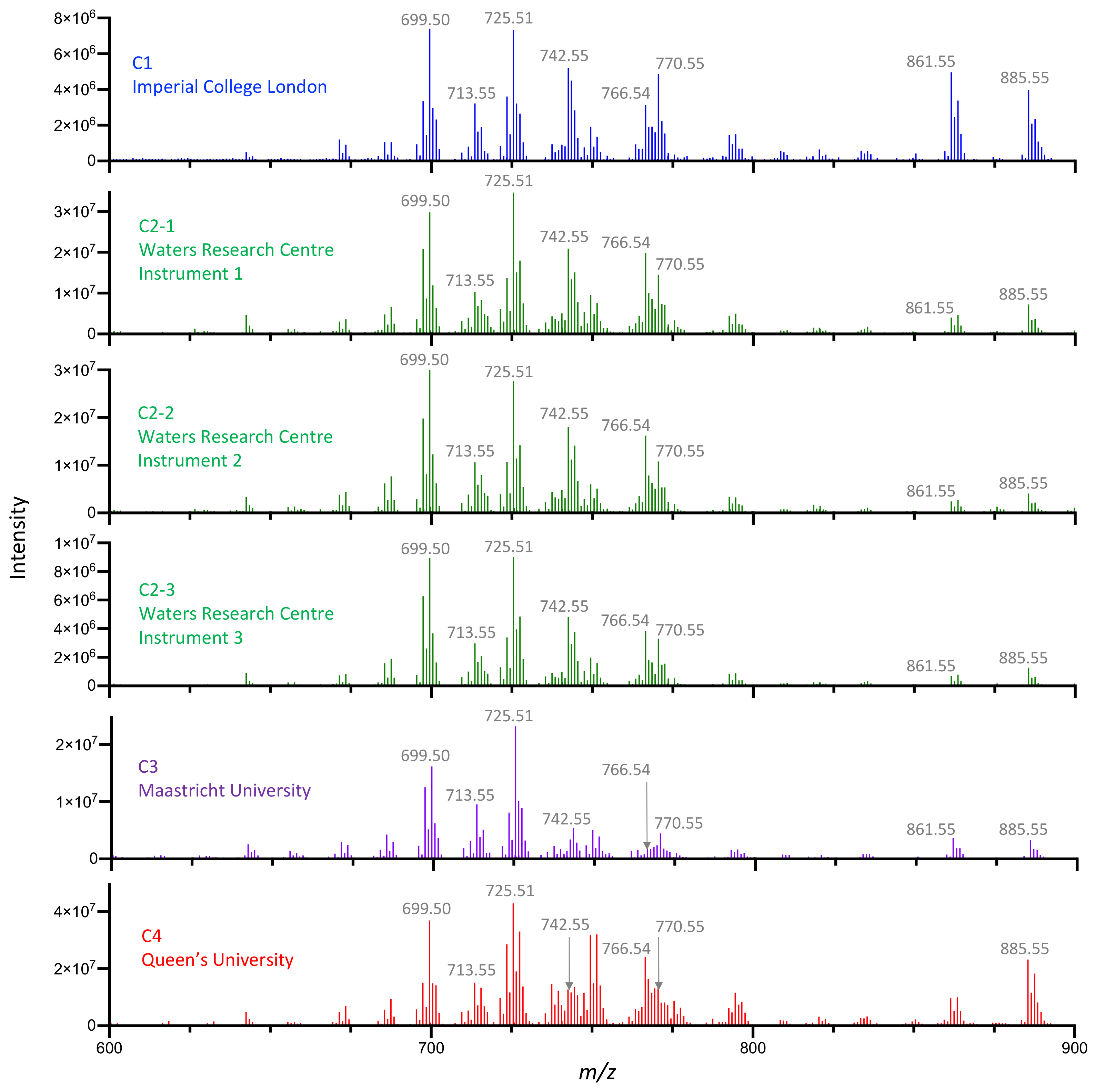
3.1. Multi-Center Characterization of Pork Liver and NIST Meat Homogenate
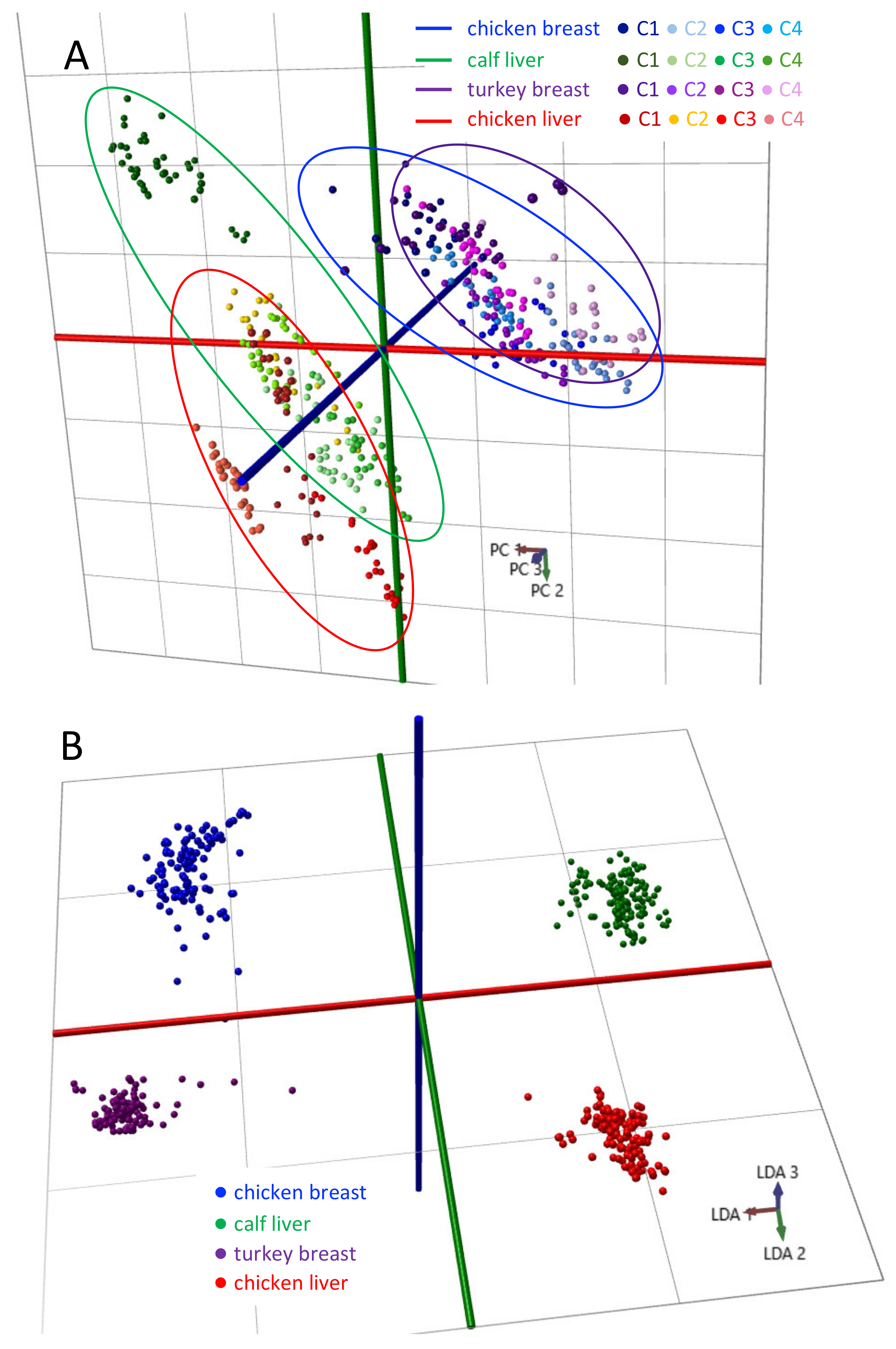
3.2. Multi-Site Classification of Local Food-Grade Meats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balog, J.; Sasi-Szabó, L.; Kinross, J.; Lewis, M.R.; Muirhead, L.J.; Veselkov, K.; Mirnezami, R.; Dezső, B.; Damjanovich, L.; Darzi, A.; et al. Intraoperative Tissue Identification Using Rapid Evaporative Ionization Mass Spectrometry. Sci. Transl. Med. 2013, 5, 194ra93. [Google Scholar] [CrossRef] [PubMed]
- Vaysse, P.-M.; Heeren, R.M.A.; Porta, T.; Balluff, B. Mass Spectrometry Imaging for Clinical Research—Latest Developments, Applications, and Current Limitations. Analyst 2017, 142, 2690–2712. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, K.-C.; Dénes, J.; Albrecht, K.; Szaniszló, T.; Balog, J.; Skoumal, R.; Katona, M.; Tóth, M.; Balogh, L.; Takáts, Z. In Vivo, in Situ Tissue Analysis Using Rapid Evaporative Ionization Mass Spectrometry. Angew. Chem. Int. Ed. Engl. 2009, 48, 8240–8242. [Google Scholar] [CrossRef] [PubMed]
- Strittmatter, N.; Lovrics, A.; Sessler, J.; McKenzie, J.S.; Bodai, Z.; Doria, M.L.; Kucsma, N.; Szakacs, G.; Takats, Z. Shotgun Lipidomic Profiling of the NCI60 Cell Line Panel Using Rapid Evaporative Ionization Mass Spectrometry. Anal. Chem. 2016, 88, 7507–7514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Rabie, P.; Sheelan, D.; Laures, A.; Spaull, J.; Dowell, S. Increasing the Discrimination Power of Rapid Evaporative Ionisation Mass Spectrometry (REIMS) in Analytical Control Tissue Quality Screening and Cell Line Sample Identification. Rapid Commun. Mass Spectrom. 2019, 35, e8525. [Google Scholar] [CrossRef]
- Cameron, S.J.S.; Bolt, F.; Perdones-Montero, A.; Rickards, T.; Hardiman, K.; Abdolrasouli, A.; Burke, A.; Bodai, Z.; Karancsi, T.; Simon, D.; et al. Rapid Evaporative Ionisation Mass Spectrometry (REIMS) Provides Accurate Direct from Culture Species Identification within the Genus Candida. Sci. Rep. 2016, 6, 36788. [Google Scholar] [CrossRef] [Green Version]
- Strittmatter, N.; Jones, E.A.; Veselkov, K.A.; Rebec, M.; Bundy, J.G.; Takats, Z. Analysis of Intact Bacteria Using Rapid Evaporative Ionisation Mass Spectrometry. Chem. Commun. 2013, 49, 6188–6190. [Google Scholar] [CrossRef]
- Strittmatter, N.; Rebec, M.; Jones, E.A.; Golf, O.; Abdolrasouli, A.; Balog, J.; Behrends, V.; Veselkov, K.A.; Takats, Z. Characterization and Identification of Clinically Relevant Microorganisms Using Rapid Evaporative Ionization Mass Spectrometry. Anal. Chem. 2014, 86, 6555–6562. [Google Scholar] [CrossRef]
- Verplanken, K.; Stead, S.; Jandova, R.; Poucke, C.V.; Claereboudt, J.; Bussche, J.V.; Saeger, S.D.; Takats, Z.; Wauters, J.; Vanhaecke, L. Rapid Evaporative Ionization Mass Spectrometry for High-Throughput Screening in Food Analysis: The Case of Boar Taint. Talanta 2017, 169, 30–36. [Google Scholar] [CrossRef]
- Rigano, F.; Mangraviti, D.; Stead, S.; Martin, N.; Petit, D.; Dugo, P.; Mondello, L. Rapid Evaporative Ionization Mass Spectrometry Coupled with an Electrosurgical Knife for the Rapid Identification of Mediterranean Sea Species. Anal. Bioanal. Chem. 2019, 411, 6603–6614. [Google Scholar] [CrossRef]
- Guitton, Y.; Dervilly-Pinel, G.; Jandova, R.; Stead, S.; Takats, Z.; Le Bizec, B. Rapid Evaporative Ionisation Mass Spectrometry and Chemometrics for High-Throughput Screening of Growth Promoters in Meat Producing Animals. Food Addit. Contam. Part A 2018, 35, 900–910. [Google Scholar] [CrossRef]
- Balog, J.; Perenyi, D.; Guallar-Hoyas, C.; Egri, A.; Pringle, S.D.; Stead, S.; Chevallier, O.P.; Elliott, C.T.; Takats, Z. Identification of the Species of Origin for Meat Products by Rapid Evaporative Ionization Mass Spectrometry. J. Agric. Food Chem. 2016, 64, 4793–4800. [Google Scholar] [CrossRef]
- St John, E.R.; Balog, J.; McKenzie, J.S.; Rossi, M.; Covington, A.; Muirhead, L.; Bodai, Z.; Rosini, F.; Speller, A.V.M.; Shousha, S.; et al. Rapid Evaporative Ionisation Mass Spectrometry of Electrosurgical Vapours for the Identification of Breast Pathology: Towards an Intelligent Knife for Breast Cancer Surgery. Breast Cancer Res. 2017, 19, 59. [Google Scholar] [CrossRef] [Green Version]
- Balog, J.; Kumar, S.; Alexander, J.; Golf, O.; Huang, J.; Wiggins, T.; Abbassi-Ghadi, N.; Enyedi, A.; Kacska, S.; Kinross, J.; et al. In Vivo Endoscopic Tissue Identification by Rapid Evaporative Ionization Mass Spectrometry (REIMS). Angew. Chem. Int. Ed. Engl. 2015, 54, 11059–11062. [Google Scholar] [CrossRef]
- Phelps, D.L.; Balog, J.; Gildea, L.F.; Bodai, Z.; Savage, A.; El-Bahrawy, M.A.; Speller, A.V.; Rosini, F.; Kudo, H.; McKenzie, J.S.; et al. The Surgical Intelligent Knife Distinguishes Normal, Borderline and Malignant Gynaecological Tissues Using Rapid Evaporative Ionisation Mass Spectrometry (REIMS). Br. J. Cancer 2018, 118, 1349–1358. [Google Scholar] [CrossRef] [Green Version]
- Tzafetas, M.; Mitra, A.; Paraskevaidi, M.; Bodai, Z.; Kalliala, I.; Bowden, S.; Lathouras, K.; Rosini, F.; Szasz, M.; Savage, A.; et al. The Intelligent Knife (IKnife) and Its Intraoperative Diagnostic Advantage for the Treatment of Cervical Disease. Proc. Natl. Acad. Sci. USA 2020, 117, 7338–7346. [Google Scholar] [CrossRef] [Green Version]
- Glunde, K.; Jie, C.; Bhujwalla, Z.M. Molecular Causes of the Aberrant Choline Phospholipid Metabolism in Breast Cancer. Cancer Res. 2004, 64, 4270–4276. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.A.; Simon, D.; Karancsi, T.; Balog, J.; Pringle, S.D.; Takats, Z. Matrix Assisted Rapid Evaporative Ionization Mass Spectrometry. Anal. Chem. 2019, 91, 9784–9791. [Google Scholar] [CrossRef]
- Genangeli, M.; Heeren, R.M.A.; Porta Siegel, T. Tissue Classification by Rapid Evaporative Ionization Mass Spectrometry (REIMS): Comparison between a Diathermic Knife and CO2 Laser Sampling on Classification Performance. Anal. Bioanal. Chem. 2019, 411, 7943–7955. [Google Scholar] [CrossRef] [Green Version]
- Gredell, D.A.; Schroeder, A.R.; Belk, K.E.; Broeckling, C.D.; Heuberger, A.L.; Kim, S.-Y.; King, D.A.; Shackelford, S.D.; Sharp, J.L.; Wheeler, T.L.; et al. Comparison of Machine Learning Algorithms for Predictive Modeling of Beef Attributes Using Rapid Evaporative Ionization Mass Spectrometry (REIMS) Data. Sci. Rep. 2019, 9, 5721. [Google Scholar] [CrossRef]
- Ross, A.; Brunius, C.; Chevallier, O.; Dervilly, G.; Elliott, C.; Guitton, Y.; Prenni, J.E.; Savolainen, O.; Hemeryck, L.; Vidkjær, N.H.; et al. Making Complex Measurements of Meat Composition Fast: Application of Rapid Evaporative Ionisation Mass Spectrometry to Measuring Meat Quality and Fraud. Meat Sci. 2021, 181, 108333. [Google Scholar] [CrossRef] [PubMed]
- Robson, K.; Birse, N.; Chevallier, O.; Elliott, C. Metabolomic Profiling to Detect Different Forms of Beef Fraud Using Rapid Evaporative Ionisation Mass Spectrometry (REIMS). NPJ Sci. Food. 2022, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ross, A.B.; Yoo, M.J.Y.; Farouk, M.M. Use of Rapid Evaporative Ionisation Mass Spectrometry Fingerprinting to Determine the Metabolic Changes to Dry-Aged Lean Beef Due to Different Ageing Regimes. Meat Sci. 2021, 181, 108438. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Yang, M.; Chen, X.; Yan, X.; Li, Y.; He, M.; Liu, T.; Chen, F.; Zhang, F. Differentiation between Fresh and Frozen-Thawed Meat Using Rapid Evaporative Ionization Mass Spectrometry: The Case of Beef Muscle. J. Agric. Food Chem. 2021, 69, 5709–5724. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Used for Training ↓ | Correct Classification Rate (%) | ||||||
|---|---|---|---|---|---|---|---|
| Total | C1 | C2-1 | C2-2 | C2-3 | C3 | C4 | |
| C1 | 88.24 | - | 71.43 | 100.0 | 75.0 | 93.1 | 100.0 |
| C2 (all) 1 | 100.0 | 100.0 | - | - | - | 100.0 | 100.0 |
| C2-1 | 98.17 | 98.2 | - | 100.0 | 100.0 | 97.0 | 98.2 |
| C3 | 88.9 | 100.0 | 76.19 | 50.0 | 85.7 | - | 100.0 |
| C4 | 69.9 | 83.2 | 59.0 | 50.0 | 50.0 | 77.2 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaufmann, M.; Vaysse, P.-M.; Savage, A.; Amgheib, A.; Marton, A.; Manoli, E.; Fichtinger, G.; Pringle, S.D.; Rudan, J.F.; Heeren, R.M.A.; et al. Harmonization of Rapid Evaporative Ionization Mass Spectrometry Workflows across Four Sites and Testing Using Reference Material and Local Food-Grade Meats. Metabolites 2022, 12, 1130. https://doi.org/10.3390/metabo12111130
Kaufmann M, Vaysse P-M, Savage A, Amgheib A, Marton A, Manoli E, Fichtinger G, Pringle SD, Rudan JF, Heeren RMA, et al. Harmonization of Rapid Evaporative Ionization Mass Spectrometry Workflows across Four Sites and Testing Using Reference Material and Local Food-Grade Meats. Metabolites. 2022; 12(11):1130. https://doi.org/10.3390/metabo12111130
Chicago/Turabian StyleKaufmann, Martin, Pierre-Maxence Vaysse, Adele Savage, Ala Amgheib, András Marton, Eftychios Manoli, Gabor Fichtinger, Steven D. Pringle, John F. Rudan, Ron M. A. Heeren, and et al. 2022. "Harmonization of Rapid Evaporative Ionization Mass Spectrometry Workflows across Four Sites and Testing Using Reference Material and Local Food-Grade Meats" Metabolites 12, no. 11: 1130. https://doi.org/10.3390/metabo12111130
APA StyleKaufmann, M., Vaysse, P.-M., Savage, A., Amgheib, A., Marton, A., Manoli, E., Fichtinger, G., Pringle, S. D., Rudan, J. F., Heeren, R. M. A., Takáts, Z., Balog, J., & Porta Siegel, T. (2022). Harmonization of Rapid Evaporative Ionization Mass Spectrometry Workflows across Four Sites and Testing Using Reference Material and Local Food-Grade Meats. Metabolites, 12(11), 1130. https://doi.org/10.3390/metabo12111130