
Metabolomic Profiles in Childhood and Adolescence Are Associated with Fetal Overnutrition


 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Characteristics
2.2. Identification of Metabolites in Offspring Associated with Any Fetal Overnutrition
2.3. Associations between Fetal Overnutrition Typologies and Offspring Metabolite Profiles
Sensitivity Analyses
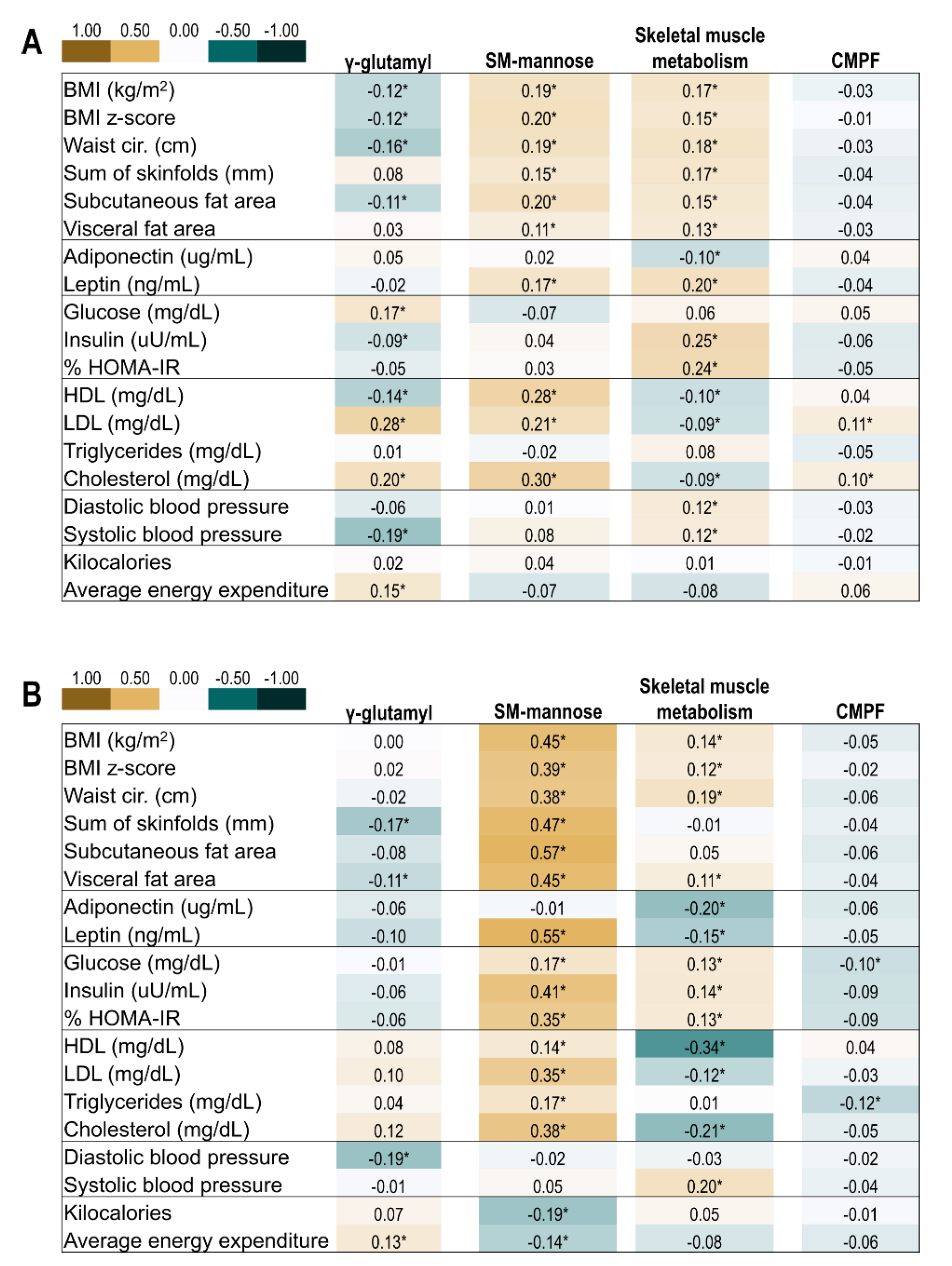
2.4. Correlation of Offspring Metabolite Profiles and Indicators of Metabolic Health
3. Discussion
3.1. Summary of Overall Findings
3.2. γ-Glutamyl Peptides Factor
3.3. Sphingomyelin-Mannose Factor
3.4. Skeletal Muscle Metabolism Factor
3.5. CMPF Factor
3.6. Strengths and Limitations
3.7. Conclusion and Future Direction
4. Materials and Methods
4.1. Study Population
4.2. Assessment of Exposure to Fetal Overnutrition
4.2.1. Gestational Diabetes Mellitus
4.2.2. Obesity
4.2.3. Typology of Fetal Overnutrition
4.3. Assessment of Metabolite Profiles in Offspring
4.4. Assessment of Conventional Biomarkers of Metabolic Risk in Offspring
4.4.1. Biomarkers
4.4.2. Anthropometric and Body Composition
4.4.3. Lifestyle Behaviors
4.5. Covariates
4.6. Statistical Analysis
4.6.1. Identification of Offspring Metabolites Associated with Any Fetal Overnutrition
4.6.2. Associations of Fetal Overnutrition Typology with Offspring Metabolite Profiles
4.6.3. Correlation of Offspring Metabolite Profiles and Indicators of Metabolic Health and Lifestyle
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perng, W.; Oken, E.; Dabelea, D. Developmental overnutrition and obesity and type 2 diabetes in offspring. Diabetologia 2019, 62, 1779–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; England, L.; Wilson, H.G.; Bish, C.; Satten, G.A.; Dietz, P. Percentage of gestational diabetes mellitus attributable to overweight and obesity. Am. J. Public Health 2010, 100, 1047–1052. [Google Scholar] [CrossRef]
- Clausen, T.D.; Mathiesen, E.R.; Hansen, T.; Pedersen, O.; Jensen, D.M.; Lauenborg, J.; Damm, P. High prevalence of type 2 diabetes and pre-diabetes in adult offspring of women with gestational diabetes mellitus or type 1 diabetes. Diabetes Care 2008, 31, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, W.L., Jr.; Scholtens, D.M.; Lowe, L.P.; Kuang, A.; Nodzenski, M.; Talbot, O.; Catalano, P.M.; Linder, B.; Brickman, W.J.; Clayton, P.; et al. Association of gestational diabetes with maternal disorders of glucose metabolism and childhood adiposity. JAMA 2018, 320, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Razaz, N.; Villamor, E.; Muraca, G.M.; Bonamy, A.-K.E.; Cnattingius, S. Maternal obesity and risk of cardiovascular diseases in offspring: A population-based cohort and sibling-controlled study. Lancet Diabetes Endocrinol. 2020, 8, 572–581. [Google Scholar] [CrossRef]
- Hivert, M.F.; Perng, W.; Watkins, S.M.; Newgard, C.S.; Kenny, L.C.; Kristal, B.S.; Patti, M.E.; Isganaitis, E.; DeMeo, D.L.; Oken, E.; et al. Metabolomics in the developmental origins of obesity and its cardiometabolic consequences. J. Dev. Orig. Health Dis. 2015, 6, 65–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, W.L., Jr.; Bain, J.R.; Nodzenski, M.; Reisetter, A.C.; Muehlbauer, M.J.; Stevens, R.D.; Ilkayeva, O.R.; Lowe, L.P.; Metzger, B.E.; Newgard, C.B.; et al. Maternal BMI and glycemia impact the fetal metabolome. Diabetes Care 2017, 40, 902–910. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.L.; Doyon, M.; Arguin, M.; Perron, P.; Bouchard, L.; Hivert, M.-F. A prospective study of maternal adiposity and glycemic traits across pregnancy and mid-childhood metabolomic profiles. Int. J. Obes. 2021, 45, 860–869. [Google Scholar] [CrossRef]
- Perng, W.; Ringham, B.M.; Smith, H.A.; Michelotti, G.; Kechris, K.M.; Dabelea, D. A prospective study of associations between in utero exposure to gestational diabetes mellitus and metabolomic profiles during late childhood and adolescence. Diabetologia 2020, 63, 296–312. [Google Scholar] [CrossRef]
- Shokry, E.; Marchioro, L.; Uhl, O.; Bermudez, M.G.; Garcia-Santos, J.A.; Segura, M.T.; Campoy, C.; Koletzko, B. Impact of maternal BMI and gestational diabetes mellitus on maternal and cord blood metabolome: Results from the PREOBE cohort study. Acta Diabetol. 2019, 56, 421–430. [Google Scholar] [CrossRef]
- Perng, W.; Rifas-Shiman, S.L.; McCulloch, S.; Chatzi, L.; Mantzoros, C.; Hivert, M.F.; Oken, E. Associations of cord blood metabolites with perinatal characteristics, newborn anthropometry, and cord blood hormones in project viva. Metabolism 2017, 76, 11–22. [Google Scholar] [CrossRef]
- Barbour, L.A.; McCurdy, C.E.; Hernandez, T.L.; Kirwan, J.P.; Catalano, P.M.; Friedman, J.E. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes Care 2007, 30, S112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oken, E.; Kleinman, K.P.; Rich-Edwards, J.; Gillman, M.W. A nearly continuous measure of birth weight for gestational age using a United States national reference. BMC Pediatrics 2003, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Available online: https://www.BMI-for-AgeandSex(5–19Years) (accessed on 5 February 2021).
- Reinehr, T.; Wolters, B.; Knop, C.; Lass, N.; Hellmuth, C.; Harder, U.; Peissner, W.; Wahl, S.; Grallert, H.; Adamski, J.; et al. Changes in the serum metabolite profile in obese children with weight loss. Eur. J. Nutr. 2015, 54, 173–181. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.J.; Fahien, L.A.; Brown, L.J.; Hasan, N.M.; Buss, J.D.; Kendrick, M.A. Perspective: Emerging evidence for signaling roles of mitochondrial anaplerotic products in insulin secretion. Am. J. Physiol.-Endocrinol. Metab. 2005, 288, E1–E15. [Google Scholar] [CrossRef] [Green Version]
- Brunengraber, H.; Roe, C.R. Anaplerotic molecules: Current and future. J. Inherit. Metab. Dis. 2006, 29, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Gladyshev, V.N. The biological significance of methionine sulfoxide stereochemistry. Free Radic. Biol. Med. 2011, 50, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Styskal, J.; Nwagwu, F.A.; Watkins, Y.N.; Liang, H.; Richardson, A.; Musi, N.; Salmon, A.B. Methionine sulfoxide reductase A affects insulin resistance by protecting insulin receptor function. Free Radic. Biol. Med. 2013, 56, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Boini, K.M.; Xia, M.; Koka, S.; Gehr, T.W.B.; Li, P.-L. Sphingolipids in obesity and related complications. Front. Biosci. 2017, 22, 96–116. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Zhang, C.; Kilicarslan, M.; Piening, B.D.; Bjornson, E.; Hallström, B.M.; Groen, A.K.; Ferrannini, E.; Laakso, M.; Snyder, M.; et al. Integrated network analysis reveals an association between plasma mannose levels and insulin resistance. Cell Metab. 2016, 24, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Samad, F.; Hester, K.D.; Yang, G.; Hannun, Y.A.; Bielawski, J. Altered adipose and plasma sphingolipid metabolism in obesity. Diabetes 2006, 55, 2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slotte, J.P. Biological functions of sphingomyelins. Prog. Lipid Res. 2013, 52, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.P.; Reichetzeder, C.; Prehn, C.; von Websky, K.; Slowinski, T.; Chen, Y.P.; Yin, L.H.; Kleuser, B.; Yang, X.S.; Adamski, J.; et al. Fetal serum metabolites are independently associated with gestational diabetes mellitus. Cell Physiol. Biochem. 2018, 45, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Lian, K.; Shentu, X.; Fang, J.; Shao, J.; Chen, M.; Wang, Y.; Zhou, M.; Sun, H. BCAA catabolic defect alters glucose metabolism in lean mice. Front. Physiol. 2019, 10, 1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiuolo, J.; Oppedisano, F.; Gratteri, S.; Muscoli, C.; Mollace, V. Regulation of uric acid metabolism and excretion. Int. J. Cardiol. 2016, 213, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurtz, P.; Soininen, P.; Kangas, A.J.; Ronnemaa, T.; Lehtimaki, T.; Kahonen, M.; Viikari, J.S.; Raitakari, O.T.; Ala-Korpela, M. Branched-chain and aromatic amino acids are predictors of insulin resistance in young adults. Diabetes Care 2013, 36, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Rhee, E.P.; Larson, M.G.; Lewis, G.D.; McCabe, E.L.; Shen, D.; Palma, M.J.; Roberts, L.D.; Dejam, A.; Souza, A.L.; et al. Metabolite profiling identifies pathways associated with metabolic risk in humans. Circulation 2012, 125, 2222–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.S.; Lin, M.; Imamura, F.; Cai, W.; Wang, L.; Feng, J.P.; Ruan, Y.; Tang, J.; Wang, F.; Yang, H.; et al. Serum metabolomics profiles in response to n-3 fatty acids in Chinese patients with type 2 diabetes: A double-blind randomised controlled trial. Sci. Rep. 2016, 6, 29522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, N.D.; Magis, A.T.; Earls, J.C.; Glusman, G.; Levy, R.; Lausted, C.; McDonald, D.T.; Kusebauch, U.; Moss, C.L.; Zhou, Y.; et al. A wellness study of 108 individuals using personal, dense, dynamic data clouds. Nat. Biotechnol. 2017, 35, 747–756. [Google Scholar] [CrossRef]
- Prentice, K.J.; Luu, L.; Allister, E.M.; Liu, Y.; Jun, L.S.; Sloop, K.W.; Hardy, A.B.; Wei, L.; Jia, W.; Fantus, I.G.; et al. The furan fatty acid metabolite CMPF is elevated in diabetes and induces beta cell dysfunction. Cell Metab. 2014, 19, 653–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankinen, M.A.; Hanhineva, K.; Kolehmainen, M.; Lehtonen, M.; Auriola, S.; Mykkänen, H.; Poutanen, K.; Schwab, U.; Uusitupa, M. CMPF Does not associate with impaired glucose metabolism in individuals with features of metabolic syndrome. PLoS ONE 2015, 10, e0124379. [Google Scholar] [CrossRef] [PubMed]
- Webber, L.S.; Srinivasan, S.R.; Wattigney, W.A.; Berenson, G.S. Tracking of serum lipids and lipoproteins from childhood to adulthood: The bogalusa heart study. Am. J. Epidemiol. 1991, 133, 884–899. [Google Scholar] [CrossRef]
- Rundle, A.G.; Factor-Litvak, P.; Suglia, S.F.; Susser, E.S.; Kezios, K.L.; Lovasi, G.S.; Cirillo, P.M.; Cohn, B.A.; Link, B.G. Tracking of obesity in childhood into adulthood: Effects on body mass index and fat mass index at age 50. Child Obes. 2020, 16, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.M.; Srinivasan, S.R.; Xu, J.-H.; Chen, W.; Kieltyka, L.; Berenson, G.S. Utility of Childhood glucose homeostasis variables in predicting adult diabetes and related cardiometabolic risk factors. Bogalusa Heart Study 2010, 33, 670–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M. Why young adults hold the key to assessing the obesity epidemic in children. Arch. Pediatrics Adolesc. Med. 2008, 162, 682–687. [Google Scholar] [CrossRef] [Green Version]
- Powe, C.E.; Allard, C.; Battista, M.-C.; Doyon, M.; Bouchard, L.; Ecker, J.L.; Perron, P.; Florez, J.C.; Thadhani, R.; Hivert, M.-F. Heterogeneous contribution of insulin sensitivity and secretion defects to gestational diabetes mellitus. Diabetes Care 2016, 39, 1052–1055. [Google Scholar] [CrossRef] [Green Version]
- Perng, W.; Hector, E.C.; Song, P.X.K.; Tellez Rojo, M.M.; Raskind, S.; Kachman, M.; Cantoral, A.; Burant, C.F.; Peterson, K.E. Metabolomic determinants of metabolic risk in mexican adolescents. Obesity 2017, 25, 1594–1602. [Google Scholar] [CrossRef]
- Perng, W.; Oken, E.; Roumeliotaki, T.; Sood, D.; Siskos, A.P.; Chalkiadaki, G.; Dermitzaki, E.; Vafeiadi, M.; Kyrtopoulos, S.; Kogevinas, M.; et al. Leptin, acylcarnitine metabolites and development of adiposity in the Rhea mother–child cohort in Crete, Greece. Obes. Sci. Pract. 2016, 2, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Perng, W.; Harte, R.; Ringham, B.M.; Baylin, A.; Bellatorre, A.; Scherzinger, A.; Goran, M.I.; Dabelea, D. A Prudent dietary pattern is inversely associated with liver fat content among multi-ethnic youth. Pediatric Obes. 2021, 16, e12758. [Google Scholar] [CrossRef] [PubMed]
- Batis, C.; Mendez, M.A.; Gordon-Larsen, P.; Sotres-Alvarez, D.; Adair, L.; Popkin, B. Using both principal component analysis and reduced rank regression to study dietary patterns and diabetes in Chinese adults. Public Health Nutr. 2016, 19, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Batis, C.; Mendez, M.A.; Sotres-Alvarez, D.; Gordon-Larsen, P.; Popkin, B. Dietary pattern trajectories during 15 years of follow-up and HbA1c, insulin resistance and diabetes prevalence among Chinese adults. J. Epidemiol. Community Health 2014, 68, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Reiss, P.T.; Xiao, L.; Zipunnikov, V.; Lindquist, M.A.; Crainiceanu, C.M. Two-way principal component analysis for matrix-variate data, with an application to functional magnetic resonance imaging data. Biostatistics 2017, 18, 214–229. [Google Scholar] [CrossRef] [PubMed]
- Crume, T.L.; Ogden, L.; West, N.A.; Vehik, K.S.; Scherzinger, A.; Daniels, S.; McDuffie, R.; Bischoff, K.; Hamman, R.F.; Norris, J.M.; et al. Association of exposure to diabetes in utero with adiposity and fat distribution in a multiethnic population of youth: The exploring perinatal outcomes among children (EPOCH) study. Diabetologia 2011, 54, 87–92. [Google Scholar] [CrossRef] [Green Version]
- National Diabetes Data Group. Classification and diagnosis of diabetes mellitus and other categories of glucose intolerance. National Diabetes Data Group. Diabetes A J. Am. Diabetes Association. 1979, 28, 1039–1057. [Google Scholar] [CrossRef]
- Perng, W.; Francis, E.C.; Smith, H.A.; Carey, J.; Wang, D.; Kechris, K.M.; Dabelea, D. Sex-specific metabolite biomarkers of NAFLD in youth: A prospective study in the EPOCH cohort. J. Clin. Endocrinol. Metab. 2020, 105, e3437–e3450. [Google Scholar] [CrossRef]
- De Onis, M.; Onyango, A.W.; Borghi, E.; Siyam, A.; Nishida, C.; Siekmann, J. Development of a WHO growth reference for school-aged children and adolescents. Bull. World Health Organ. 2007, 85, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Cullen, K.W.; Watson, K.; Zakeri, I. Relative reliability and validity of the block kids questionnaire among youth aged 10 to 17 Years. J. Am. Diet. Assoc. 2008, 108, 862–866. [Google Scholar] [CrossRef]
- Weston, A.T.; Petosa, R.; Pate, R.R. Validation of an instrument for measurement of physical activity in youth. Med. Sci. Sports Exerc. 1997, 29, 138–143. [Google Scholar] [CrossRef]
- Benjamini, Y.; Liu, W. A step-down multiple hypotheses testing procedure that controls the false discovery rate under independence. J. Stat. Plan. Inference 1999, 82, 163–170. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Overnutrition Typology | |||||
|---|---|---|---|---|---|
| No GDM or Obesity | Obese Only | GDM Only | GDM & Obesity | p1 | |
| Maternal Characteristics | n = 293 | n = 66 | n = 56 | n = 25 | |
| Pre-pregnancy BMI (kg/m2) | 23.2 ± 3.1 | 35.0 ± 5.3 | 24.0 ± 3.4 | 35.1 ± 4.1 | - |
| Education level | 0.10 | ||||
| <High school | 3.1 (9) | 6.1 (4) | 5.4 (3) | 4.0 (1) | |
| High school or some college | 44.4 (130) | 62.1 (41) | 46.4 (26) | 56.0 (14) | |
| Associates degree or higher | 52.6 (154) | 31.8 (21) | 48.2 (27) | 40.0 (10) | |
| Smoked during pregnancy | 7.2 (21) | 6.1 (4) | 17.9 (10) | 20.0 (5) | 0.01 |
| Offspring characteristics at birth | |||||
| Female | 48.8 (143) | 47.0 (31) | 46.4 (26) | 28.0 (7) | 0.26 |
| Race/ethnicity | <0.001 | ||||
| Non-Hispanic White | 42.3 (124) | 18.2 (12) | 71.4 (40) | 52.0 (13) | |
| Hispanic | 44.0 (129) | 57.6 (38) | 21.4 (12) | 44.0 (11) | |
| Non-Hispanic Black | 7.2 (21) | 21.2 (14) | 3.6 (2) | 4.0 (1) | |
| Non-Hispanic Other | 6.5 (19) | 3.0 (2) | 3.6 (2) | 0.0 (0) | |
| Birthweight for gestational age z-score 2 | −0.4 ± 0.9 | −0.3 ± 1.0 | −0.1 ± 1.0 | 0.0 ± 0.9 | 0.02 |
| Childhood visit | |||||
| Age, years | 10.7 ± 1.4 | 10.5 ± 1.4 | 9.5 ± 1.8 | 9.8 ± 1.5 | <0.001 |
| BMI (kg/m2) | 18.6 ± 4.0 | 21.1 ± 5.5 | 18.1 ± 4.2 | 20.6 ± 5.7 | <0.001 |
| BMI z-score | 0.2 ± 1.2 | 0.9 ± 1.1 | 0.1 ± 1.4 | 0.8 ± 1.1 | <0.001 |
| Kilocalories | 1791.5 ± 565.3 | 1819.8 ± 581.1 | 1773.0 ± 495.5 | 1727.4 ± 415.2 | 0.90 |
| Energy expenditure | 68.4 ±11.3 | 65.15 ± 9.41 | 66.04 ± 9.39 | 65.64 ± 11.91 | 0.09 |
| Adolescent visit | |||||
| Age, years | 16.7 ± 1.1 | 16.4 ± 1.3 | 15.8 ± 1.1 | 16.0 ± 1.0 | <0.001 |
| BMI (kg/m2) | 22.7 ± 4.8 | 28.0 ± 7.1 | 22.6 ± 4.6 | 24.8 ± 6.3 | <0.001 |
| BMI z-score 3 | 0.2 ± 1.1 | 1.2 ± 1.0 | 0.4 ± 1.1 | 0.8 ± 1.1 | <0.001 |
| Kilocalories | 1672.5 ± 717.0 | 1599.4 ± 762.3 | 1730.2 ± 887.6 | 1660.7 ± 577.5 | 0.88 |
| Energy expenditure | 70.5 ± 16.1 | 66.87 ± 11.89 | 67.8 ± 13.5 | 78.25 ± 20.12 | 0.05 |
| Compound | Superclass | Subclass | p-Value | FDR p-Value |
|---|---|---|---|---|
| Tyrosine | Amino Acid | Tyrosine Metabolism | 0.001 | 0.119 |
| Homoarginine | Amino Acid | Urea cycle; Arginine and Proline Metabolism | 0.002 | 0.144 |
| 2-hydroxy-3-methylvalerate | Amino Acid | Leucine, Isoleucine and Valine Metabolism | 0.004 | 0.144 |
| 3-methyl-2-oxobutyrate | Amino Acid | Leucine, Isoleucine and Valine Metabolism | 0.006 | 0.144 |
| 2-aminoadipate | Amino Acid | Lysine Metabolism | 0.007 | 0.154 |
| Glycine | Amino Acid | Glycine, Serine and Threonine Metabolism | 0.007 | 0.154 |
| N-acetylglycine | Amino Acid | Glycine, Serine and Threonine Metabolism | 0.008 | 0.158 |
| Methionine sulfoxide | Amino Acid | Methionine, Cysteine, SAM, Taurine | 0.010 | 0.167 |
| Alpha-hydroxyisocaproate | Amino Acid | Leucine, Isoleucine and Valine Metabolism | 0.014 | 0.190 |
| Mannitol/sorbitol | Carbohydrate | Fructose, Mannose and Galactose Metabolism | 0.003 | 0.144 |
| Glucuronate | Carbohydrate | Aminosugar Metabolism | 0.004 | 0.144 |
| Mannose | Carbohydrate | Fructose, Mannose and Galactose Metabolism | 0.005 | 0.144 |
| Pantothenate | Cofactors, Vitamins | Pantothenate and CoA Metabolism | 0.001 | 0.119 |
| Alpha-ketoglutarate | Energy | TCA Cycle | 0.001 | 0.119 |
| Citrate | Energy | TCA Cycle | 0.004 | 0.144 |
| Malate | Energy | TCA Cycle | 0.006 | 0.144 |
| Succinate | Energy | TCA Cycle | 0.012 | 0.181 |
| 12-HETE | Lipid | Eicosanoid | 0.000 | 0.119 |
| 13-HODE + 9-HODE | Lipid | Fatty Acid, Monohydroxy | 0.002 | 0.119 |
| Hydroxy-CMPF 1 | Lipid | Fatty Acid, Dicarboxylate | 0.005 | 0.144 |
| Choline | Lipid | Phospholipid Metabolism | 0.005 | 0.144 |
| 3-hydroxybutyroylglycine 1 | Lipid | Fatty Acid Metabolism(Acyl Glycine) | 0.005 | 0.144 |
| 7-alpha-hydroxy-3-oxo-4-cholestenoate (7-Hoca) | Lipid | Sterol | 0.006 | 0.144 |
| Palmitoyl-arachidonoyl-glycerol (16:0/20:4) [2] 1 | Lipid | Diacylglycerol | 0.006 | 0.144 |
| N-oleoylserine | Lipid | Endocannabinoid | 0.007 | 0.154 |
| 1-linoleoyl-GPA (18:2) 1 | Lipid | Lysophospholipid | 0.009 | 0.158 |
| Hexanoylcarnitine (C6) | Lipid | Fatty Acid Metabolism(Acyl Carnitine) | 0.009 | 0.161 |
| 3-carboxy-4-methyl-5-propyl-2-furanpropanoate (CMPF) | Lipid | Fatty Acid, Dicarboxylate | 0.012 | 0.180 |
| Glycosyl-N-palmitoyl-sphingosine (d18:1/16:0) | Lipid | Hexosylceramides (HCER) | 0.012 | 0.181 |
| 1-(1-enyl-palmitoyl)-GPC (P-16:0) 1 | Lipid | Lysoplasmalogen | 0.013 | 0.188 |
| Sphingomyelin (d18:2/14:0, d18:1/14:1) 1 | Lipid | Sphingomyelins | 0.014 | 0.190 |
| Dodecadienoate (12:2) 1 | Lipid | Fatty Acid, Dicarboxylate | 0.016 | 0.193 |
| 1-(1-enyl-palmitoyl)-2-oleoyl-GPE (P-16:0/18:1) 1 | Lipid | Plasmalogen | 0.016 | 0.193 |
| Sphingomyelin (d18:0/18:0, d19:0/17:0) 1 | Lipid | Dihydrosphingomyelins | 0.016 | 0.193 |
| Dihydroorotate | Nucleotide | Pyrimidine Metabolism, Orotate contain. | 0.002 | 0.119 |
| Urate | Nucleotide | Purine Metabolism, (Hypo)Xanthine/Inosine | 0.009 | 0.158 |
| N1-methyladenosine | Nucleotide | Purine Metabolism, Adenine contain. | 0.010 | 0.167 |
| Guanosine | Nucleotide | Purine Metabolism, Guanine contain. | 0.013 | 0.189 |
| Fibrinopeptide A, des-ala(1) 1 | Peptide | Fibrinogen Cleavage Peptide | 0.001 | 0.119 |
| Gamma-glutamylglutamate | Peptide | Gamma-glutamyl Amino Acid | 0.004 | 0.144 |
| Gamma-glutamylcitrulline 1 | Peptide | Gamma-glutamyl Amino Acid | 0.004 | 0.144 |
| Gamma-glutamyl-alpha-lysine | Peptide | Gamma-glutamyl Amino Acid | 0.005 | 0.144 |
| Glycylvaline | Peptide | Dipeptide | 0.008 | 0.158 |
| Gamma-glutamylthreonine | Peptide | Gamma-glutamyl Amino Acid | 0.009 | 0.158 |
| Gamma-glutamyl-2-aminobutyrate | Peptide | Gamma-glutamyl Amino Acid | 0.009 | 0.163 |
| Gamma-glutamylglycine | Peptide | Gamma-glutamyl Amino Acid | 0.014 | 0.190 |
| Phenylalanylglycine | Peptide | Dipeptide | 0.015 | 0.193 |
| Sulfate of piperine metabolite C16H19NO3 (2) 1 | Xenobiotics | Food Component/Plant | 0.000 | 0.119 |
| Sulfate of piperine metabolite C16H19NO3 (3) 1 | Xenobiotics | Food Component/Plant | 0.001 | 0.119 |
| Quinate | Xenobiotics | Food Component/Plant | 0.004 | 0.144 |
| Piperine | Xenobiotics | Food Component/Plant | 0.004 | 0.144 |
| Perfluorooctanesulfonate (PFOS) | Xenobiotics | Chemical | 0.011 | 0.174 |
| Factor Loading | ||||
|---|---|---|---|---|
| Childhood Visit | Adolescent Visit | Compound | Superclass | Subclass |
| Factor label: γ-glutamyl | ||||
| Factor 1 (44% variance) | Factor 1 (20% variance) | |||
| 0.81 | 0.83 | Gamma-glutamylglutamate | Peptide | Gamma-glutamyl Amino Acid |
| 0.77 | 0.81 | Gamma-glutamyl-alpha-lysine | Peptide | Gamma-glutamyl Amino Acid |
| 0.72 | 0.69 | Gamma-glutamylglycine | Peptide | Gamma-glutamyl Amino Acid |
| 0.70 | 0.74 | Methionine sulfoxide | Amino Acid | Methionine, Cysteine, SAM, Taurine Metabolism |
| 0.66 | 0.68 | Glycylvaline | Peptide | Dipeptide |
| 0.66 | 0.68 | 1-linoleoyl-GPA (18:2) | Lipid | Lysophospholipid |
| 0.61 | <0.40 | 13-HODE + 9-HODE | Lipid | Fatty Acid, Monohydroxy |
| 0.61 | 0.65 | Choline | Lipid | Phospholipid Metabolism |
| <0.40 | 0.62 | Gamma-glutamylthreonine | Peptide | Gamma-glutamyl Amino Acid |
| Factor label: Sphingomyelin-mannose | ||||
| Factor 3 a (8% variance) | Factor 2 a (13% variance) | |||
| 0.61 | 0.58 | Sphingomyelin (d18:2/14:0, d18:1/14:1) | Lipid | Sphingomyelins |
| 0.59 | 0.64 | Sphingomyelin (d18:0/18:0, d19:0/17:0) | Lipid | Dihydrosphingomyelins |
| 0.54 | 0.59 | Mannose | Carbohydrate | Fructose, Mannose and Galactose Metabolism |
| 0.52 | 0.58 | Homoarginine | Amino Acid | Urea cycle; Arginine and Proline Metabolism |
| 0.45 | 0.50 | N1-methyladenosine | Nucleotide | Purine Metabolism, Adenine containing |
| Factor label: Skeletal muscle metabolism | ||||
| Factor 4 (6% variance) | Factor 4 (10% variance) | |||
| 0.63 | 0.76 | Alpha-hydroxyisocaproate | Amino Acid | Leucine, Isoleucine and Valine Metabolism |
| 0.49 | 0.62 | 2-hydroxy-3-methylvalerate | Amino Acid | Leucine, Isoleucine and Valine Metabolism |
| 0.40 | 0.51 | Malate | Energy | TCA Cycle |
| 0.40 | 0.51 | Urate | Nucleotide | Purine Metabolism, (Hypo)Xanthine/Inosine |
| 0.40 | <0.40 | Citrate | Energy | TCA Cycle |
| <0.40 | 0.41 | 7-alpha-hydroxy-3-oxo-4-cholestenoate (7-Hoca) | Lipid | Sterol |
| Factor label: 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) | ||||
| Factor 6 (3% variance) | Factor 6 (4% variance) | |||
| 0.74 | 0.91 | 3-carboxy-4-methyl-5-propyl-2-furanpropanoate (CMPF) | Lipid | Fatty Acid, Dicarboxylate |
| 0.71 | 0.90 | Hydroxy-CMPF | Lipid | Fatty Acid, Dicarboxylate |
| Factor 2 a (11% variance) | Factor 3 a (13% variance) | |||
| 0.58 | 0.94 | Sulfate of piperine metabolite C16H19NO3 (2) | Xenobiotics | Food Component/Plant |
| 0.56 | 0.93 | Sulfate of piperine metabolite C16H19NO3 (3) | Xenobiotics | Food Component/Plant |
| 0.51 | 0.85 | Piperine | Xenobiotics | Food Component/Plant |
| 0.50 | <0.40 | 2-aminoadipate | Amino Acid | Lysine Metabolism |
| 0.45 | <0.40 | 2-hydroxy-3-methylvalerate | Amino Acid | Leucine, Isoleucine and Valine Metabolism |
| Factor 5 (4% variance) | Factor 5 (4% variance) | |||
| 0.50 | <0.40 | Sulfate of piperine metabolite C16H19NO3 (2) | Xenobiotics | Food Component/Plant |
| 0.49 | <0.40 | Sulfate of piperine metabolite C16H19NO3 (3) | Xenobiotics | Food Component/Plant |
| 0.43 | <0.40 | Piperine | Xenobiotics | Food Component/Plant |
| <0.40 | 0.53 | Dodecadienoate (12:2) * | Lipid | Fatty Acid, Dicarboxylate |
| <0.40 | 0.53 | 3-hydroxybutyroylglycine * | Lipid | Fatty Acid Metabolism (Acyl Glycine) |
| <0.40 | 0.44 | Hexanoylcarnitine (C6) | Lipid | Fatty Acid Metabolism (Acyl Carnitine) |
| <0.40 | 0.57 | N-acetylglycine | Amino Acid | Glycine, Serine and Threonine Metabolism |
| <0.40 | 0.45 | Glycine | Amino Acid | Glycine, Serine and Threonine Metabolism |
| OB + GDM vs. GDM Only | OB + GDM vs. OB Only | OB Only vs. GDM Only | |
|---|---|---|---|
| Factor | Adjusted | Adjusted | Adjusted |
| γ-glutamyl | −0.20 (−0.50, 0.10) | −0.14 (−0.44, 0.17) | −0.06 (−0.32, 0.20) |
| Sphingomyelin-mannose | 0.29 (−0.04, 0.63) | −0.03 (−0.38, 0.33) | 0.32 (0.07, 0.57) * |
| Skeletal muscle metabolism | 0.36 (0.09, 0.64) * | 0.47 (0.21, 0.72) * | −0.10 (−0.34, 0.13) |
| CMPF | 0.50 (0.11, 0.89) * | 0.05 (−0.34, 0.44) | 0.45 (0.17, 0.73) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francis, E.C.; Kechris, K.; Cohen, C.C.; Michelotti, G.; Dabelea, D.; Perng, W. Metabolomic Profiles in Childhood and Adolescence Are Associated with Fetal Overnutrition. Metabolites 2022, 12, 265. https://doi.org/10.3390/metabo12030265
Francis EC, Kechris K, Cohen CC, Michelotti G, Dabelea D, Perng W. Metabolomic Profiles in Childhood and Adolescence Are Associated with Fetal Overnutrition. Metabolites. 2022; 12(3):265. https://doi.org/10.3390/metabo12030265
Chicago/Turabian StyleFrancis, Ellen C., Katerina Kechris, Catherine C. Cohen, Gregory Michelotti, Dana Dabelea, and Wei Perng. 2022. "Metabolomic Profiles in Childhood and Adolescence Are Associated with Fetal Overnutrition" Metabolites 12, no. 3: 265. https://doi.org/10.3390/metabo12030265
APA StyleFrancis, E. C., Kechris, K., Cohen, C. C., Michelotti, G., Dabelea, D., & Perng, W. (2022). Metabolomic Profiles in Childhood and Adolescence Are Associated with Fetal Overnutrition. Metabolites, 12(3), 265. https://doi.org/10.3390/metabo12030265