
Effects of Different Prenatal Nutrition Strategies on the Liver Metabolome of Bulls and Its Correlation with Body and Liver Weight

 ,
, 
Abstract
:
1. Introduction
2. Results
2.1. Pre-Slaughter Body Weight and Liver Weight
2.2. Univariate Analysis of Liver Metabolome
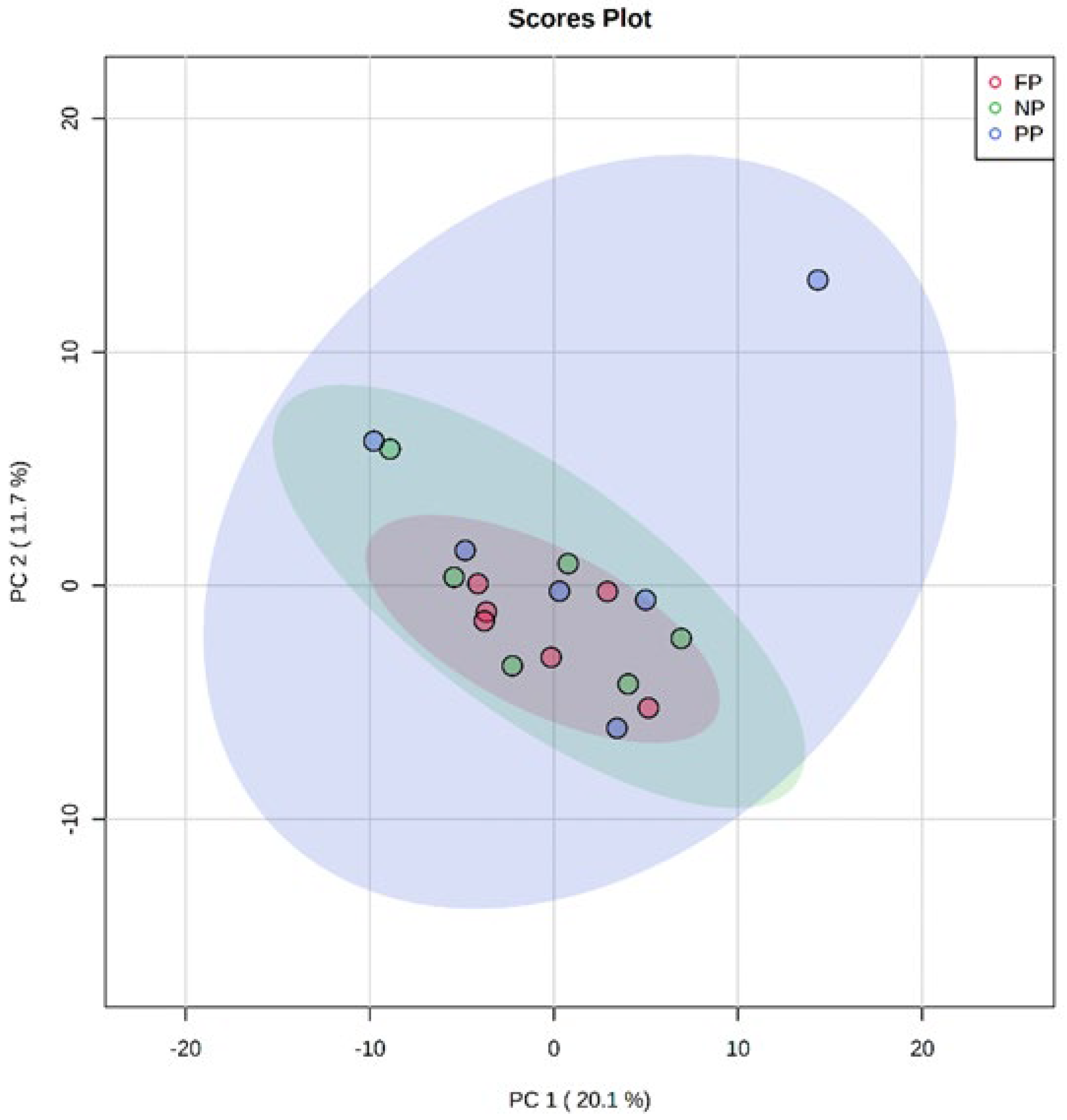
2.3. Principal Component Analysis (PCA)
2.4. Pearson’s Correlation Analysis
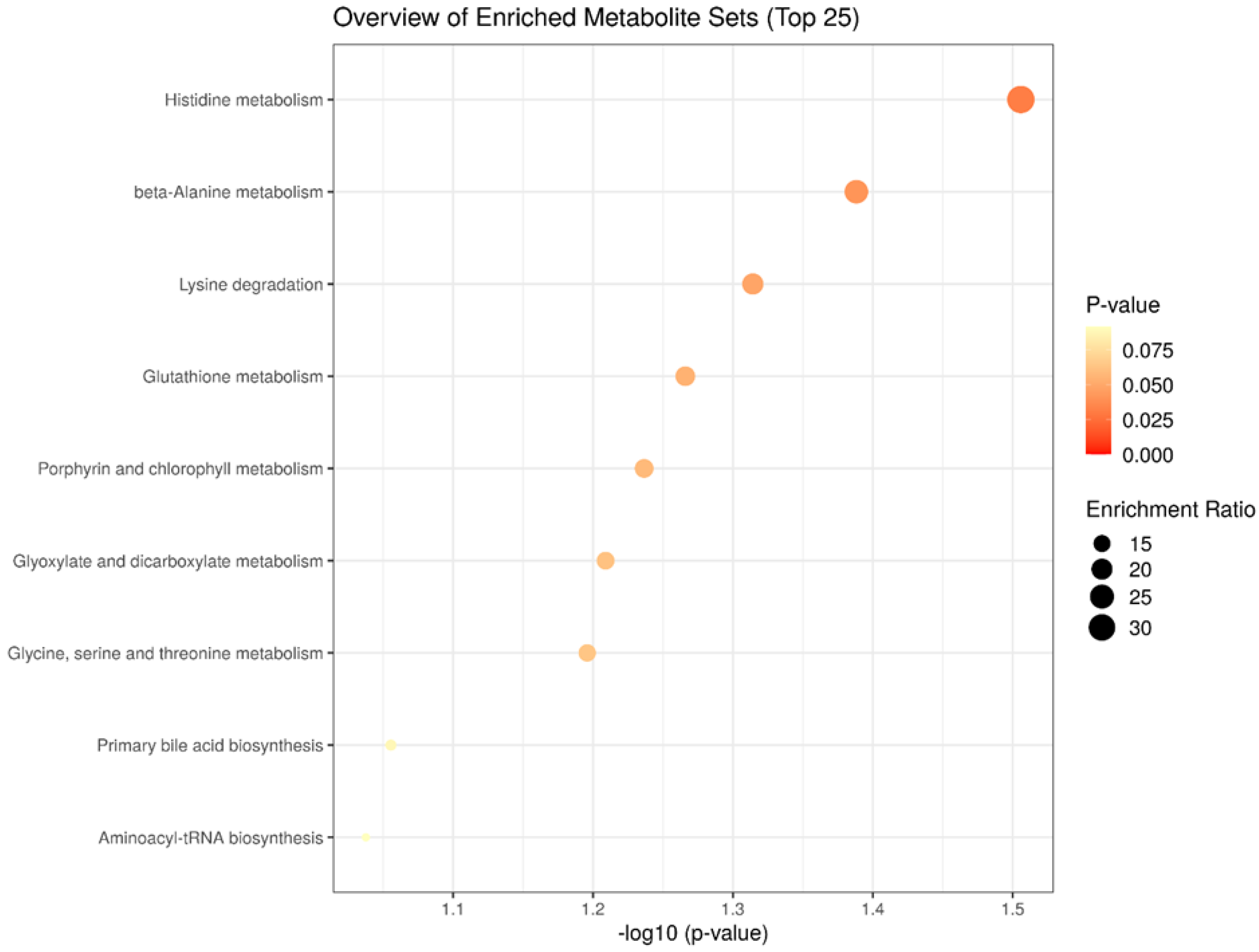
2.5. Enrichment Analysis
3. Discussion
4. Materials and Methods
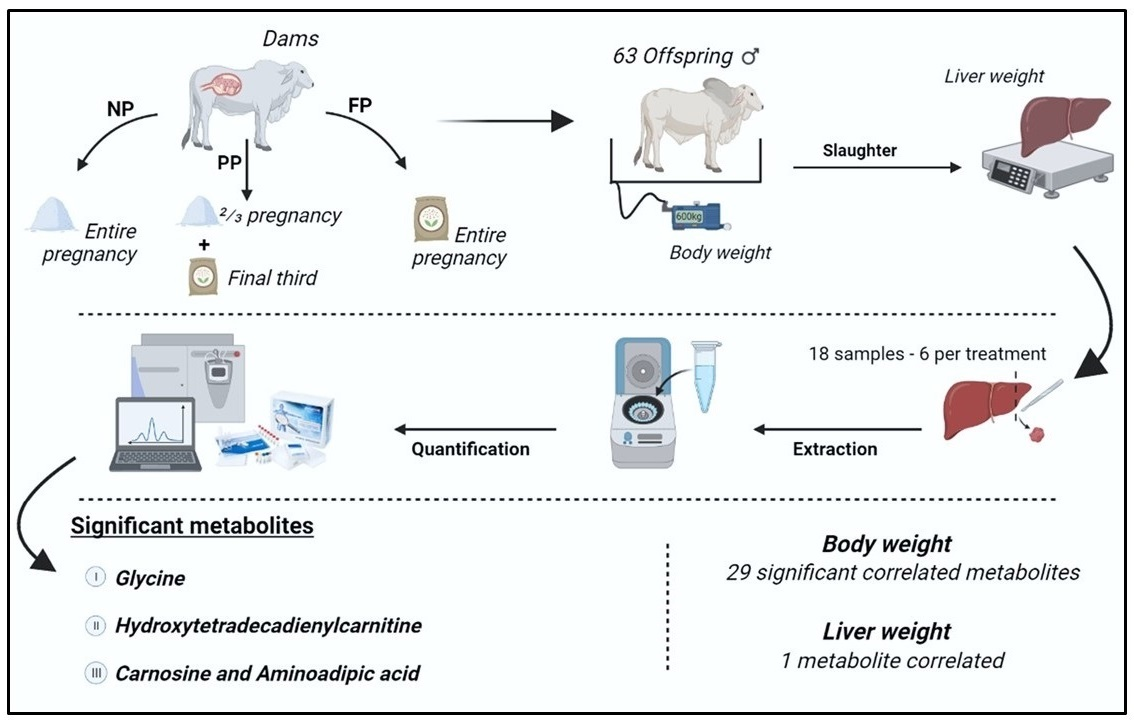
4.1. Experimental Design
4.2. Phenotypes, Liver Sample Collection and Preparation
4.3. Targeted Metabolomics
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zago, D.; Canozzi, M.E.A.; Barcellos, J.O.J. Pregnant Cow Nutrition and Its Effects on Foetal Weight—A Meta-Analysis. J. Agric. Sci. 2019, 157, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Long, N.M.; Prado-Cooper, M.J.; Krehbiel, C.R.; Desilva, U.; Wettemann, R.P. Effects of Nutrient Restriction of Bovine Dams during Early Gestation on Postnatal Growth, Carcass and Organ Characteristics, and Gene Expression in Adipose Tissue and Muscle. J. Anim. Sci. 2010, 88, 3251–3261. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Zhao, J.X.; Yan, X.; Huang, Y.; Nicodemus, L.V.; Yue, W.; Mccormick, R.J.; Zhu, M.J. Fetal Muscle Development, Mesenchymal Multipotent Cell Differentiation, and Associated Signaling Pathways. J. Anim. Sci. 2011, 89, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Franco, P.N.; Durrant, L.M.; Doan, C.; Carreon, D.; Beltran, A.; Jullienne, A.; Obenaus, A.; Pearce, W.J. Maternal Undernutrition Modulates Neonatal Rat Cerebrovascular Structure, Function, and Vulnerability to Mild Hypoxic-Ischemic Injury via Corticosteroid-Dependent and -Independent Mechanisms. Int. J. Mol. Sci. 2021, 22, 680. [Google Scholar] [CrossRef]
- Ravelli, A.C.J.; Van Der Meulen, J.H.P.; Michels, R.P.J.; Osmond, C.; Barker, D.J.P.; Hales, C.N.; Bleker, O.P. Glucose Tolerance in Adults after Prenatal Exposure to Famine. Lancet 1998, 351, 173–177. [Google Scholar] [CrossRef]
- Vonnahme, K.A.; Lemley, C.O.; Caton, J.S.; Meyer, A.M. Impacts of Maternal Nutrition on Vascularity of Nutrient Transferring Tissues during Gestation and Lactation. Nutrients 2015, 7, 3497–3523. [Google Scholar] [CrossRef]
- Caton, J.S.; Crouse, M.S.; Reynolds, L.P.; Neville, T.L.; Dahlen, C.R.; Ward, A.K.; Swanson, K.C. Maternal Nutrition and Programming of Offspring Energy Requirements. Transl. Anim. Sci. 2019, 3, 976–990. [Google Scholar] [CrossRef] [Green Version]
- Grazul-Bilska, A.T.; Borowicz, P.P.; Johnson, M.L.; Minten, M.A.; Bilski, J.J.; Wroblewski, R.; Redmer, D.A.; Reynolds, L.P. Placental Development during Early Pregnancy in Sheep: Vascular Growth and Expression of Angiogenic Factors in Maternal Placenta. Reproduction 2010, 140, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Sandovici, I.; Hoelle, K.; Angiolini, E.; Constância, M. Placental Adaptations to the Maternal–Fetal Environment: Implications for Fetal Growth and Developmental Programming. Reprod. Biomed. Online 2012, 25, 68–89. [Google Scholar] [CrossRef] [Green Version]
- Diniz, W.J.S.; Crouse, M.S.; Cushman, R.A.; McLean, K.J.; Caton, J.S.; Dahlen, C.R.; Reynolds, L.P.; Ward, A.K. Cerebrum, Liver, and Muscle Regulatory Networks Uncover Maternal Nutrition Effects in Developmental Programming of Beef Cattle during Early Pregnancy. Sci. Rep. 2021, 11, 2771. [Google Scholar] [CrossRef]
- Foroutan, A.; Devos, J.; Wishart, D.S.; Li, C.; Colazo, M.; Kastelic, J.; Thundathil, J.; Fitzsimmons, C. Impact of Prenatal Maternal Nutrition and Parental Residual Feed Intake (RFI) on MRNA Abundance of Metabolic Drivers of Growth and Development in Young Angus Bulls. Livest. Sci. 2021, 243, 104365. [Google Scholar] [CrossRef]
- Stalker, M.J. Pathologic Basis of Veterinary Disease, 4th ed.; McGavin, M.D., Zachary, J.F., Eds.; Canadian Veterinary Medical Association: St. Louis, MO, USA, 2007; Volume 48, ISBN 0-3230-2870-5. [Google Scholar]
- Cônsolo, N.R.B.; Buarque, V.L.M.; Silva, J.; Poleti, M.D.; Barbosa, L.C.G.S.; Higuera-Padilla, A.; Gómez, J.F.M.; Colnago, L.A.; Gerrard, D.E.; Saran Netto, A.; et al. Muscle and Liver Metabolomic Signatures Associated with Residual Feed Intake in Nellore Cattle. Anim. Feed Sci. Technol. 2021, 271, 114757. [Google Scholar] [CrossRef]
- Fonseca, L.D.; Eler, J.P.; Pereira, M.A.; Rosa, A.F.; Alexandre, P.A.; Moncau, C.T.; Salvato, F.; Rosa-Fernandes, L.; Palmisano, G.; Ferraz, J.B.S.; et al. Liver Proteomics Unravel the Metabolic Pathways Related to Feed Efficiency in Beef Cattle. Sci. Rep. 2019, 9, 5364. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, P.A.; Kogelman, L.J.A.; Santana, M.H.A.; Passarelli, D.; Pulz, L.H.; Fantinato-Neto, P.; Silva, P.L.; Leme, P.R.; Strefezzi, R.F.; Coutinho, L.L.; et al. Liver Transcriptomic Networks Reveal Main Biological Processes Associated with Feed Efficiency in Beef Cattle. BMC Genom. 2015, 16, 1073. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S. Metabolomics for Investigating Physiological and Pathophysiological Processes. Physiol. Rev. 2019, 99, 1819–1875. [Google Scholar] [CrossRef]
- Pintus, R.; Dessì, A.; Mussap, M.; Fanos, V. Metabolomics Can Provide New Insights into Perinatal Nutrition. Acta Paediatr. 2021, 1–9. [Google Scholar] [CrossRef]
- Paul, H.A.; Bomhof, M.R.; Vogel, H.J.; Reimer, R.A. Diet-Induced Changes in Maternal Gut Microbiota and Metabolomic Profiles Influence Programming of Offspring Obesity Risk in Rats. Sci. Rep. 2016, 6, 20683. [Google Scholar] [CrossRef]
- Ross, A.B.; Barman, M.; Hartvigsson, O.; Lundell, A.C.; Savolainen, O.; Hesselmar, B.; Wold, A.E.; Sandberg, A.S. Umbilical Cord Blood Metabolome Differs in Relation to Delivery Mode, Birth Order and Sex, Maternal Diet and Possibly Future Allergy Development in Rural Children. PLoS ONE 2021, 16, e0242978. [Google Scholar] [CrossRef]
- Elolimy, A.; Alharthi, A.; Zeineldin, M.; Parys, C.; Helmbrecht, A.; Loor, J.J. Supply of Methionine During Late-Pregnancy Alters Fecal Microbiota and Metabolome in Neonatal Dairy Calves Without Changes in Daily Feed Intake. Front. Microbiol. 2019, 10, 2159. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ford, S.P.; Means, W.J.; Hess, B.W.; Nathanielsz, P.W.; Du, M. Maternal Nutrient Restriction Affects Properties of Skeletal Muscle in Offspring. J. Physiol. 2006, 575, 241–250. [Google Scholar] [CrossRef]
- Long, N.M.; Vonnahme, K.A.; Hess, B.W.; Nathanielsz, P.W.; Ford, S.P. Effects of Early Gestational Undernutrition on Fetal Growth, Organ Development, and Placentomal Composition in the Bovine. J. Anim. Sci. 2009, 87, 1950–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Otomaru, K.; Oshima, K.; Goto, Y.; Oshima, I.; Muroya, S.; Sano, M.; Saneshima, R.; Nagao, Y.; Kinoshita, A.; et al. Effects of Low and High Levels of Maternal Nutrition Consumed for the Entirety of Gestation on the Development of Muscle, Adipose Tissue, Bone, and the Organs of Wagyu Cattle Fetuses. Anim. Sci. J. 2021, 92, e13600. [Google Scholar] [CrossRef] [PubMed]
- Paradis, F.; Wood, K.M.; Swanson, K.C.; Miller, S.P.; McBride, B.W.; Fitzsimmons, C. Maternal Nutrient Restriction in Mid-to-Late Gestation Influences Fetal MRNA Expression in Muscle Tissues in Beef Cattle. BMC Genom. 2017, 18, 632. [Google Scholar] [CrossRef] [PubMed]
- Clavé, A.; Ripoll, G.; Casasús, I.; Sanz, A. Long-Term Effects of Early Maternal Undernutrition on the Growth, Physiological Profiles, Carcass and Meat Quality of Male Beef Offspring. Res. Vet. Sci. 2022, 142, 1–11. [Google Scholar] [CrossRef]
- Maresca, S.; Valiente, S.L.; Rodriguez, A.M.; Testa, L.M.; Long, N.M.; Quintans, G.I.; Pavan, E. The Influence of Protein Restriction during Mid- to Late Gestation on Beef Offspring Growth, Carcass Characteristic and Meat Quality. Meat Sci. 2019, 153, 103–108. [Google Scholar] [CrossRef]
- Hou, Y.; Yao, K.; Yin, Y.; Wu, G. Endogenous Synthesis of Amino Acids Limits Growth, Lactation, and Reproduction in Animals. Adv. Nutr. 2016, 7, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, X.; Hou, Y.; Yin, Y.; Qiu, Y.; Wu, G.; Hu, C.A.A. Roles of Amino Acids in Preventing and Treating Intestinal Diseases: Recent Studies with Pig Models. Amino Acids 2017, 49, 1277–1291. [Google Scholar] [CrossRef]
- Li, P.; Wu, G. Roles of Dietary Glycine, Proline, and Hydroxyproline in Collagen Synthesis and Animal Growth. Amino Acids 2018, 50, 29–38. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, J.; Wu, G. Glycine Metabolism in Animals and Humans: Implications for Nutrition and Health. Amino Acids 2013, 45, 463–477. [Google Scholar] [CrossRef]
- Ghaffari, M.H.; Sadri, H.; Schuh, K.; Dusel, G.; Prehn, C.; Adamski, J.; Koch, C.; Sauerwein, H. Alterations of the Acylcarnitine Profiles in Blood Serum and in Muscle from Periparturient Cows with Normal or Elevated Body Condition. J. Dairy Sci. 2020, 103, 4777–4794. [Google Scholar] [CrossRef]
- Makrecka-Kuka, M.; Sevostjanovs, E.; Vilks, K.; Volska, K.; Antone, U.; Kuka, J.; Makarova, E.; Pugovics, O.; Dambrova, M.; Liepinsh, E. Plasma Acylcarnitine Concentrations Reflect the Acylcarnitine Profile in Cardiac Tissues. Sci. Rep. 2017, 7, 17528. [Google Scholar] [CrossRef] [Green Version]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or Inflicting Insulin Resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, J.W. Review: Lipid Biology in the Periparturient Dairy Cow: Contemporary Perspectives. Animal 2020, 14, s165–s175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Jang, H.B.; Kim, W.H.; Park, K.J.; Kim, K.Y.; Park, S.I.; Lee, H.J. 2-Aminoadipic Acid (2-AAA) as a Potential Biomarker for Insulin Resistance in Childhood Obesity. Sci. Rep. 2019, 9, 13610. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, J.; Theves, M.; Strauch, C.; Nemet, I.; Liu, X.; Qian, J.; Giblin, F.J.; Monnier, V.M. Mechanism of Lysine Oxidation in Human Lens Crystallins during Aging and in Diabetes. J. Biol. Chem. 2009, 284, 34618–34627. [Google Scholar] [CrossRef] [Green Version]
- Estaras, M.; Ameur, F.Z.; Estévez, M.; Díaz-Velasco, S.; Gonzalez, A. The Lysine Derivative Aminoadipic Acid, a Biomarker of Protein Oxidation and Diabetes-Risk, Induces Production of Reactive Oxygen Species and Impairs Trypsin Secretion in Mouse Pancreatic Acinar Cells. Food Chem. Toxicol. 2020, 145, 111594. [Google Scholar] [CrossRef]
- Choi, C.B.; Kwon, H.; Kim, S., II; Yang, U.M.; Lee, J.H.; Park, E.K. Effects of Rice Bran, Flax Seed, and Sunflower Seed on Growth Performance, Carcass Characteristics, Fatty Acid Composition, Free Amino Acid and Peptide Contents, and Sensory Evaluations of Native Korean Cattle (Hanwoo). Asian-Australas. J. Anim. Sci. 2016, 29, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Tucker, H.A.; Hanigan, M.D.; Escobar, J.; Doane, P.H.; Donkin, S.S. Hepatic Expression of Aminoadipate Semialdehyde Synthase Is Unchanged by Postruminal Lysine Supply in Lactating Dairy Cows. J. Dairy Sci. 2017, 100, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, H.Y.; Wang, Y.M.; Yang, Z.Q.; Liu, J.X.; Wu, Y.M.; Yan, T.; Ye, H.W. Effects of Dietary Supplementation of Methionine and Lysine on Milk Production and Nitrogen Utilization in Dairy Cows. J. Dairy Sci. 2010, 93, 3661–3670. [Google Scholar] [CrossRef]
- Wu, G. Amino Acids: Biochemistry and Nutrition; CRC Press: Boca Raton, FL, USA, 2013; ISBN 9781439861905. [Google Scholar]
- Hipkiss, A.R.; Gaunitz, F. Inhibition of Tumour Cell Growth by Carnosine: Some Possible Mechanisms. Amino Acids 2014, 46, 327–337. [Google Scholar] [CrossRef]
- Barca, A.; Ippati, S.; Urso, E.; Vetrugno, C.; Storelli, C.; Maffia, M.; Romano, A.; Verri, T. Carnosine Modulates the Sp1-Slc31a1/Ctr1 Copper-Sensing System and Influences Copper Homeostasis in Murine CNS-Derived Cells. Am. J. Physiol. Cell Physiol. 2019, 316, C235–C245. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and Pathophysiology of Carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Oppermann, H.; Alvanos, A.; Seidel, C.; Meixensberger, J.; Gaunitz, F. Carnosine Influences Transcription via Epigenetic Regulation as Demonstrated by Enhanced Histone Acetylation of the Pyruvate Dehydrogenase Kinase 4 Promoter in Glioblastoma Cells. Amino Acids 2019, 51, 61–71. [Google Scholar] [CrossRef]
- Purchas, R.W.; Rutherfurd, S.M.; Pearce, P.D.; Vather, R.; Wilkinson, B.H.P. Concentrations in Beef and Lamb of Taurine, Carnosine, Coenzyme Q10, and Creatine. Meat Sci. 2004, 66, 629–637. [Google Scholar] [CrossRef]
- Artioli, G.G.; Gualano, B.; Smith, A.; Stout, J.; Lancha, A.H. Role of β-Alanine Supplementation on Muscle Carnosine and Exercise Performance. Med. Sci. Sports Exerc. 2010, 42, 1162–1173. [Google Scholar] [CrossRef]
- Harris, R.C.; Tallon, M.J.; Dunnett, M.; Boobis, L.; Coakley, J.; Kim, H.J.; Fallowfield, J.L.; Hill, C.A.; Sale, C.; Wise, J.A. The Absorption of Orally Supplied β-Alanine and Its Effect on Muscle Carnosine Synthesis in Human Vastus Lateralis. Amino Acids 2006, 30, 279–289. [Google Scholar] [CrossRef]
- Schober, P.; Boer, C.; Schwarte, L.A. Correlation Coefficients: Appropriate Use and Interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Antonelo, D.S.; Gómez, J.F.M.; Cônsolo, N.R.B.; Mazon, M.R.; Nubiato, K.E.Z.; De Souza, C.; Saran, A.; Silva, S.D.L.E. Effects of Combining Immunocastration and β-Adrenergic Agonists on the Blood Metabolites and Their Correlations with Performance and Carcass Traits of Finished Nellore Cattle. Rev. Bras. Zootec. 2020, 49, 13–20. [Google Scholar] [CrossRef]
- Novais, F.J.; Pires, P.R.L.; Alexandre, P.A.; Dromms, R.A.; Iglesias, A.H.; Ferraz, J.B.S.; Styczynski, M.P.W.; Fukumasu, H. Identification of a Metabolomic Signature Associated with Feed Efficiency in Beef Cattle. BMC Genom. 2019, 20, 8. [Google Scholar] [CrossRef] [Green Version]
- Antonelo, D.; Gómez, J.F.M.; Cônsolo, N.R.B.; Beline, M.; Colnago, L.A.; Schilling, W.; Zhang, X.; Suman, S.P.; Gerrard, D.E.; Balieiro, J.C.C.; et al. Metabolites and Metabolic Pathways Correlated With Beef Tenderness. Meat Muscle Biol. 2020, 4, 19–20. [Google Scholar] [CrossRef]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal Programming of Skeletal Muscle Development in Ruminant Animals. J. Anim. Sci. 2010, 88, E51–E60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schalch Junior, F.J.; Polizel, G.H.G.; Cançado, F.A.C.Q.; Fernandes, A.C.; Mortari, I.; Pires, P.R.L.; Fukumasu, H.; Santana, M.H.d.A.; Saran Netto, A. Prenatal Supplementation in Beef Cattle and Its Effects on Plasma Metabolome of Dams and Calves. Metabolites 2022, 12, 347. [Google Scholar] [CrossRef] [PubMed]
- Polizel, G.H.G.; Strefezzi, R.D.F.; Cracco, R.C.; Fernandes, A.C.; Zuca, C.B.; Castellar, H.H.; Baldin, G.C.; Santana, M.H. de A. Effects of Different Maternal Nutrition Approaches on Weight Gain and on Adipose and Muscle Tissue Development of Young Bulls in the Rearing Phase. Trop. Anim. Health Prod. 2021, 53, 536. [Google Scholar] [CrossRef] [PubMed]
- Polizel, G.H.G.; Fantinato-Neto, P.; Rangel, R.B.; Grigoletto, L.; Bussiman, F.D.O.; Cracco, R.C.; Garcia, N.P.; Ruy, I.M.; Ferraz, J.B.S.; de Santana, M.H.A. Evaluation of Reproductive Traits and the Effect of Nutrigenetics on Bulls Submitted to Fetal Programming. Livest. Sci. 2021, 247, 104487. [Google Scholar] [CrossRef]
- Zukunft, S.; Prehn, C.; Röhring, C.; Möller, G.; Hrabě de Angelis, M.; Adamski, J.; Tokarz, J. High-Throughput Extraction and Quantification Method for Targeted Metabolomics in Murine Tissues. Metabolomics 2018, 14, 18. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits (kg) | NP | PP | FP | SEM | p Value |
|---|---|---|---|---|---|
| BW | 591.2 ± 40.05 | 602.6 ± 49.65 | 597.4 ± 51.06 | 5.858 | 0.685 |
| LW | 7.227 ± 0.734 | 7.295 ± 0.779 | 7.286 ± 0.714 | 0.092 | 0.924 |
| Metabolite | NP | PP | FP | SEM | p Value |
|---|---|---|---|---|---|
| Gly | 1232 ± 151.5 ab | 1099 ± 86.60 a | 1325 ± 187.3 b | 57.91 | 0.024 |
| C14:2-OH | 0.006 ± 0.004 a | 0.002 ± 0.005 ab | 0.001 ± 0.001 b | 0.001 | 0.042 |
| Alpha-aaa | 8.155 ± 2.106 a | 8.432 ± 3.799 a | 4.457 ± 1.773 b | 1.045 | 0.043 |
| Carnosine | 122.6 ± 29.84 a | 107.3 ± 27.33 ab | 84.36 ± 12.27 b | 9.451 | 0.050 |
| Ingredients (Dry Matter) | Mineral Supplement | Protein-Energy Supplement |
| Corn (%) | 35.00 | 60.00 |
| Soybean meal (%) | - | 30.00 |
| Dicalcium phosphate (%) | 10.00 | - |
| Urea 45% (%) | - | 2.50 |
| Salt (%) | 30.00 | 5.00 |
| Minerthal 160 MD (%) * | 25.00 | 2.50 |
| Nutrients | Mineral Supplement | Protein-Energy Supplement |
| Total digestible nutrients (%) | 26.76 | 67.55 |
| Crude protein (%) | 2.79 | 24.78 |
| Non-protein nitrogen (%) | - | 7.03 |
| Acid detergent fiber (%) | 1.25 | 4.76 |
| Neutral detergent fiber (%) | 4.29 | 11.24 |
| Fat (%) | 1.26 | 2.61 |
| Calcium (g/kg) | 74.11 | 6.20 |
| Phosphorus (g/kg) | 59.38 | 7.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polizel, G.H.G.; Cançado, F.A.C.Q.; Dias, E.F.F.; Fernandes, A.C.; Cracco, R.C.; Carmona, B.T.; Castellar, H.H.; Poleti, M.D.; Santana, M.H.d.A. Effects of Different Prenatal Nutrition Strategies on the Liver Metabolome of Bulls and Its Correlation with Body and Liver Weight. Metabolites 2022, 12, 441. https://doi.org/10.3390/metabo12050441
Polizel GHG, Cançado FACQ, Dias EFF, Fernandes AC, Cracco RC, Carmona BT, Castellar HH, Poleti MD, Santana MHdA. Effects of Different Prenatal Nutrition Strategies on the Liver Metabolome of Bulls and Its Correlation with Body and Liver Weight. Metabolites. 2022; 12(5):441. https://doi.org/10.3390/metabo12050441
Chicago/Turabian StylePolizel, Guilherme Henrique Gebim, Fernando Augusto Correia Queiroz Cançado, Evandro Fernando Ferreira Dias, Arícia Christofaro Fernandes, Roberta Cavalcante Cracco, Brenda Tonsic Carmona, Henrique Hespanhol Castellar, Mirele Daiana Poleti, and Miguel Henrique de Almeida Santana. 2022. "Effects of Different Prenatal Nutrition Strategies on the Liver Metabolome of Bulls and Its Correlation with Body and Liver Weight" Metabolites 12, no. 5: 441. https://doi.org/10.3390/metabo12050441
APA StylePolizel, G. H. G., Cançado, F. A. C. Q., Dias, E. F. F., Fernandes, A. C., Cracco, R. C., Carmona, B. T., Castellar, H. H., Poleti, M. D., & Santana, M. H. d. A. (2022). Effects of Different Prenatal Nutrition Strategies on the Liver Metabolome of Bulls and Its Correlation with Body and Liver Weight. Metabolites, 12(5), 441. https://doi.org/10.3390/metabo12050441