
The Role of Quorum Sensing Molecules in Bacterial–Plant Interactions


Abstract
:1. Introduction
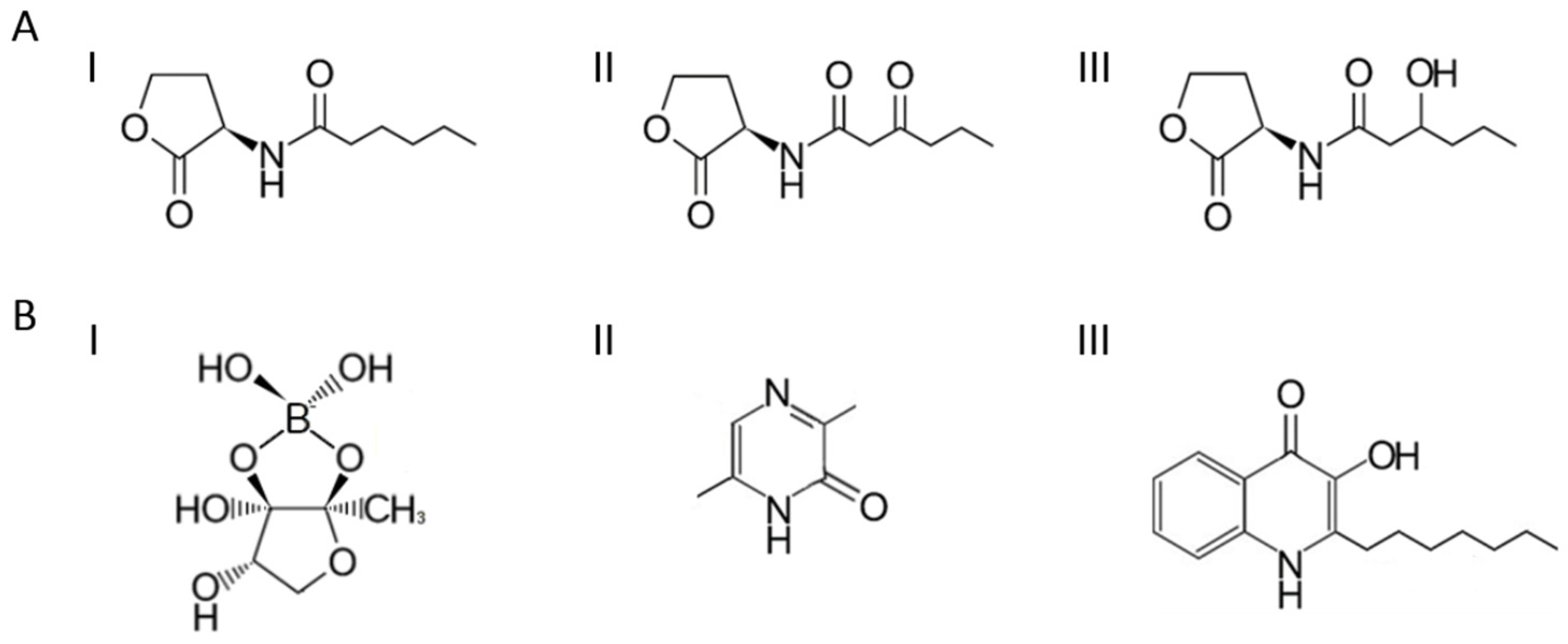
2. Types and Importance of Signaling Molecules
3. Mechanism of the Synthesis and Action of Autoinducers
Mechanism of Action of AI-1 Autoinducers in Gram-Negative Bacteria
4. QS System Inhibitors and Degrading Enzymes
4.1. Use of Genetic Engineering and Protein Engineering Methods to Obtain Stable and Active Quorum Quenching Enzymes
4.2. QSI Applications in Agriculture
5. Effects of AHL Compounds on Plant Growth and Health
5.1. Effects of AHL on Plant Root Morphology
5.2. Effects of AHL on the Expression of Plant Genes Associated with Defense Mechanisms
5.3. Enhancing Resistance to Pathogens and Insects
5.4. Participation of QS Molecules in the Nitrogen Cycle
6. The Use of Metagenomics to Study QS and QSI (or QQ) Diversity
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Nealson, K.H.; Hastings, J.W. Bacterial Bioluminescence: Its Control and Ecological Significance. Microbiol. Rev. 1979, 43, 496–518. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum Sensing Signal-Response Systems in Gram-Negative Bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Ruhal, R.; Kataria, R. Biofilm Patterns in Gram-Positive and Gram-Negative Bacteria. Microbiol. Res. 2021, 251, 126829. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, X.; Zheng, J.; Zou, Y.; Qiu, S.; Dai, Y. AHL-Mediated Quorum Sensing to Regulate Bacterial Substance and Energy Metabolism: A Review. Microbiol. Res. 2022, 262, 127102. [Google Scholar] [CrossRef]
- Wagner, V.E.; Bushnell, D.; Passador, L.; Brooks, A.I.; Iglewski, B.H. Microarray Analysis of Pseudomonas aeruginosa Quorum-Sensing Regulons: Effects of Growth Phase and Environment. J. Bacteriol. 2003, 185, 2080–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, T.G.; Fuqua, C. What’s in a Name? The Semantics of Quorum Sensing. Trends Microbiol. 2010, 18, 383–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Bassler, B.L. Bacterial Quorum Sensing in Complex and Dynamically Changing Environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Hansen, M.R.; Jakobsen, T.H.; Bang, C.G.; Cohrt, A.E.; Hansen, C.L.; Clausen, J.W.; le Quement, S.T.; Tolker-Nielsen, T.; Givskov, M.; Nielsen, T.E. Triazole-Containing N-Acyl Homoserine Lactones Targeting the Quorum Sensing System in Pseudomonas aeruginosa. Bioorg. Med. Chem. 2015, 23, 1638–1650. [Google Scholar] [CrossRef]
- Williams, P.; Winzer, K.; Chan, W.C.; Cámara, M. Look Who’s Talking: Communication and Quorum Sensing in the Bacterial World. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1119–1134. [Google Scholar] [CrossRef]
- Biswa, P.; Doble, M. Production of Acylated Homoserine Lactone by Gram-Positive Bacteria Isolated from Marine Water. FEMS Microbiol. Lett. 2013, 343, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Bose, U.; Ortori, C.A.; Sarmad, S.; Barrett, D.A.; Hewavitharana, A.K.; Hodson, M.P.; Fuerst, J.A.; Shaw, P.N. Production of N-Acyl Homoserine Lactones by the Sponge-Associated Marine Actinobacteria Salinispora arenicola and Salinispora pacifica. FEMS Microbiol. Lett. 2017, 364, fnx002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, J.C.; Watters, C.; Wong, H.L.; Visscher, P.T.; Burns, B.P. Isolation of Novel Quorum-Sensing Active Bacteria from Microbial Mats in Shark Bay Australia. FEMS Microbiol. Ecol. 2019, 95, fiz035. [Google Scholar] [CrossRef] [PubMed]
- Yin Wong, S.; Charlesworth, J.C.; Benaud, N.; Burns, B.P.; Ferrari, B.C. Communication within East Antarctic Soil Bacteria. Appl. Environ. Microbiol. 2019, 86, e01968-19. [Google Scholar] [CrossRef]
- Li, Z.; Nair, S.K. Quorum Sensing: How Bacteria Can Coordinate Activity and Synchronize Their Response to External Signals? Protein Sci. 2012, 21, 1403–1417. [Google Scholar] [CrossRef]
- Schikora, A.; Schenk, S.T.; Stein, E.; Molitor, A.; Zuccaro, A.; Kogel, K.H. N-Acyl-Homoserine Lactone Confers Resistance toward Biotrophic and Hemibiotrophic Pathogens via Altered Activation of AtMPK6. Plant Physiol. 2011, 157, 1407–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Todd, C.D.; Desikan, R.; Yang, Y.; Hu, X. N-3-Oxo-Decanoyl-l-Homoserine-Lactone Activates Auxin-Induced Adventitious Root Formation via Hydrogen Peroxide- and Nitric Oxide-Dependent Cyclic GMP Signaling in Mung Bean. Plant Physiol. 2012, 158, 725–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhao, Q.; Jia, Z.; Song, C.; Huang, Y.; Ma, H.; Song, S. N-3-Oxo-Octanoyl-Homoserine Lactone-Mediated Priming of Resistance to Pseudomonas syringae Requires. the Salicylic Acid Signaling Pathway in Arabidopsis thaliana. BMC Plant Biol. 2020, 20, 38. [Google Scholar] [CrossRef] [PubMed]
- Dow, L. How Do Quorum-Sensing Signals Mediate Algae–Bacteria Interactions? Microorganisms 2021, 9, 1391. [Google Scholar] [CrossRef]
- Churchill, M.E.A.; Chen, L. Structural Basis of Acyl-Homoserine Lactone-Dependent Signaling. Chem. Rev. 2011, 111, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Decho, A.W.; Norman, R.S.; Visscher, P.T. Quorum Sensing in Natural Environments: Emerging Views from Microbial Mats. Trends Microbiol. 2010, 18, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Prescott, R.D.; Decho, A.W. Flexibility and Adaptability of Quorum Sensing in Nature. Trends Microbiol. 2020, 28, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Q.; Dong, D.; Hu, H.; Wu, B.; Ren, H. AHLs-Mediated Quorum Sensing Threshold and Its Response towards Initial Adhesion of Wastewater Biofilms. Water Res. 2021, 194, 116925. [Google Scholar] [CrossRef] [PubMed]
- Boşgelmez-Tinaz, G.; Ulusoy, S. Characterization of N-Butanoyl-l-Homoserine Lactone (C4-HSL) Deficient Clinical Isolates of Pseudomonas aeruginosa. Microb. Pathog. 2008, 44, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Werner, K.M.; Perez, L.J.; Ghosh, R.; Semmelhack, M.F.; Bassler, B.L. Caenorhabditis elegans Recognizes a Bacterial Quorum-Sensing Signal Molecule through Theawcon Neuron. J. Biol. Chem. 2014, 289, 26566–26573. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, A.L.; Taylor, T.A.; Beatty, J.T.; Greenberg, E.P. Long-Chain Acyl-Homoserine Lactone Quorum-Sensing Regulation of Rhodobacter capsulatus Gene Transfer Agent Production. J. Bacteriol. 2002, 184, 6515–6521. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.F.; Purmal, K.; Chin, S.; Chan, X.Y.; Chan, K.G. Long Chain N-Acyl Homoserine Lactone Production by Enterobacter sp. Isolated from Human Tongue Surfaces. Sensors 2012, 12, 14307–14314. [Google Scholar] [CrossRef] [Green Version]
- Lau, Y.Y.; Sulaiman, J.; Chen, J.W.; Yin, W.F.; Chan, K.G. Quorum Sensing Activity of Enterobacter Asburiae Isolated from Lettuce Leaves. Sensors 2013, 13, 14189–14199. [Google Scholar] [CrossRef]
- dos Reis Ponce, A.; Martins, M.L.; de Araujo, E.F.; Mantovani, H.C.; Vanetti, M.C.D. AiiA Quorum-Sensing Quenching Controls Proteolytic Activity and Biofilm Formation by Enterobacter Cloacae. Curr. Microbiol. 2012, 65, 758–763. [Google Scholar] [CrossRef]
- Rice, S.A.; Koh, K.S.; Queck, S.Y.; Labbate, M.; Lam, K.W.; Kjelleberg, S. Biofilm Formation and Sloughing in Serratia Marcescens Are Controlled by Quorum Sensing and Nutrient Cues. J. Bacteriol. 2005, 187, 3477–3485. [Google Scholar] [CrossRef]
- Papadopoulos, C.J.; Carson, C.F.; Chang, B.J.; Riley, T.V. Role of the MexAB-OprM Efflux Pump of Pseudomonas aeruginosa in Tolerance to Tea Tree (Melaleuca alternifolia) Oil and Its Monoterpene Components Terpinen-4-Ol, 1,8-Cineole, and α-Terpineol ∇. Appl. Environ. Microbiol. 2008, 74, 1932–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Byun, T.; Dussen, H.-J.; Duke, K.R. Degradation of N-acylhomoserine lactones, the bacterial quorum-sensing molecules, by acylase. J. Biotechnol. 2003, 101, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y.Z.; Zhao, B.H.; Zhang, K.; Liang, D.B.; Wei, J.; Wang, X.J.; Li, J.; Chen, G.H. Effects of PH on AHL Signal Release and Properties of ANAMMOX Granules with Different Biomass Densities. Environ. Sci. 2019, 5, 1723–1735. [Google Scholar] [CrossRef]
- Ziegler, E.W.; Brown, A.B.; Nesnas, N.; Palmer, A.G. Abiotic Hydrolysis Kinetics of N-Acyl-L-Homoserine Lactones: Natural Silencing of Bacterial Quorum Sensing Signals. Eur. J. Org. Chem. 2019, 2019, 2850–2856. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Ai, Q.; Tan, L. Autoinducer-2 promotes Pseudomonas aeruginosa PAO1 acute lung infection via the IL-17A pathway. Front. Microbiol. 2022, 13, 948646. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Capalash, N.; Sharma, P. Quorum Sensing in Thermophiles: Prevalence of Autoinducer-2 System. BMC Microbiol. 2018, 18, 62. [Google Scholar] [CrossRef]
- Gutierrez, J.A.; Crowder, T.; Rinaldo-Matthis, A.; Ho, M.C.; Almo, S.C.; Schramm, V.L. Transition State Analogs of 5′-Methylthioadenosine Nucleosidase Disrupt Quorum Sensing. Nat. Chem. Biol. 2009, 5, 251–257. [Google Scholar] [CrossRef]
- Kim, C.S.; Gatsios, A.; Cuesta, S.; Lam, Y.C.; Wei, Z.; Chen, H.; Russell, R.M.; Shine, E.E.; Wang, R.; Wyche, T.P.; et al. Characterization of Autoinducer-3 Structure and Biosynthesis in E. coli. ACS Cent. Sci. 2020, 6, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Reading, N.C.; Sperandio, V. Quorum Sensing: The Many Languages of Bacteria. Quorum Sensing: The Many Languages of Bacteria. FEMS Microbiol. Lett. 2006, 254, 1–11. [Google Scholar] [CrossRef]
- Sperandio, V.; Torres, A.G.; Jarvis, B.; Nataro, J.P.; Kaper, J.B. Bacteria-host communication: The language of hormones. Proc. Natl. Acad. Sci. USA 2003, 100, 8951–8956. [Google Scholar] [CrossRef]
- Flavier, A.B.; Clough, S.J.; Schell, M.A.; Denny, T.P. Identification of 3-hydroxypalmitic acid methyl ester as a novel autoregulator controlling virulence in Ralstonia solanacearum. Mol. Microbiol. 1997, 26, 251–259. [Google Scholar] [CrossRef] [PubMed]
- LaSarre, B.; Federle, M.J. Exploiting Quorum Sensing To Confuse Bacterial Pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Jayaraman, A.; Wood, T.K. Indole Is an Inter-Species Biofilm Signal Mediated by SdiA. BMC Microbiol. 2007, 7, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobias, N.J.; Brehm, J.; Kresovic, D.; Brameyer, S.; Bode, H.B.; Heermann, R. New Vocabulary for Bacterial Communication. ChemBiochem 2020, 21, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Kumar, L.; Patel, S.K.S.; Kharga, K.; Kumar, R.; Kumar, P.; Pandohee, J.; Kulshresha, S.; Harjai, K.; Chhibber, S. Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria. Molecules 2022, 27, 7584. [Google Scholar] [CrossRef]
- Whitehead, N.A.; Barnard, A.M.L.; Slater, H.; Simpson, N.J.L.; Salmond, G.P.C. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef]
- Billot, R.; Plener, L.; Jacquet, P.; Elias, M.; Chabrière, E.; Daudé, D. Engineering Acyl-Homoserine Lactone-Interfering Enzymes toward Bacterial Control. J. Biol. Chem. 2020, 295, 12993–13007. [Google Scholar] [CrossRef]
- Montebello, A.N.; Brecht, R.M.; Turner, R.D.; Ghali, M.; Pu, X.; Nagarajan, R. Acyl-ACP Substrate Recognition in Burkholderia Mallei Bmai1 Acyl-Homoserine Lactone Synthase. Biochemistry 2014, 53, 6231–6242. [Google Scholar] [CrossRef] [Green Version]
- Guan, R.; Ho, M.C.; Almo, S.C.; Schramm, V.L. Methylthioinosine Phosphorylase from Pseudomonas aeruginosa. Structure and Annotation of a Novel Enzyme in Quorum Sensing. Biochemistry 2011, 50, 1247–1254. [Google Scholar] [CrossRef] [Green Version]
- Winzer, K.; Hardie, K.R.; Burgess, N.; Doherty, N.; Kirke, D.; Holden, M.T.G.; Linforth, R.; Cornell, K.A.; Taylor, A.J.; Hill, P.J.; et al. LuxS: Its role in central metabolism and the in vitro synthesis of 4-hydroxy-5-methyl-3(2H)-furanone. Microbiology 2002, 148 Pt 4, 909922. [Google Scholar] [CrossRef]
- Verma, S.C.; Miyashiro, T. Quorum Sensing in the Squid-Vibrio Symbiosis. Int. J. Mol. Sci. 2013, 14, 16386–16401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimbrough, J.H.; Stabb, E.V. Comparative Analysis Reveals Regulatory Motifs at the AinS/AinR Pheromone-Signaling Locus of Vibrio fischeri. Sci. Rep. 2017, 7, 11734. [Google Scholar] [CrossRef] [PubMed]
- Lupp, C.; Ruby, E.G. Vibrio Fischeri Uses Two Quorum-Sensing Systems for the Regulation of Early and Late Colonization Factors. J. Bacteriol. 2005, 187, 3620–3629. [Google Scholar] [CrossRef] [Green Version]
- Girard, L. Quorum Sensing in Vibrio spp.: The Complexity of Multiple Signalling Molecules in Marine and Aquatic Environments. Crit. Rev. Microbiol. 2019, 45, 451–471. [Google Scholar] [CrossRef]
- Lipa, P.; Kozieł, M.; Janczarek, M. Zjawisko Quorum Sensing bakterii Gram-ujemnych: Cząsteczki sygnałowe i inhibitory oraz ich potencjalne zastosowanie terapeutyczne [Quorum sensing in Gram-negative bacteria: Signal molecules, inhibitors and their potential therapeutic application]. Postepy Biochem. 2017, 63, 242–260. [Google Scholar]
- Li, Y.; Qu, H.P.; Liu, J.L.; Wan, H.Y. Correlation between Group Behavior and Quorum Sensing in Pseudomonas aeruginosa Isolated from Patients with Hospitalacquired Pneumonia. J. Thorac. Dis. 2014, 6, 810–817. [Google Scholar] [CrossRef]
- Lee, J.; Zhang, L. The Hierarchy Quorum Sensing Network in Pseudomonas aeruginosa. Protein Cell 2015, 6, 26–41. [Google Scholar] [CrossRef] [Green Version]
- Soukarieh, F.; Williams, P.; Stocks, M.J.; Cámara, M. Pseudomonas aeruginosa Quorum Sensing Systems as Drug Discovery Targets: Current Position and Future Perspectives. J. Med. Chem. 2018, 61, 10385–10402. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Mao, S.; Wang, H.; Ye, X. The Molecular Architecture of Pseudomonas aeruginosa Quorum-Sensing Inhibitors. Mar. Drugs 2022, 20, 488. [Google Scholar] [CrossRef]
- Lang, J.; Faure, D. Functions and Regulation of Quorum-Sensing in Agrobacterium Tumefaciens. Front. Plant Sci. 2014, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Matejczyk, M.; Suchowierska, M. Charakterystyka zjawiska quorum sensing i jego znaczenie w aspekcie formowania i funkcjonowania biofilmu w inżynierii środowiska, budownictwie, medycynie oraz gospodarstwie domowym. Civ. Environ. Eng. 2011, 2, 71–75. [Google Scholar]
- Jones, S.; Yu, B.; Bainton, N.J.; Birdsall, M.; Bycroft, B.W.; Chhabra, S.R.; Cox, A.J.R.; Golby, P.; Reeves, P.J.; Stephens, S. The Lux Autoinducer Regulates the Production of Exoenzyme Virulence Determinants in Erwinia carotovora and Pseudomonas aeruginosa. EMBO J. 1993, 12, 2477–2482. [Google Scholar] [CrossRef] [PubMed]
- van Houdt, R.; Givskov, M.; Michiels, C.W. Quorum Sensing in Serratia. FEMS Microbiol. Rev. 2007, 31, 407–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalia, V.C. Quorum Sensing Inhibitors: An Overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Kang, Y.C.; Lee, J.K. Quorum Sensing Inhibitors as Antipathogens: Biotechnological Applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef]
- Bai, Y.-B.; Shi, M.-Y.; Wang, W.-W.; Wu, L.-Y.; Bai, Y.-T.; Li, B.; Zhou, X.-Z.; Zhang, J.-Y. Novel Quorum Sensing Inhibitor Echinatin as an Antibacterial Synergist against Escherichia coli. Front. Microbiol. 2022, 13, 1003692. [Google Scholar] [CrossRef]
- Joshi, J.R.; Khazanov, N.; Senderowitz, H.; Burdman, S.; Lipsky, A.; Yedidia, I. Plant Phenolic Volatiles Inhibit Quorum Sensing in Pectobacteria and Reduce Their Virulence by Potential Binding to ExpI and ExpR Proteins. Sci. Rep. 2016, 6, 38126. [Google Scholar] [CrossRef] [Green Version]
- Pun, M.; Khazanov, N.; Galsurker, O.; Weitman, M.; Kerem, Z.; Senderowitz, H.; Yedidia, I. Phloretin, an Apple Phytoalexin, Affects the Virulence and Fitness of Pectobacterium Brasiliense by Interfering with Quorum-Sensing. Front. Plant Sci. 2021, 12, 1261. [Google Scholar] [CrossRef]
- Zheng, C.J.; Sohn, M.J.; Lee, S.; Kim, W.G. Meleagrin, a New Fabi Inhibitor from Penicillium chryosogenum with at Least One Additional Mode of Action. PLoS ONE 2013, 8, e0078922. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, S.; Teplitski, M.; Bayer, M.; Gunasekera, S.; Proksch, P.; Paul, V.J. Inhibition of Marine Biofouling by Bacterial Quorum Sensing Inhibitors. Biofouling 2011, 27, 893–905. [Google Scholar] [CrossRef]
- Jack, A.A.; Khan, S.; Powell, L.C.; Pritchard, M.F.; Beck, K.; Sadh, H.; Sutton, L.; Cavaliere, A.; Florance, H.; Rye, P.D.; et al. Alginate Oligosaccharide-Induced Modification of the LasI-LasR and RhlI-RhlR Quorum-Sensing Systems in Pseudomonas aeruginosa. AntiMicrob. Agents Chemother. 2018, 62, e02318-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Onsøyen, E.; Myrvold, R.; Howe, R.A.; Walsh, T.R.; Hill, K.E.; et al. Overcoming drug resistance with alginate oligosaccharides able to potentiate the action of selected antibiotics. AntiMicrob. Agents Chemother. 2012, 56, 5134–5141. [Google Scholar] [CrossRef] [PubMed]
- Karnjana, K.; Nobsathian, S.; Soowannayan, C.; Zhao, W.; Tang, Y.J.; Wongprasert, K. Purification and Evaluation of N-Benzyl Cinnamamide from Red Seaweed Gracilaria Fisheri as an Inhibitor of Vibrio harveyi AI-2 Quorum Sensing. Mar. Drugs 2020, 18, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.P.; Zeng, H.; Wan, C.X.; Zhou, Z.B. Amicoumacins from a Desert Bacterium: Quorum Sensing Inhibitor against Chromobacterium violaceum. Nat. Prod Res. 2020, 32, 5508–5512. [Google Scholar] [CrossRef]
- Kamarudheen, N.; Rao, K.V.B. Fatty Acyl Compounds from Marine Streptomyces Griseoincarnatus Strain HK12 against Two Major Bio-Film Forming Nosocomial Pathogens; an in Vitro and in Silico Approach. Microb. Pathog. 2019, 127, 121–130. [Google Scholar] [CrossRef]
- Kapadia, C.; Kachhdia, R.; Singh, S.; Gandhi, K.; Poczai, P.; Alfarraj, S.; Ansari, M.J.; Gafur, A.; Sayyed, R.Z. Pseudomonas aeruginosa inhibits quorum-sensing mechanisms of soft rot pathogen Lelliottia amnigena RCE to regulate its virulence factors and biofilm formation. Front. Microbiol. 2022, 13, 977669. [Google Scholar] [CrossRef]
- Taha, M.N.; Saafan, A.E.; Ahmedy, A.; el Gebaly, E.; Khairalla, A.S. Two Novel Synthetic Peptides Inhibit Quorum Sensing-Dependent Biofilm Formation and Some Virulence Factors in Pseudomonas aeruginosa PAO1. J. Microbiol. 2019, 57, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.-M.; Cheng, C.; Liu, T.; Zhao, Y.-L.; Guo, B.; Tang, L.; Yang, Y.-Y. Synthesis and Antibiofilm Evaluation of N-Acyl-2-Aminopyrimidine Derivatives against Acinetobacter baumannii. Bioorg. Med. Chem. 2022, 76, 117095. [Google Scholar] [CrossRef]
- Yoshihara, A.; Shimatani, M.; Sakata, M.; Takemura, C.; Senuma, W.; Hikichi, Y.; Kai, K. Quorum Sensing Inhibition Attenuates the Virulence of the Plant Pathogen Ralstonia solanacearum Species Complex. ACS Chem. Biol. 2020, 15, 3050–3059. [Google Scholar] [CrossRef]
- Manefield, M.; de Nys, R.; Read, R.; Steinberg, P.; Kjelleberg, S.; Wales, S. Evidence That Halogenated Furanones from Delisea Pulchra Inhibit Acylated Homoserine Lactone (AHL)-Mediated Gene Expression by Displacing the AHL Signal from Its Receptor Protein. Microbiology 1999, 145, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Shi, Y.; Zeng, G.; Gu, Y.; Chen, G.; Shi, L.; Hu, Y.; Tang, B.; Zhou, J. Acyl-Homoserine Lactone-Based Quorum Sensing and Quorum Quenching Hold Promise to Determine the Performance of Biological Wastewater Treatments: An Overview. Chemosphere 2016, 157, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Delago, A.; Mandabi, A.; Meijler, M.M. Natural Quorum Sensing Inhibitors—Small Molecules, Big Messages. Isr J. Chem. 2016, 56, 310–320. [Google Scholar] [CrossRef]
- Lyons, T.; Gahan, C.G.M.; O’Sullivan, T.P. Structure-Activity Relationships of Furanones, Dihydropyrrolones and Thiophenones as Potential Quorum Sensing Inhibitors. Future Med. Chem. 2020, 12, 1925–1943. [Google Scholar] [CrossRef]
- Joshi, J.R.; Khazanov, N.; Charkowski, A.; Faigenboim, A.; Senderowitz, H.; Yedidia, I. Interkingdom Signaling Interference: The Effect of Plant-Derived Small Molecules on Quorum Sensing in Plant-Pathogenic Bacteria. Annu. Rev. Phytopathol. 2021, 59, 153–190. [Google Scholar] [CrossRef]
- Bouyahya, A.; Dakka, N.; Et-Touys, A.; Abrini, J.; Bakri, Y. Medicinal Plant Products Targeting Quorum Sensing for Combating Bacterial Infections. Asian Pac. J. Trop. Med. 2017, 10, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Deryabin, D.; Galadzhieva, A.; Kosyan, D.; Duskaev, G. Plant-Derived Inhibitors of AHL-Mediated Quorum Sensing in Bacteria: Modes of Action. Int. J. Mol. Sci. 2019, 20, 5588. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Gao, Y.; Chen, X.; Yu, Z.; Li, X. Quorum Quenching Enzymes and Their Application in Degrading Signal Molecules to Block Quorum Sensing-Dependent Infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef]
- Fetzner, S. Quorum Quenching Enzymes. J. Biotechnol. 2015, 201, 2–14. [Google Scholar] [CrossRef]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum Sensing and Quorum Quenching: The Yin and Yang of Bacterial Communication. ChemBioChem 2009, 10, 205–216. [Google Scholar] [CrossRef]
- Lord, D.M.; Baran, A.U.; Wood, T.K.; Peti, W.; Page, R. BdcA, a Protein Important for Escherichia coli Biofilm Dispersal, Is a Short-Chain Dehydrogenase/Reductase That Binds Specifically to NADPH. PLoS ONE 2014, 9, e0105751. [Google Scholar] [CrossRef]
- Molina, L.; Constantinescu, F.; Michel, L.; Reimmann, C.; Duffy, B.; Défago, G. Degradation of Pathogen Quorum-Sensing Molecules by Soil Bacteria: A Preventive and Curative Biological Control Mechanism. FEMS Microbiol. Ecol. 2003, 45, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.A.; Singh, A.K.; Nerurkar, A. Disrupting the Quorum Sensing Mediated Virulence in Soft Rot Causing Pectobacterium carotovorum by Marine Sponge Associated Bacillus sp. OA10. World J. Microbiol. Biotechnol 2021, 37, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Yu, Z.; Chu, W. Effect of Quorum-Quenching Bacterium Bacillus sp. QSI-1 on Protein Profiles and Extracellular Enzymatic Activities of Aeromonas hydrophila YJ-1. BMC Microbiol. 2019, 19, 135. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Zhou, S.; Zhu, W.; Zhuang, X. Quorum Quenching Bacteria Bacillus sp. QSI-1 Protect Zebrafish (Danio Rerio) from Aeromonas hydrophila Infection. Sci. Rep. 2014, 4, 5446. [Google Scholar] [CrossRef] [Green Version]
- Prazdnova, E.V.; Gorovtsov, A.V.; Vasilchenko, N.G.; Kulikov, M.P.; Statsenko, V.N.; Bogdanova, A.A.; Refeld, A.G.; Brislavskiy, Y.A.; Chistyakov, V.A.; Chikindas, M.L. Quorum-Sensing Inhibition by Gram-Positive Bacteria. Microorganisms 2022, 10, 350. [Google Scholar] [CrossRef]
- Barbey, C.; Crepin, A.; Bergeau, D.; Ouchiha, A.; Mijouin, L.; Taupin, L.; Orange, N.; Feuilloley, M.; Dufour, A.; Burini, J.-F.; et al. In Planta Biocontrol of Pectobacterium Atrosepticum by Rhodococcus Erythropolis Involves Silencing of Pathogen Communication by the Rhodococcal Gamma-Lactone Catabolic Pathway. PLoS ONE 2013, 8, e66642. [Google Scholar] [CrossRef]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum Quenching: Role in Nature and Applied Developments. FEMS Microbiol. Rev. 2015, 40, 86–116. [Google Scholar] [CrossRef]
- Sikdar, R.; Elias, M. Quorum Quenching Enzymes and Their Effects on Virulence, Biofilm, and Microbiomes: A Review of ReCent. Advances. Expert Rev. Anti Infect. Ther. 2020, 18, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Okano, C.; Murota, D.; Nasuno, E.; Iimura, K.I.; Kato, N. Effective Quorum Quenching with a Conformation-Stable Recombinant Lactonase Possessing a Hydrophilic Polymeric Shell Fabricated via Electrospinning. Mater. Sci. Eng. C 2019, 98, 437–444. [Google Scholar] [CrossRef]
- Sakr, M.M.; Elkhatib, W.F.; Aboshanab, K.M.; Mantawy, E.M.; Yassien, M.A.; Hassouna, N.A. In Vivo Evaluation of a Recombinant N-Acylhomoserine Lactonase Formulated in a Hydrogel Using a Murine Model Infected with MDR Pseudomonas aeruginosa Clinical Isolate, CCASUP2. AMB Express 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Zhang, B.; Zhuang, X.; Guo, L.; McLean, R.J.C.; Chu, W. Recombinant N-Acyl Homoserine Lactone-Lactonase AiiAQSI-1 Attenuates Aeromonas hydrophila Virulence Factors, Biofilm Formation and Reduces Mortality in Crucian carp. Mar. Drugs 2019, 17, 499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Cai, Y.; Xu, Z.; Fu, B.; Chen, Q.; Cui, Y.; Ruan, Z.; Liang, Y.; Peng, N.; Zhao, S. Heterologous Expression of AHL Lactonase AiiK by Lactobacillus casei MCJΔ1 with Great Quorum Quenching Ability against Aeromonas hydrophila AH-1 and AH-4. Microb. Cell Fact 2020, 19, 1–14. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, P.; Wu, J.; Yan, W.; Xie, S.; Sun, X.; Ye, B.C.; Chu, X. N-Acylhomoserine Lactonase-Based Hybrid Nanoflowers: A Novel and Practical Strategy to Control Plant Bacterial Diseases. J. Nanobiotechnol. 2022, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.D.; Seelig, B. Advances in the Directed Evolution of Proteins. Curr. Opin Chem. Biol. 2014, 22, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lin, J.; Zhang, Y.; Zhang, J.; Feng, T.; Li, H.; Wang, X.; Sun, Q.; Zhang, X.; Wang, Y. Activity Improvement and Vital Amino Acid Identification on the Marine-Derived Quorum Quenching Enzyme MOML by Protein Engineering. Mar. Drugs 2019, 17, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugayah, S.A.; Gerth, M.L. Engineering Quorum Quenching Enzymes: Progress and Perspectives. Biochem. Soc. Trans. 2019, 47, 793–800. [Google Scholar] [CrossRef]
- Aslanli, A.; Lyagin, I.; Efremenko, E. Novel Approach to Quorum Quenching: Rational Design of Antibacterials in Combination with Hexahistidine-Tagged Organophosphorus Hydrolase. Biol. Chem. 2018, 399, 869–879. [Google Scholar] [CrossRef]
- Jacquet, P.; Hiblot, J.; Daudé, D.; Bergonzi, C.; Gotthard, G.; Armstrong, N.; Chabrière, E.; Elias, M. Rational Engineering of a Native Hyperthermostable Lactonase into a Broad Spectrum Phosphotriesterase. Sci. Rep. 2017, 7, 16745. [Google Scholar] [CrossRef] [Green Version]
- Kyeong, H.H.; Kim, J.H.; Kim, H.S. Design of N-Acyl Homoserine Lactonase with High Substrate Specificity by a Rational Approach. Appl. Microbiol. Biotechnol. 2015, 99, 4735–4742. [Google Scholar] [CrossRef]
- Cassán, F.; Diaz-Zorita, M. Azospirillum sp. in Current Agriculture: From the Laboratory to the Field. Soil Biol. Biochem. 2016, 103, 117–130. [Google Scholar] [CrossRef]
- Vial, L.; Cuny, C.; Gluchoff-Fiasson, K.; Comte, G.; Oger, P.M.; Faure, D.; Dessaux, Y.; Bally, R.; Wisniewski-Dyé, F. N-Acyl-Homoserine Lactone-Mediated Quorum-Sensing in Azospirillum: An Exception Rather than a Rule. FEMS Microbiol. Ecol. 2006, 58, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Gualpa, J.; Lopez, G.; Nievas, S.; Coniglio, A.; Halliday, N.; Cámara, M.; Cassán, F. Azospirillum Brasilense Az39, a Model Rhizobacterium with AHL Quorum-Quenching Capacity. J. Appl. Microbiol. 2019, 126, 1850–1860. [Google Scholar] [CrossRef]
- Zhang, R.; Vivanco, J.M.; Shen, Q. The Unseen Rhizosphere Root–Soil–Microbe Interactions for Crop Production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef]
- Hartmann, A. Quorum Sensing N-Acyl-Homoserine Lactone Signal Molecules of Plant Beneficial Gram-Negative Rhizobacteria Support Plant Growth and Resistance to Pathogens. Rhizosphere 2020, 16, 100258. [Google Scholar] [CrossRef]
- von Rad, U.; Klein, I.; Dobrev, P.I.; Kottova, J.; Zazimalova, E.; Fekete, A.; Hartmann, A.; Schmitt-Kopplin, P.; Durner, J. Response of Arabidopsis thaliana to N-Hexanoyl-DL-Homoserine-Lactone, a Bacterial Quorum Sensing Molecule Produced in the Rhizosphere. Planta 2008, 229, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Liu, F.; Ma, H.; Hao, S.; Zhao, Q.; Bian, Z.; Jia, Z.; Song, S. Two G-Protein-Coupled-Receptor Candidates, Cand2 and Cand7, Are Involved in Arabidopsis Root Growth Mediated by the Bacterial Quorum-Sensing Signals N-Acyl-Homoserine Lactones. Biochem. Biophys. Res. Commun. 2012, 417, 991–995. [Google Scholar] [CrossRef]
- Liu, F.; Bian, Z.; Jia, Z.; Zhao, Q.; Song, S. The GCR1 and GPA1 Participate in Promotion of Arabidopsis Primary Root Elongation Induced by N-Acyl-Homoserine Lactones, the Bacterial Quorum-Sensing Signals. Mol. Plant-Microbe Interact. 2012, 25, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Zhang, C.; Jia, Z.; Huang, Y.; Li, H.; Song, S. Involvement of Calmodulin in Regulation of Primary Root Elongation by N-3-Oxo-Hexanoyl Homoserine Lactone in Arabidopsis thaliana. Front. Plant Sci. 2015, 5, 807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Li, M.; Jia, Z.; Liu, F.; Ma, H.; Huang, Y.; Song, S. AtMYB44 Positively Regulates the Enhanced Elongation of Primary Roots Induced by N-3-Oxo-Hexanoyl-Homoserine Lactone in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2016, 29, 774–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Bimerew, M.; Ma, Y.; Müller, H.; Ovadis, M.; Eberl, L.; Berg, G.; Chernin, L. Quorum-Sensing Signaling Is Required for Production of the Antibiotic Pyrrolnitrin in a Rhizospheric Biocontrol Strain of Serratia plymuthica. FEMS Microbiol. Lett. 2007, 270, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Ortíz-Castro, R.; Martínez-Trujillo, M.; López-Bucio, J. N-Acyl-L-Homoserine Lactones: A Class of Bacterial Quorum-Sensing Signals Alter Post-Embryonic Root Development in Arabidopsis thaliana. Plant Cell Environ. 2008, 31, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Veliz-Vallejos, D.F.; van Noorden, G.E.; Yuan, M.; Mathesius, U. A Sinorhizobium Meliloti-Specific N-Acyl Homoserine Lactone Quorum-Sensing Signal Increases Nodule Numbers in Medicago truncatula Independent of Autoregulation. Front. Plant Sci. 2014, 5, 551. [Google Scholar] [CrossRef] [PubMed]
- Schuhegger, R.; Ihring, A.; Gantner, S.; Bahnweg, G.; Knappe, C.; Vogg, G.; Hutzler, P.; Schmid, M.; van Breusegem, F.; Eberl, L.; et al. Induction of Systemic Resistance in Tomato by N-Acyl-L-Homoserine Lactone-Producing Rhizosphere Bacteria. Plant Cell Environ. 2006, 29, 909–918. [Google Scholar] [CrossRef]
- Schenk, S.T.; Schikora, A. AHL-Priming Functions via Oxylipin and Salicylic Acid. Front. Plant Sci. 2015, 5, 784. [Google Scholar] [CrossRef]
- Sieper, T.; Forczek, S.; Matucha, M.; Krämer, P.; Hartmann, A.; Schröder, P. N-Acyl-Homoserine Lactone Uptake and Systemic Transport in Barley Rest upon Active Parts of the Plant. New Phytol. 2014, 201, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Götz-Rösch, C.; Sieper, T.; Fekete, A.; Schmitt-Kopplin, P.; Hartmann, A.; Schröder, P. Influence of Bacterial N-Acyl-Homoserine Lactoneson Growth Parameters, Pigments, Antioxidative Capacities and the Xenobiotic Phase II Detoxification Enzymesin Barley and Yam Bean. Front. Plant Sci. 2015, 6, 205. [Google Scholar] [CrossRef] [Green Version]
- Schikora, A.; Schenk, S.T.; Hartmann, A. Beneficial Effects of Bacteria-Plant Communication Based on Quorum Sensing Molecules of the N-Acyl Homoserine Lactone Group. Plant Mol. Biol. 2016, 90, 605–612. [Google Scholar] [CrossRef]
- Schenk, S.T.; Hernández-Reyes, C.; Samans, B.; Stein, E.; Neumann, C.; Schikora, M.; Reichelt, M.; Mithöfer, A.; Becker, A.; Kogel, K.H.; et al. N-Acyl-Homoserine Lactone Primes Plants for Cell Wall Reinforcement and Induces Resistance to Bacterial Pathogens via the Salicylic Acid/Oxylipin Pathway. Plant Cell 2014, 26, 2708–2723. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Rothballer, M.; Schmid, M.; Esperschütz, J.; Hartmann, A. Acidovorax radicis sp. nov., a Wheat-Root-Colonizing Bacterium. Int. J. Syst. Evol. Microbiol. 2011, 61, 2589–2594. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Li, D.; Trost, E.; Mayer, K.F.; Corina, V.; Heller, W.; Schmid, M.; Hartmann, A.; Rothballer, M. Systemic Responses of Barley to the 3-Hydroxy-Decanoyl-Homoserine Lactone Producing Plant Beneficial Endophyte Acidovorax Radicis N35. Front. Plant Sci. 2016, 7, 1868. [Google Scholar] [CrossRef] [Green Version]
- Gantner, S.; Schmid, M.; Dürr, C.; Schuhegger, R.; Steidle, A.; Hutzler, P.; Langebartels, C.; Eberl, L.; Hartmann, A.; Dazzo, F.B. In Situ Quantitation of the Spatial Scale of Calling Distances and Population Density-Independent N-Acylhomoserine Lactone-Mediated Communication by Rhizobacteria Colonized on Plant Roots. FEMS Microbiol. Ecol. 2006, 56, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, A.; Grimm, M.; Ojiro, I.; Krumwiede, J.; Schikora, A. Impact of Quorum Sensing Molecules on Plant Growth and Immune System. Front. Microbiol. 2020, 11, 1545. [Google Scholar] [CrossRef] [PubMed]
- Wehner, G.; Schikora, A.; Ordon, F.; Will, T. Priming Negatively Affects Feeding Behaviour and Aphid Biomass of Rhopalosiphum padi on Barley. J. Pest Sci. 2021, 94, 1237–1247. [Google Scholar] [CrossRef]
- Moshynets, O.V.; Babenko, L.M.; Rogalsky, S.P.; Iungin, O.S.; Foster, J.; Kosakivska, I.V.; Potters, G.; Spiers, A.J. Priming Winter Wheat Seeds with the Bacterial Quorum Sensing Signal N-Hexanoyl-L-Homoserine Lactone (C6-HSL) Shows Potential to Improve Plant Growth and Seed Yield. PLoS ONE 2019, 14, e0209460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G.S.; Kumar, A.; Verma, N. Bacterial Homoserine Lactones as a Nanocomposite Fertilizer and Defense Regulator for Chickpeas. Environ. Sci. Nano 2019, 6, 1246–1258. [Google Scholar] [CrossRef]
- Deangelis, K.M.; Lindow, S.E.; Firestone, M.K. Bacterial Quorum Sensing and Nitrogen Cycling in Rhizosphere Soil. FEMS Microbiol. Ecol. 2008, 66, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Saraf, M.; Sharma, S. Quorum Sensing Enhances Nitrogen Uptake in Plant. In Soil Nitrogen Ecology, 1st ed.; (Soil Biology Book 62); Cruz, C., Vishwakarma, K., Choundhacy, D.K., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; Volume 62, pp. 371–388. [Google Scholar]
- Mellbye, B.L.; Giguere, A.T.; Bottomley, P.J.; Sayavedra-Soto, L.A. Quorum Quenching of Nitrobacter Winogradskyi Suggests That Quorum Sensing Regulates Fluxes of Nitrogen Oxide(s) during Nitrification. mBio 2016, 7, e01753-16. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Zhang, Y.; Shen, Q.; Gao, J.; Zhuang, G.; Zhuang, X. Effects of Exogenous Short-Chain N-Acyl Homoserine Lactone on Denitrifying Process of Paracoccus Denitrificans. J. Environ. Sci. (China) 2017, 54, 33–39. [Google Scholar] [CrossRef]
- Wang, N.; Gao, J.; Liu, Y.; Wang, Q.; Zhuang, X.; Zhuang, G. Realizing the Role of N-Acyl-Homoserine Lactone-Mediated Quorum Sensing in Nitrification and Denitrification: A Review. Chemosphere 2021, 274, 129970. [Google Scholar] [CrossRef]
- Zhu, Z.; Yang, Y.; Fang, A.; Lou, Y.; Xie, G.; Ren, N.; Xing, D. Quorum Sensing Systems Regulate Heterotrophic Nitrification-Aerobic Denitrification by Changing the Activity of Nitrogen-Cycling Enzymes. Environ. Sci. Ecotechnol. 2020, 2, 100026. [Google Scholar] [CrossRef]
- Kimura, N. Metagenomic Approaches to Understanding Phylogenetic Diversity in Quorum Sensing. Virulence 2014, 5, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Schipper, C.; Hornung, C.; Bijtenhoorn, P.; Quitschau, M.; Grond, S.; Streit, W.R. Metagenome-Derived Clones Encoding Two Novel Lactonase Family Proteins Involved in Biofilm Inhibition in Pseudomonas aefuginosa. Appl. Environ. Microbiol. 2009, 75, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Cardenas, J.A.; Manzano-Agugliaro, F. The Metagenomics Worldwide Research. Curr. Genet. 2017, 63, 819–829. [Google Scholar] [CrossRef]
- Torres, M.; Uroz, S.; Salto, R.; Fauchery, L.; Quesada, E.; Llamas, I. HqiA, a Novel Quorum-Quenching Enzyme Which Expands the AHL Lactonase Family. Sci. Rep. 2017, 7, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Yang, Y.; Zhu, X.Y.; Zhang, X.H.; Yu, M. Metagenomic Insights Into the Microbial Assemblage Capable of Quorum Sensing and Quorum Quenching in Particulate Organic Matter in the Yellow Sea. Front. Microbiol. 2021, 11, 602010. [Google Scholar] [CrossRef]
- Cirou, A.; Uroz, S.; Chapelle, E.; Latour, X.; Orange, N.; Faure, D.; Dessaux, Y. Quorum Sensing as a Target for Novel Biocontrol Strategies Directed at Pectobacterium. In ReCent. Developments in Management of Plant Diseases; Gisi, U., Chet, I., Gullino, M.L., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 121–131. [Google Scholar]
- Liu, X.; Cao, L.C.; Fan, X.J.; Liu, Y.H.; Xie, W. Engineering of a Thermostable Esterase Est816 to Improve Its Quorum-Quenching Activity and the Underlying Structural Basis. Sci. Rep. 2016, 6, 38137. [Google Scholar] [CrossRef] [Green Version]
- Riaz, K.; Elmerich, C.; Moreira, D.; Raffoux, A.; Dessaux, Y.; Faure, D. A Metagenomic Analysis of Soil Bacteria Extends the Diversity of Quorum-Quenching Lactonases. Environ. Microbiol. 2008, 10, 560–570. [Google Scholar] [CrossRef]
- Bijtenhoorn, P.; Mayerhofer, H.; Müller-Dieckmann, J.; Utpatel, C.; Schipper, C.; Hornung, C.; Szesny, M.; Grond, S.; Thürmer, A.; Brzuszkiewicz, E.; et al. A Novel Metagenomic Short-Chain Dehydrogenase/Reductase Attenuates Pseudomonas aeruginosa Biofilm Formation and Virulence on Caenorhabditis elegans. PLoS ONE 2011, 6, e26278. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
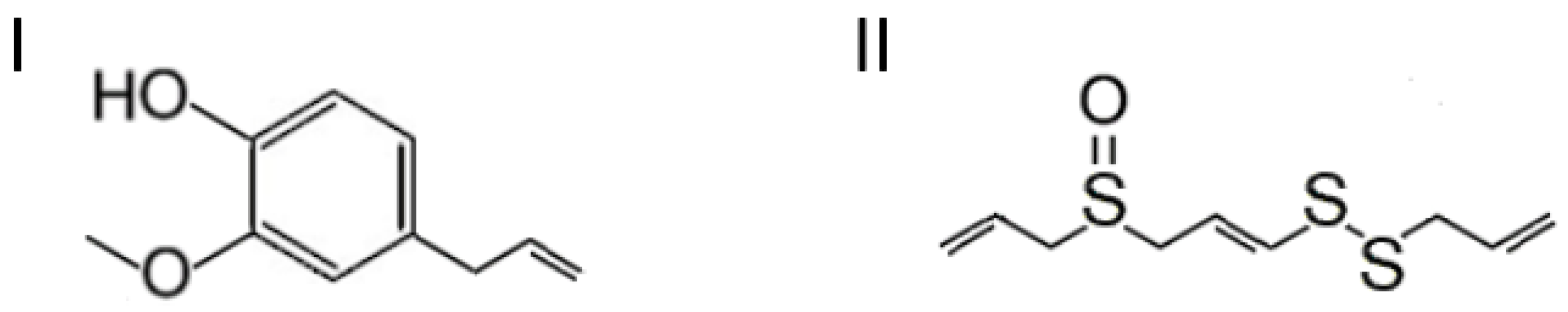{kind=link}
| Type of Source Organisms | Inhibitors | Source | QSI Sensitive Microorganism | Effective Concentration of QS Inhibition | Action | Application | References |
|---|---|---|---|---|---|---|---|
| Higher plants | echinatin | Glycyrrhiza L. | Escherichia coli O157:H7 | 50 µM | Inhibition of biofilm formation. Inhibition of EPS 1 production. Inhibition of bacterial motility. Reduction in the expression of QS-regulated genes (luxS, pfs, lsrB, lsrK, lsrR, flhC, flhD, fliC, csgD, and stx2). | Antimicrobial agents against antibiotic-resistant E. coli; potential for treating E. coli infection. | [66] |
| carvacrol and eugenol | Carvacol from aromatic plants, thyme and oregano. Eugenol from cinnamon and clove oils | P. carotovorum subsp. brasiliense Pcb1692 P. aroidearum PC1 | 250 μM | Reduction of biofilm formation. Inhibit secretion of PCWDEs 2 (i.e., pectate lyase (Pel), polygalacturonase (Peh), and protease (Prt)). Inhibition of AHL production, potentially via direct interaction with ExpI/ExpR proteins. Downregulation of QS-regulated genes (rsmA, acrD and nssA). | Potential for soft rot disease control. | [67] | |
| phloretin | apple | Pectobacterium brasiliense | 200 µM | Reduction of biofilm formation. Reduction of bacterial motility. Reduction of the secretion of plant cell wall-degrading enzymes. Reduction in AHLs 3 production. Inhibition of expi activity. Downregulation of QS-regulated genes (expI, expR, luxS, rsmB), plant cell wall-degrading enzymes genes (pel, peh and prt) and motility genes (motA, fim, fliA, flhC and flhD). | Potential for plant-pathogenic bacteria control. | [68] | |
| Marine organisms | meleagrin | Penicillum chrysogenium | Chromobacterium violaceum | 138.42 µM | Inhibition of bacterial enoyl-acyl carrier protein reductase (FabI). | Antimicrobial agents against antibiotic-resistant human pathogens; potential for treating pathogenic infection. | [69,70] |
| alginate oligomer (OligoG CF-5/20) | Laminaria hyperborea | Pseudomonas aeruginosa | 2% | Inhibition of biofilm formation. Inhibition of bacterial motility. Reduction in AHLs’ production. Alteration in the extracellular production of the pseudomonal virulence factors pyo- cyanin, rhamnolipids, elastase, and total protease. Reduction in the expression of both the las and rhl systems. | Control chronic infections and biofilm-associated problems of P. aeruginosa. | [71,72] | |
| N-benzyl cinnamamide | Gracilaria fisheri | Vibrio harveyi | 1.66 mg/mL | Inhibition of biofilm formation. RReduction in bioluminescence via inhibition of AI-2 signaling. | Potential antimicrobial drug against V. harveyi. | [73] | |
| Bacteria | Amicoumacins | TRM B-02 Taklimakan desert bacterium | Chromobacterium violaceum | 31.25 µg/mL | Inhibition of the violacein biosynthetic pathway via downregulation of the expression of violacein operon A (vioA), vioB, vioD and vioE and upregulation of the expression of violacein operon C vioC, competitively inhibiting the binding of FAD 4 with the vioD enzyme. | Antimicrobial agents against antibiotic-resistant human pathogens; potential for treating pathogenic infection. | [74] |
| Fatty acyl compounds | Streptomyces griseoincarnatus HK12 | Pseudomonas aeruginosa Staphylococcus aureus | 100 μg/mL | Binding to the conserved sites of substrate binding in the quorum sensing system, LasI. | Antimicrobial agents against crucial nosocomial respiratory pathogen. | [75] | |
| Cyclic dipeptides (CDPs) | Pseudomonas aeruginosa RKC | Lelliottia amnigena RCE | 10 mg/mL | Regulation diverse metabolites of the pathogen diketopiperazine. Inhibition of QS-mediated pathogenicity via competitive binding with receptors | Potential for soft rot disease control. | [76] | |
| Synthetic compounds | synthetic peptides (LIVRHK and LIVRRK) | Pseudomonas aeruginosa PAO1 | 100 μg/mL | Inhibition of biofilm formation. Inhibits the production of virulence factors, including pyocyanin, protease, and rhamnolipids. downregulation of the expression of genes lasI, lasR, rhlI, and rhlR. | Control chronic infections and biofilm-associated problems of P. aeruginosa. | [77] | |
| N-acyl-2-aminopyrimidine derivatives | Acinetobacter baumannii | 3.8 μM | Inhibition of biofilm formation. Reduction in EPS production. Reduction of bacterial motility. | Antimicrobial agents against antibiotic-resistant human pathogens; potential for treating pathogenic infection. | [78] | ||
| PQIs (phc quorum sensing inhibitors) | Ralstonia solanacearum OE1-1 | 41.2 nM– 731 μM depends on (R)- or (S)- enantiomers | Act as competitive antagonists of 3-OH MAME 5. Inhibition of QS-dependent gene expression; repression inhibition of the production of ralfuranone and EPS. | Potential for plant-pathogenic bacteria control. | [79] |
| Autoinducer | Plant | Impact Effects | References |
|---|---|---|---|
| C6-HSL 3O-C6-HSL 3O-C8-HSL | Arabidopsis thaliana | Main root growth stimulation. | [115,116,117,118,119] |
| 3O-C10-HSL | Vigna radiata | Adventitious roots growth stimulation. | [17] |
| C4-HSL C6-HSL 3OHC4-HSL 3OHC6-HSL | Phaseolus L. Solanum lycopersicum | Systemic resilience similar to ISR. | [120] |
| C10-HSL | Arabidopsis thaliana | Inhibition of main root growth. Adventitious roots growth stimulation. Root hairs growth stimulation. | [121] |
| 3O-C8-HSL 3O-C14-HSL | Arabidopsis thaliana | Increased resistance to Pseudomonas syringae pv. tomato DC3000. | [16,18] |
| 3O-C14-HSL | Medicago truncatula | Increased root nodulation. | [122] |
| C4-HSL C6-HSL | Solanum lycopersicum | Increasing the content of salicylic acid. PR1a induction. Chitinase induction. | [123] |
| C6-HSL C8-HSL | Arabidopsis thaliana | Root growth stimulation by GCR1/GPA1 genes. | [117] |
| 3-okso-C14-HSL | Arabidopsis thaliana | Stimulating the expression of antioxidant and defense genes through the oxylipin and salicylic acid pathway. | [124] |
| C8-HSL C10-HSL | Hordeum L. Triticum L. | Root stimulation. Increasing the production of phase II antioxidant and detoxifying enzymes. | [125,126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majdura, J.; Jankiewicz, U.; Gałązka, A.; Orzechowski, S. The Role of Quorum Sensing Molecules in Bacterial–Plant Interactions. Metabolites 2023, 13, 114. https://doi.org/10.3390/metabo13010114
Majdura J, Jankiewicz U, Gałązka A, Orzechowski S. The Role of Quorum Sensing Molecules in Bacterial–Plant Interactions. Metabolites. 2023; 13(1):114. https://doi.org/10.3390/metabo13010114
Chicago/Turabian StyleMajdura, Jan, Urszula Jankiewicz, Agnieszka Gałązka, and Sławomir Orzechowski. 2023. "The Role of Quorum Sensing Molecules in Bacterial–Plant Interactions" Metabolites 13, no. 1: 114. https://doi.org/10.3390/metabo13010114
APA StyleMajdura, J., Jankiewicz, U., Gałązka, A., & Orzechowski, S. (2023). The Role of Quorum Sensing Molecules in Bacterial–Plant Interactions. Metabolites, 13(1), 114. https://doi.org/10.3390/metabo13010114