
Metabolomic Analysis of Wooden Breast Myopathy Shows a Disturbed Lipid Metabolism

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Sample Analysis
2.3. Data Analysis
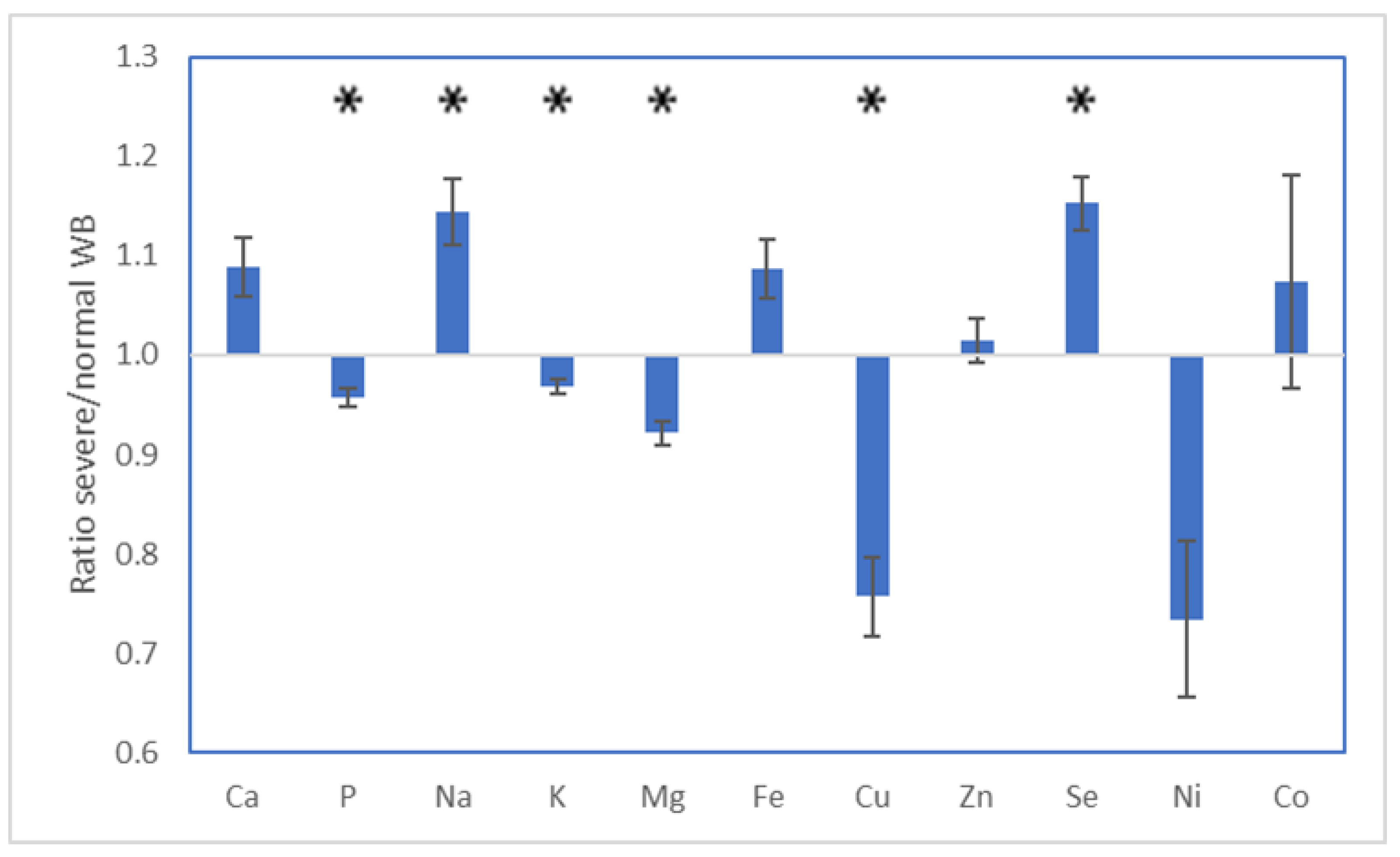
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petracci, M.; Mudalal, S.; Bonfiglio, A.; Cavani, C. Occurrence of white striping under commercial conditions and its impact on breast meat quality in broiler chickens. Poult. Sci. 2013, 92, 1670–1675. [Google Scholar] [CrossRef] [PubMed]
- Petracci, M.; Mudalal, S.; Soglia, F.; Cavani, C. Meat quality in fast-growing broiler chickens. World’s Poult. Sci. J. 2015, 71, 363–374. [Google Scholar] [CrossRef]
- Petracci, M.; Soglia, F.; Madruga, M.; Carvalho, L.; Ida, E.; Estévez, M. Wooden-breast, white striping, and spaghetti meat: Causes, consequences and consumer perception of emerging broiler meat abnormalities. Compr. Rev. Food Sci. Food Saf. 2019, 18, 565–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roma, R.; Corrado, S.; De Boni, A.; Forleo, M.B.; Fantin, V.; Moretti, M.; Palmieri, N.; Vitali, A.; Camillo, D.C. Life cycle assessment in the livestock and derived edible products sector. In Life Cycle Assessment in the Agri-Food Sector; Springer: Berlin/Heidelberg, Germany, 2015; pp. 251–332. [Google Scholar]
- Costantini, M.; Ferrante, V.; Guarino, M.; Bacenetti, J. Environmental sustainability assessment of poultry productions through life cycle approaches: A critical review. Trends Food Sci. Technol. 2021, 110, 201–212. [Google Scholar] [CrossRef]
- Tixier-Boichard, M. From the jungle fowl to highly performing chickens: Are we reaching limits? World’s Poult. Sci. J. 2020, 76, 2–17. [Google Scholar] [CrossRef]
- Alnahhas, N.; Berri, C.; Chabault, M.; Chartrin, P.; Boulay, M.; Bourin, M.C.; Bihan-Duval, L. Genetic parameters of white striping in relation to body weight, carcass composition, and meat quality traits in two broiler lines divergently selected for the ultimate pH of the pectoralis major muscle. BMC Genet. 2016, 17, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.R.; Suyemoto, M.M.; Sarsour, A.H.; Cordova, H.A.; Oviedo-Rondón, E.O.; Wineland, M.; Barnes, H.J.; Borst, L.B. Temporal characterization of wooden breast myopathy (“woody breast”) severity and correlation with growth rate and lymphocytic phlebitis in three commercial broiler strains and a random-bred broiler strain. Avian Pathol. 2019, 48, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Soglia, F.; Petracci, M.; Davoli, R.; Zappaterra, M. A critical review of the mechanisms involved in the occurrence of growth-related abnormalities affecting broiler chicken breast muscles. Poult. Sci. 2021, 100, 101180. [Google Scholar] [CrossRef] [PubMed]
- Geronimo, B.C.; Prudencio, S.H.; Soares, A.L. Biochemical and technological characteristics of wooden breast chicken fillets and their consumer acceptance. J. Food Sci. Technol. 2022, 59, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, P.; Beitia, A.; Weil, J.; Suesuttajit, N.; Hilton, K.; Caldas, J.; Umberson, C.; Martinez, D.; Kong, B.; Owens, C.M. Woody breast myopathy broiler show age-dependent adaptive differential gene expression in Pectoralis major and altered in-vivo triglyceride kinetics in adipogenic tissues. Poult. Sci. 2021, 100, 101092. [Google Scholar] [CrossRef]
- Petracci, M.; Mudalal, S.; Babini, E.; Cavani, C. Effect of white striping on chemical composition and nutritional value of chicken breast meat. Ital. J. Anim. Sci. 2014, 13, 3138. [Google Scholar] [CrossRef]
- Kuttappan, V.; Hargis, B.; Owens, C. White striping and woody breast myopathies in the modern poultry industry: A review. Poult. Sci. 2016, 95, 2724–2733. [Google Scholar] [CrossRef] [PubMed]
- Kuttappan, V.A.; Lee, Y.S.; Erf, G.F.; Meullenet, J.F.; McKee, S.R.; Owens, C.M. Consumer acceptance of visual appearance of broiler breast meat with varying degrees of white striping. Poult. Sci. 2012, 91, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Boerboom, G.; van Kempen, T.; Navarro-Villa, A.; Pérez-Bonilla, A. Unraveling the cause of white striping in broilers using metabolomics. Poult. Sci. 2018, 97, 3977–3986. [Google Scholar] [CrossRef]
- Muroya, S.; Ueda, S.; Komatsu, T.; Miyakawa, T.; Ertbjerg, P. MEATabolomics: Muscle and meat metabolomics in domestic animals. Metabolites 2020, 10, 188. [Google Scholar] [CrossRef]
- Sihvo, H.-K.; Lindén, J.; Airas, N.; Immonen, K.; Valaja, J.; Puolanne, E. Wooden breast myodegeneration of pectoralis major muscle over the growth period in broilers. Veter Pathol. 2017, 54, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sihvo, H.K.; Immonen, K.; Puolanne, E. Myodegeneration with fibrosis and regeneration in the pectoralis major muscle of broilers. Veter Pathol. 2013, 51, 619–623. [Google Scholar] [CrossRef]
- Wold, J.P.; Veiseth-Kent, E.; Høst, V.; Løvland, A. Rapid on-line detection and grading of wooden breast myopathy in chicken fillets by near-infrared spectroscopy. PLoS ONE 2017, 12, e0173384. [Google Scholar] [CrossRef] [Green Version]
- Soglia, F.; Laghi, L.; Canonico, L.; Cavani, C.; Petracci, M. Functional property issues in broiler breast meat related to emerging muscle abnormalities. Food Res. Int. 2016, 89, 1071–1076. [Google Scholar] [CrossRef]
- Livingston, M.L.; Landon, C.; Barnes, H.; Brake, J. White striping and wooden breast myopathies of broiler breast muscle is affected by time-limited feeding, genetic background, and egg storage. Poult. Sci. 2019, 98, 217–226. [Google Scholar] [CrossRef]
- Brothers, B.; Zhuo, Z.; Papah, M.B.; Abasht, B. RNA-seq analysis reveals spatial and sex differences in pectoralis major muscle of broiler chickens contributing to difference in susceptibility to wooden breast disease. Front. Physiol. 2019, 10, 764. [Google Scholar] [CrossRef] [PubMed]
- Papah, M.B.; Abasht, B. Dysregulation of lipid metabolism and appearance of slow myofiber-specific isoforms accompany the development of Wooden Breast myopathy in modern broiler chickens. Sci. Rep. 2019, 9, 17170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tijare, V.V.; Yang, F.L.; Kuttappan, V.A.; Alvarado, C.Z.; Coon, C.N.; Owens, C.M. Meat quality of broiler breast fillets with white striping and woody breast muscle myopathies. Poult. Sci. 2016, 95, 2167–2173. [Google Scholar] [CrossRef] [PubMed]
- Bikker, P.; ten Tije, N.; Tijkorte, A. Fosforbenutting Bij Biologisch Gehouden Vleesvarkens; Wageningen Livestock Research: Wageningen, The Netherlands, 2017. [Google Scholar]
- van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genomic. 2006, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Mandal, R.; Sinelnikov, I.V.; Broadhurst, D.; Wishart, D.S. MetaboAnalyst 2.0—A comprehensive server for metabolomic data analysis. Nucleic Acids Res. 2012, 40, W127–W133. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wishart, D.S. Metabolomic data processing, analysis, and interpretation using MetaboAnalyst. Curr. Protoc. Bioinform. 2011, 34, 14.10.1–14.10.48. [Google Scholar] [CrossRef]
- Vinaixa, M.; Samino, S.; Saez, I.; Duran, J.; Guinovart, J.J.; Yanes, O. A Guideline to Univariate Statistical Analysis for LC/MS-Based Untargeted Metabolomics-Derived Data. Metabolites 2012, 2, 775. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Zampiga, M.; Soglia, F.; Petracci, M.; Meluzzi, A.; Sirri, F. Effect of different arginine-to-lysine ratios in broiler chicken diets on the occurrence of breast myopathies and meat quality attributes. Poult. Sci. 2019, 98, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Lake, J.A.; Yan, Y.; Dekkers, J.C.; Qiu, J.; Brannick, E.M.; Abasht, B. Identification of circulating metabolites associated with wooden breast and white striping. PLoS ONE 2022, 17, e0274208. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, R.J.; Farney, T.M.; Trepanowski, J.F.; McCarthy, C.G.; Canale, R.E.; Schilling, B.K. Comparison of pre-workout nitric oxide stimulating dietary supplements on skeletal muscle oxygen saturation, blood nitrate/nitrite, lipid peroxidation, and upper body exercise performance in resistance trained men. J. Int. Soc. Sports Nutr. 2010, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Sun, W.-D.; Li, J.-C.; Pan, J.-Q.; Liu, Y.-J.; Wang, J.-Y.; Wang, X.-L. L-arginine prevents reduced expression of endothelial nitric oxide synthase (NOS) in pulmonary arterioles of broilers exposed to cool temperatures. Veter J. 2007, 173, 151–157. [Google Scholar] [CrossRef]
- Guo, Y.; Shi, B.; Yan, S.; Xu, Y.; Li, J.; Li, T. Effects of arginine on cytokines and nitric oxide synthesis in broilers. J. Anim. Plant Sci. 2015, 25, 366–371. [Google Scholar]
- Haug, A.; R⊘dbotten, R.; Mydland, L.T.; Christophersen, O.A. Increased broiler muscle carnosine and anserine following histidine supplementation of commercial broiler feed concentrate. Acta Agric. 2008, 58, 71–77. [Google Scholar] [CrossRef]
- Kai, S.; Watanabe, G.; Kubota, M.; Kadowaki, M.; Fujimura, S. Effect of dietary histidine on contents of carnosine and anserine in muscles of broilers. Anim. Sci. J. 2015, 86, 541–546. [Google Scholar] [CrossRef]
- Kim, S.-K.; Kim, Y.-M.; Baek, I.-K.; Auh, J.-H. Carnosine and anserine in chicken: Distribution, age-dependency and their anti-glycation activity. J. Food Sci. Anim. Resour. 2012, 32, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Abasht, B.; Mutryn, M.F.; Michalek, R.D.; Lee, W.R. Oxidative stress and metabolic perturbations in wooden breast disorder in chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Zhang, K.; Tian, G.; Bai, S.; Ding, X.; Wang, J.; Peng, H.; Lv, L.; Xuan, Y.; Zeng, Q. Effects of dietary corn germ meal levels on growth performance, serum biochemical parameters, meat quality, and standardized ileal digestibility of amino acids in Pekin ducks. Poult. Sci. 2022, 101, 101779. [Google Scholar] [CrossRef]
- Calzada, E.; Onguka, O.; Claypool, S.M. Phosphatidylethanolamine metabolism in health and disease. Int. Rev. Cell Mol. Biol. 2016, 321, 29–88. [Google Scholar] [CrossRef] [PubMed]
- Böttinger, L.; Horvath, S.E.; Kleinschroth, T.; Hunte, C.; Daum, G.; Pfanner, N.; Becker, T. Phosphatidylethanolamine and cardiolipin differentially affect the stability of mitochondrial respiratory chain supercomplexes. J. Mol. Biol. 2012, 423, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhof, I.; Canivenc-Gansel, E.; Meyer, U.; Conzelmann, A. Phosphatidylethanolamine is the donor of the phosphorylethanolamine linked to the α1, 4-linked mannose of yeast GPI structures. Glycobiology 2000, 10, 1271–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Agellon, L.B.; Allen, T.M.; Umeda, M.; Jewell, L.; Mason, A.; Vance, D.E. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis. Cell Metab. 2006, 3, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez, L.O.; Leon, M.; Einav, S.; Varon, J. Beyond muscle destruction: A systematic review of rhabdomyolysis for clinical practice. Crit. Care 2016, 20, 135. [Google Scholar] [CrossRef] [Green Version]
- Romero-Aristizabal, C.; Marks, D.S.; Fontana, W.; Apfeld, J. Regulated spatial organization and sensitivity of cytosolic protein oxidation in Caenorhabditis elegans. Nat. Commun. 2014, 5, 5020. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, N.; Nielsen, T.K.; Bjørn, S.P.; Axelsson, I.; Paglia, G.; Voldborg, B.G.; Palsson, B.O.; Rolfsson, Ó. Biochemical characterization of human gluconokinase and the proposed metabolic impact of gluconic acid as determined by constraint based metabolic network analysis. PLoS ONE 2014, 9, e98760. [Google Scholar] [CrossRef] [Green Version]
- De Koning, T.J.; Snell, K.; Duran, M.; Berger, R.; Poll-The, B.-T.; Surtees, R. L-serine in disease and development. Biochem. J. 2003, 371, 653–661. [Google Scholar] [CrossRef]
- Mishra, R.C.; Tripathy, S.; Quest, D.; Desai, K.M.; Akhtar, J.; Dattani, I.D.; Gopalakrishnan, V. L-Serine lowers while glycine increases blood pressure in chronic L-NAME-treated and spontaneously hypertensive rats. Am. J. Hypertens. 2008, 26, 2339–2348. [Google Scholar] [CrossRef]
- Conte Camerino, D.; Tricarico, D.; Pierno, S.; Desaphy, J.F.; Liantonio, A.; Pusch, M.; Burdi, R.; Camerino, C.; Fraysse, B.; De Luca, A. Taurine and skeletal muscle disorders. Neurochem. Res. 2004, 29, 135–142. [Google Scholar] [CrossRef]
- Lambert, I.H. Regulation of the cellular content of the organic osmolyte taurine in mammalian cells. Neurochem. Res. 2004, 29, 27–63. [Google Scholar] [CrossRef] [PubMed]
- Toue, S.; Kodama, R.; Amao, M.; Kawamata, Y.; Kimura, T.; Sakai, R. Screening of toxicity biomarkers for methionine excess in rats. J. Nutr. 2006, 136, 1716S–1721S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogdahl, Å. Digestion and absorption of lipids in poultry. J. Nutr. 1985, 115, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Nir, I.; Nitsan, Z.; Vax, A. The influence of force feeding and of protein supplementation to the diet on the metabolisable energy of diets, digestibility of nutrients, nitrogen retention and digestive enzymes output in geese. Ann. Biol. Anim. Biochim. Biophys. 1973, 13, 465–479. [Google Scholar] [CrossRef] [Green Version]
- Abasht, B.; Papah, M.B.; Qiu, J. Evidence of vascular endothelial dysfunction in Wooden Breast disorder in chickens: Insights through gene expression analysis, ultra-structural evaluation and supervised machine learning methods. PLoS ONE 2021, 16, e0243983. [Google Scholar] [CrossRef]
- Liu, R.; Kong, F.; Xing, S.; He, Z.; Bai, L.; Sun, J.; Tan, X.; Zhao, D.; Zhao, G.; Wen, J. Dominant changes in the breast muscle lipid profiles of broiler chickens with wooden breast syndrome revealed by lipidomics analyses. J. Anim. Sci. Biotechnol. 2022, 13, 93. [Google Scholar] [CrossRef]
- van Kempen, T.A.; Reijersen, M.; de Bruijn, C.; De Smet, S.; Michiels, J.; Traber, M.G.; Lauridsen, C. Vitamin E plasma kinetics in swine show low bioavailability and short half-life of all-rac-α-tocopheryl acetate. J. Anim. Sci. 2016, 94, 4188–4195. [Google Scholar] [CrossRef]
- Thompson, G.R. Lipid related consequences of intestinal malabsorption. Gut 1989, 30, 29–34. [Google Scholar] [CrossRef]
- Mutryn, M.F.; Brannick, E.M.; Fu, W.; Lee, W.R.; Abasht, B. Characterization of a novel chicken muscle disorder through differential gene expression and pathway analysis using RNA-sequencing. BMC Genom. 2015, 16, 399. [Google Scholar] [CrossRef] [Green Version]
- Mandatori, D.; Pelusi, L.; Schiavone, V.; Pipino, C.; Di Pietro, N.; Pandolfi, A. The dual role of vitamin K2 in “bone-vascular crosstalk”: Opposite effects on bone loss and vascular calcification. Nutrients 2021, 13, 1222. [Google Scholar] [CrossRef]
- Kong, F.; Zhao, G.; He, Z.; Sun, J.; Wang, X.; Liu, D.; Zhu, D.; Liu, R.; Wen, J. Serum Creatine Kinase as a Biomarker to Predict Wooden Breast in vivo for Chicken Breeding. Front. Physiol. 2021, 12, 711711. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Torres, A.G.; Ribas de Pouplana, L. Inosine in biology and disease. Genes 2021, 12, 600. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, V.; Van Der Giet, M.; Mischak, H.; Morgan, M.; Zidek, W.; Jankowski, J. Dinucleoside polyphosphates: Strong endogenous agonists of the purinergic system. Br. J. Pharmacol. 2009, 157, 1142–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dong, X.; Puolanne, E.; Ertbjerg, P. Effect of wooden breast degree on lipid and protein oxidation and citrate synthase activity of chicken pectoralis major muscle. LWT 2022, 154, 112884. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient Supplementation (%) | Starter Phase (d0–7) | Grower Phase (d7–25) | Finisher Phase (d25–35) |
|---|---|---|---|
| Wheat | 52.63 | 56.00 | 61.70 |
| Soybean meal 47 | 28.77 | 25.89 | 21.51 |
| Maize | 10.00 | 8.81 | 8.00 |
| Soy oil | 4.41 | 4.25 | 4.37 |
| Glycerin 99.5% | 2.00 | 2.00 | |
| Soycomil | 0.20 | ||
| Monocalcium phosphate | 1.24 | 0.63 | 0.32 |
| Calcium carbonate, | 1.17 | 0.54 | 0.45 |
| Sodium bicarbonate | 0.28 | 0.22 | 0.24 |
| NaCl | 0.15 | 0.16 | 0.16 |
| L-lysine∙HCl | 0.30 | 0.27 | 0.29 |
| DL-methionine | 0.23 | 0.21 | 0.19 |
| L-threonine | 0.066 | 0.070 | 0.076 |
| Xylanase (Axtra XB) | 0.10 | 0.10 | 0.10 |
| Phytase (Phyzyme XP TPT) | 0.10 | 0.10 | 0.10 |
| Maxiban 16% | 0.063 | ||
| Elancoban 20% | 0.050 | ||
| Premix 1 | 0.50 | 0.50 | 0.50 |
| Calculated Nutrient values | |||
| Crude protein | 21.7 | 20.4 | 19.4 |
| AME broilers (kcal/kg) | 2850 | 2922 | 3000 |
| dLys | 11.5 | 10.60 | 9.80 |
| dMet | 5.03 | 4.69 | 4.30 |
| Ca | 9.0 | 5.50 | 4.5 |
| dP | 4.6 | 3.50 | 2.90 |
| WS | |||||
|---|---|---|---|---|---|
| Normal | Moderate | Severe | WB Total | ||
| WB | Normal | 15 | 15 | 7 | 37 |
| Severe | 4 | 9 | 1 | 14 | |
| WS total | 19 | 24 | 8 | 51 | |
| Pathway Name | p-Value (FDR Corrected) | Impact |
|---|---|---|
| Histidine metabolism | 0.002106 | 0.54917 |
| Starch and sucrose metabolism | 0.019798 | 0.22073 |
| Beta-alanine metabolism | 0.019798 | 0.51119 |
| Taurine and hypotaurine metabolism | 0.023189 | 0.71428 |
| Arginine and proline metabolism | 0.026478 | 0.48672 |
| Glutathione metabolism | 0.038001 | 0.37762 |
| Glycerophospholipid metabolism | 0.042407 | 0.43635 |
| Cysteine and methionine metabolism | 0.042453 | 0.44302 |
| D-Glutamine and D-glutamate metabolism | 0.043772 | 0.50000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boerboom, G.M.; Navarro-Villa, A.; van Kempen, T.A.T.G. Metabolomic Analysis of Wooden Breast Myopathy Shows a Disturbed Lipid Metabolism. Metabolites 2023, 13, 20. https://doi.org/10.3390/metabo13010020
Boerboom GM, Navarro-Villa A, van Kempen TATG. Metabolomic Analysis of Wooden Breast Myopathy Shows a Disturbed Lipid Metabolism. Metabolites. 2023; 13(1):20. https://doi.org/10.3390/metabo13010020
Chicago/Turabian StyleBoerboom, Gavin M., Alberto Navarro-Villa, and Theo A. T. G. van Kempen. 2023. "Metabolomic Analysis of Wooden Breast Myopathy Shows a Disturbed Lipid Metabolism" Metabolites 13, no. 1: 20. https://doi.org/10.3390/metabo13010020
APA StyleBoerboom, G. M., Navarro-Villa, A., & van Kempen, T. A. T. G. (2023). Metabolomic Analysis of Wooden Breast Myopathy Shows a Disturbed Lipid Metabolism. Metabolites, 13(1), 20. https://doi.org/10.3390/metabo13010020