
Positive Correlation of Triacylglycerols with Increased Chain Length and Unsaturation with ω-O-Acylceramide and Ceramide-NP as Well as Acidic pH in the Skin Surface of Healthy Korean Adults

 , and
, and 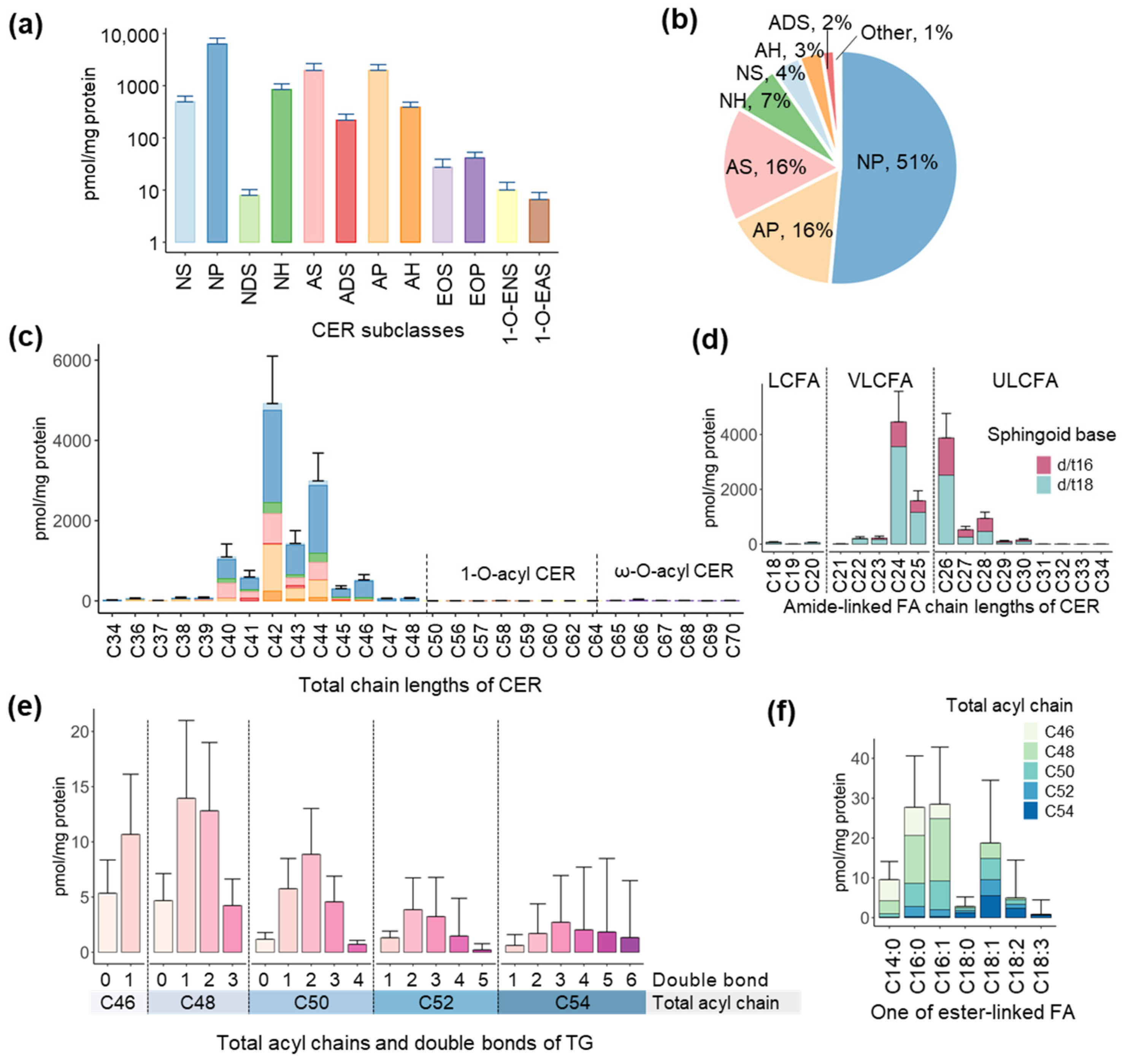{kind=link}
{kind=link}
{kind=link}
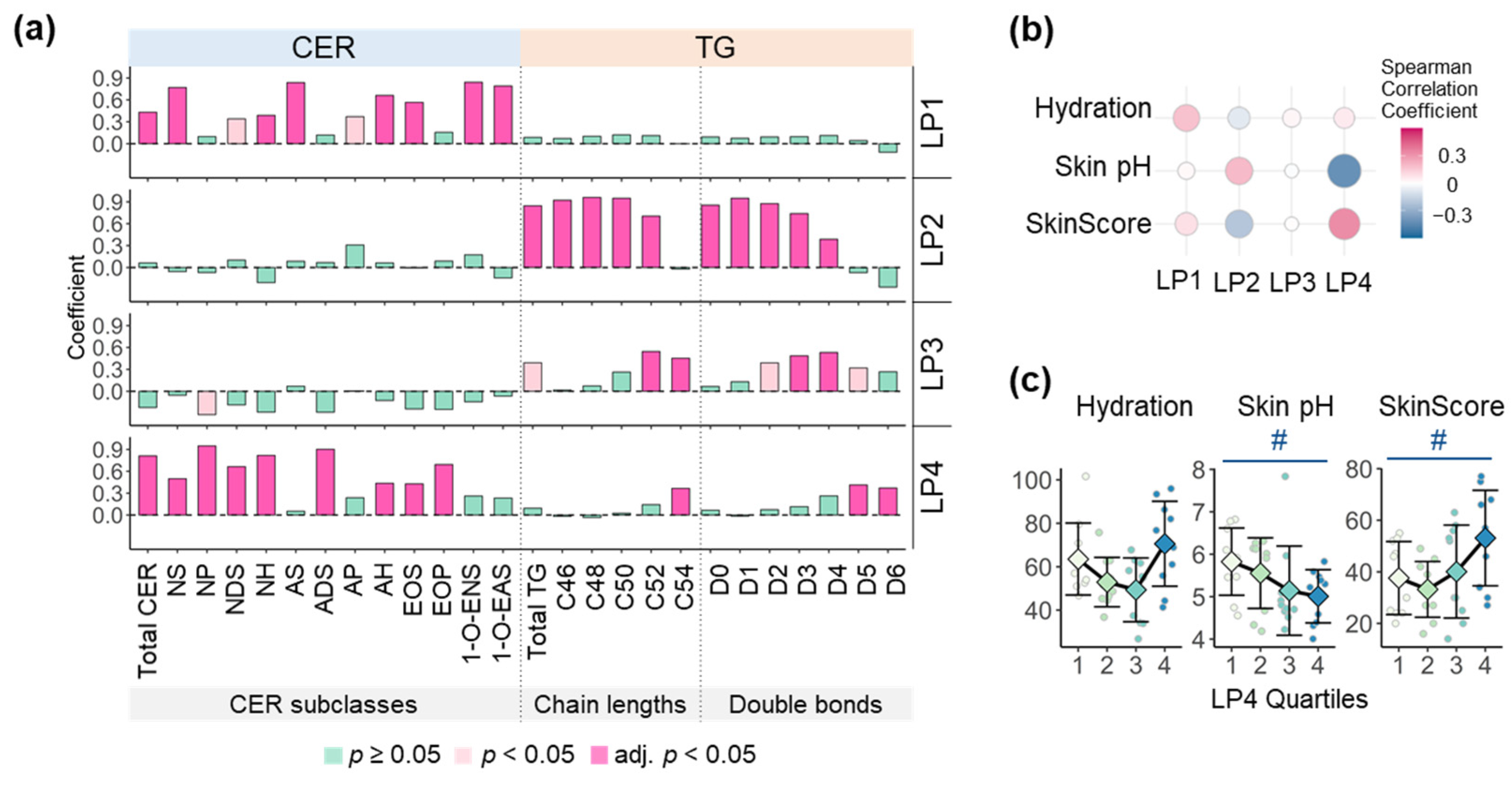{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants and Measurement of Skin Health Parameters
2.2. Lipidomic Analysis
2.3. Statistical Analysis and Bioinformatics
3. Results
3.1. Profiles of CER and TG in the Skin Surface
3.2. Associations of TG-FA with Ester-Linked FAs of acylCER
3.3. Associations of TG with Overall CER Subclasses
3.4. Skin Surface Lipids and Skin Health Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | atopic dermatitis |
| ANOVA | analysis of variance |
| BH | Benjamini & Hochberg |
| CER | ceramide |
| CLE | corneocyte lipid envelope |
| DGAT2 | diacylglycerol acyltransferase2 |
| FA | fatty acid |
| FFA | free fatty acid |
| GLA | γ-linolenic acid |
| HPTLC | high performance thin layer chromatography |
| LA | linoleic acid |
| LCFA | long-chain fatty acid |
| LP | Lipid pattern |
| LPLA2 | lysosomal phospholipase A2 |
| PUFA | polyunsaturated fatty acid |
| SB | sphingoid base |
| SC | stratum corneum |
| TG | triacylglycerol |
| ULCFA | ultra long-chain fatty acid |
| VLCFA | very long-chain fatty acid |
References
- Elias, P.M.; Fartasch, M.; Crumrine, D.; Behne, M.; Uchida, Y.; Holleran, W.M. Origin of the Corneocyte Lipid Envelope (CLE): Observations in Harlequin Ichthyosis and Cultured Human Keratinocytes. J. Investig. Dermatol. 2000, 115, 765–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, P.M. Epidermal Lipids, Barrier Function, and Desquamation. J. Investig. Dermatol. 1983, 80, S44–S49. [Google Scholar] [CrossRef] [PubMed]
- Boer, M.; Duchnik, E.; Maleszka, R.; Marchlewicz, M. Structural and Biophysical Characteristics of Human Skin in Maintaining Proper Epidermal Barrier Function. Adv. Dermatol. Allergol. 2016, 33, 1–5. [Google Scholar] [CrossRef]
- Fluhr, J.W.; Kao, J.; Jain, M.; Ahn, S.K.; Feingold, K.R.; Elias, P.M. Generation of Free Fatty Acids from Phospholipids Regulates Stratum Corneum Acidification and Integrity. J. Investig. Dermatol. 2001, 117, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meckfessel, M.H.; Brandt, S. The Structure, Function, and Importance of Ceramides in Skin and Their Use as Therapeutic Agents in Skin-Care Products. J. Am. Acad. Dermatol. 2014, 71, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Breiden, B.; Sandhoff, K. The Role of Sphingolipid Metabolism in Cutaneous Permeability Barrier Formation. Biochim. Biophys. Acta 2014, 1841, 441–452. [Google Scholar] [CrossRef]
- Motta, S.; Monti, M.; Sesana, S.; Caputo, R.; Carelli, S.; Ghidoni, R. Ceramide Composition of the Psoriatic Scale. Biochim. Biophys. Acta 1993, 1182, 147–151. [Google Scholar] [CrossRef]
- Rabionet, M.; Gorgas, K.; Sandhoff, R. Ceramide Synthesis in the Epidermis. Biochim. Biophys. Acta 2014, 1841, 422–434. [Google Scholar] [CrossRef]
- Nadaban, A.; Gooris, G.S.; Beddoes, C.M.; Dalgliesh, R.M.; Bouwstra, J.A. Phytosphingosine Ceramide Mainly Localizes in the Central Layer of the Unique Lamellar Phase of Skin Lipid Model Systems. J. Lipid Res. 2022, 63, 100258. [Google Scholar] [CrossRef]
- Di Nardo, A.; Wertz, P.; Giannetti, A.; Seidenari, S. Ceramide and Cholesterol Composition of the Skin of Patients with Atopic Dermatitis. Acta Derm. Venereol. 1998, 78, 27–30. [Google Scholar] [CrossRef]
- Bleck, O.; Abeck, D.; Ring, J.; Hoppe, U.; Vietzke, J.P.; Wolber, R.; Brandt, O.; Schreiner, V. Two Ceramide Subfractions Detectable in Cer(AS) Position by HPTLC in Skin Surface Lipids of Non-Lesional Skin of Atopic Eczema. J. Investig. Dermatol. 1999, 113, 894–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knox, S.; O’Boyle, N.M. Skin Lipids in Health and Disease: A Review. Chem. Phys. Lipids 2021, 236, 105055. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Holleran, W.M. Omega-O-Acylceramide, a Lipid Essential for Mammalian Survival. J. Dermatol. Sci. 2008, 51, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased Level of Ceramides in Stratum Corneum of Atopic Dermatitis: An Etiologic Factor in Atopic Dry Skin? J. Investig. Dermatol. 1991, 96, 523–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, J.; Narita, H.; Kondo, N.; Hotta, M.; Takagi, Y.; Masukawa, Y.; Kitahara, T.; Takema, Y.; Koyano, S.; Yamazaki, S.; et al. Changes in the Ceramide Profile of Atopic Dermatitis Patients. J. Investig. Dermatol. 2010, 130, 2511–2514. [Google Scholar] [CrossRef] [Green Version]
- Rabionet, M.; Bayerle, A.; Marsching, C.; Jennemann, R.; Gröne, H.J.; Yildiz, Y.; Wachten, D.; Shaw, W.; Shayman, J.A.; Sandhoff, R. 1-O-Acylceramides Are Natural Components of Human and Mouse Epidermis. J. Lipid Res. 2013, 54, 3312–3321. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.J.; Feingold, K.R. The Expression and Regulation of Enzymes Mediating the Biosynthesis of Triglycerides and Phospholipids in Keratinocytes/Epidermis. Dermato-Endocrinology 2011, 3, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Radner, F.P.W.; Fischer, J. The Important Role of Epidermal Triacylglycerol Metabolism for Maintenance of the Skin Permeability Barrier Function. Biochim. Biophys. Acta 2014, 1841, 409–415. [Google Scholar] [CrossRef]
- Ujihara, M.; Nakajima, K.; Yamamoto, M.; Teraishi, M.; Uchida, Y.; Akiyama, M.; Shimizu, H.; Sano, S. Epidermal Triglyceride Levels Are Correlated with Severity of Ichthyosis in Dorfman-Chanarin Syndrome. J. Dermatol. Sci. 2010, 57, 102–107. [Google Scholar] [CrossRef]
- Stone, S.J.; Myers, H.M.; Watkins, S.M.; Brown, B.E.; Feingold, K.R.; Elias, P.M.; Farese, R.V. Lipopenia and Skin Barrier Abnormalities in DGAT2-Deficient Mice. J. Biol. Chem. 2004, 279, 11767–11776. [Google Scholar] [CrossRef]
- Lee, J.Y.; Liu, K.H.; Cho, Y.; Kim, K.P. Enhanced Triacylglycerol Content and Gene Expression for Triacylglycerol Metabolism, Acyl-Ceramide Synthesis, and Corneocyte Lipid Formation in the Epidermis of Borage Oil Fed Guinea Pigs. Nutrients 2019, 11, 2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melton, J.L.; Wertz, P.W.; Swartzendruber, D.C.; Downing, D.T. Effects of Essential Fatty Acid Deficiency on Epidermal O-Acylsphingolipids and Transepidermal Water Loss in Young Pigs. Biochim. Biophys. Acta 1987, 921, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Voynova, N.S.; Vionnet, C.; Ejsing, C.S.; Conzelmann, A. A Novel Pathway of Ceramide Metabolism in Saccharomyces Cerevisiae. Biochem. J. 2012, 447, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Lu, L.; Zhao, Z.; Cai, M.; Gao, N.; Han, G. Lipidomics Profiling of Skin Surface Lipids in Senile Pruritus. Lipids Health Dis. 2020, 19, 171. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, N.; Sato, W.J.; Kelly, A.; Ganguli-Indra, G.; Indra, A.K. Epidermal Lipids: Key Mediators of Atopic Dermatitis Pathogenesis. Trends Mol. Med. 2019, 25, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.K.; Shon, J.C.; Seo, H.S.; Liu, K.H.; Lee, J.W.; Ahn, S.K.; Hong, S.P. Decrease of Ceramides with Long-Chain Fatty Acids in Psoriasis: Possible Inhibitory Effect of Interferon Gamma on Chain Elongation. Exp. Dermatol. 2022, 31, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.J.; Lee, G.M.; Liu, K.H.; Jung, D.H. Effect of Korean Red Ginseng on Plasma Ceramide Levels in Postmenopausal Women with Hypercholesterolemia: A Pilot Randomized Controlled Trial. Metabolites 2021, 11, 417. [Google Scholar] [CrossRef]
- Cajka, T.; Fiehn, O. Comprehensive Analysis of Lipids in Biological Systems by Liquid Chromatography-Mass Spectrometry. Trends Anal. Chem. 2014, 61, 192–206. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, C.; Han, X. Selection of Internal Standards for Accurate Quantification of Complex Lipid Species in Biological Extracts by Electrospray Ionization Mass Spectrometry-What, How and Why? Mass Spectrom. Rev. 2017, 36, 693–714. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Ko, Y.; Park, Y.K.; Kim, N.I.; Ha, W.K.; Cho, Y. Dietary Effect of Lactoferrin-Enriched Fermented Milk on Skin Surface Lipid and Clinical Improvement of Acne Vulgaris. Nutrition 2010, 26, 902–909. [Google Scholar] [CrossRef]
- T’Kindt, R.; Jorge, L.; Dumont, E.; Couturon, P.; David, F.; Sandra, P.; Sandra, K. Profiling and Characterizing Skin Ceramides Using Reversed-Phase Liquid Chromatography-Quadrupole Time-of-Flight Mass Spectrometry. Anal. Chem. 2012, 84, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Kawana, M.; Miyamoto, M.; Ohno, Y.; Kihara, A. Comparative Profiling and Comprehensive Quantification of Stratum Corneum Ceramides in Humans and Mice by LC/MS/MS. J. Lipid Res. 2020, 61, 884–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, M.; Ohno, Y.; Kihara, A. Whole Picture of Human Stratum Corneum Ceramides, Including the Chain-Length Diversity of Long-Chain Bases. J. Lipid Res. 2022, 63, 100235. [Google Scholar] [CrossRef] [PubMed]
- Školová, B.; Januìššová, B.; Zbytovská, J.; Gooris, G.; Bouwstra, J.; Slepička, P.; Berka, P.; Roh, J.; Palát, K.; Hrabálek, A.; et al. Ceramides in the Skin Lipid Membranes: Length Matters. Langmuir 2013, 29, 15624–15633. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, T.; Klose, C.; Gerl, M.J.; Wójcik-Maciejewicz, A.; Herzog, R.; Simons, K.; Reich, A.; Surma, M.A. Large-Scale Human Skin Lipidomics by Quantitative, High-Throughput Shotgun Mass Spectrometry. Sci. Rep. 2017, 7, 43761. [Google Scholar] [CrossRef] [Green Version]
- Crowther, J.M. Method for Quantification of Oils and Sebum Levels on Skin Using the Sebumeter(®). Int. J. Cosmet. Sci. 2016, 38, 210–216. [Google Scholar] [CrossRef]
- Marzouki, Z.M.H.; Taha, A.M.; Gomaa, K.S. Fatty Acid Profiles of Sebaceous Triglycerides by Capillary Gas Chromatography with Mass-Selective Detection. J. Chromatogr. B Biomed. Sci. Appl. 1988, 425, 11–24. [Google Scholar] [CrossRef]
- Schafer, L.; Kragballe, K. Abnormalities in Epidermal Lipid Metabolism in Patients with Atopic Dermatitis. J. Investig. Dermatol. 1991, 96, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Janssens, M.; van Smeden, J.; Gooris, G.S.; Bras, W.; Portale, G.; Caspers, P.J.; Vreeken, R.J.; Hankemeier, T.; Kezic, S.; Wolterbeek, R.; et al. Increase in Short-Chain Ceramides Correlates with an Altered Lipid Organization and Decreased Barrier Function in Atopic Eczema Patients. J. Lipid Res. 2012, 53, 2755–2766. [Google Scholar] [CrossRef] [Green Version]
- Abe, A.; Shayman, J.A.; Radin, N.S. A Novel Enzyme That Catalyzes the Esterification of N-Acetylsphingosine. Metabolism of C2-Ceramides. J. Biol. Chem. 1996, 271, 14383–14389. [Google Scholar] [CrossRef]
- Shayman, J.A.; Kelly, R.; Kollmeyer, J.; He, Y.; Abe, A. Group XV Phospholipase A2, a Lysosomal Phospholipase A2. Prog. Lipid Res. 2011, 50, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Smeden, J.; Bouwstra, J.A. Stratum Corneum Lipids: Their Role for the Skin Barrier Function in Healthy Subjects and Atopic Dermatitis Patients. Ski. Barrier Funct. 2016, 49, 8–26. [Google Scholar] [CrossRef]
- Selander, C.; Zargari, A.; Möllby, R.; Rasool, O.; Scheynius, A. Higher PH Level, Corresponding to That on the Skin of Patients with Atopic Eczema, Stimulates the Release of Malassezia Sympodialis Allergens. Allergy 2006, 61, 1002–1008. [Google Scholar] [CrossRef] [PubMed]
- Eberlein-Konig, B.; Schafer, T.; Huss-Marp, J.; Darsow, U.; Mohrenschlager, M.; Herbert, O.; Abeck, D.; Kramer, U.; Behrendt, H.; Ring, J. Skin Surface PH, Stratum Corneum Hydration, Trans-Epidermal Water Loss and Skin Roughness Related to Atopic Eczema and Skin Dryness in a Population of Primary School Children. Acta Derm. Venereol. 2000, 80, 188–191. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-Y.; Jeon, S.; Han, S.; Liu, K.-H.; Cho, Y.; Kim, K.-P. Positive Correlation of Triacylglycerols with Increased Chain Length and Unsaturation with ω-O-Acylceramide and Ceramide-NP as Well as Acidic pH in the Skin Surface of Healthy Korean Adults. Metabolites 2023, 13, 31. https://doi.org/10.3390/metabo13010031
Lee J-Y, Jeon S, Han S, Liu K-H, Cho Y, Kim K-P. Positive Correlation of Triacylglycerols with Increased Chain Length and Unsaturation with ω-O-Acylceramide and Ceramide-NP as Well as Acidic pH in the Skin Surface of Healthy Korean Adults. Metabolites. 2023; 13(1):31. https://doi.org/10.3390/metabo13010031
Chicago/Turabian StyleLee, Ju-Young, Sanghun Jeon, Sangshin Han, Kwang-Hyeon Liu, Yunhi Cho, and Kun-Pyo Kim. 2023. "Positive Correlation of Triacylglycerols with Increased Chain Length and Unsaturation with ω-O-Acylceramide and Ceramide-NP as Well as Acidic pH in the Skin Surface of Healthy Korean Adults" Metabolites 13, no. 1: 31. https://doi.org/10.3390/metabo13010031
APA StyleLee, J. -Y., Jeon, S., Han, S., Liu, K. -H., Cho, Y., & Kim, K. -P. (2023). Positive Correlation of Triacylglycerols with Increased Chain Length and Unsaturation with ω-O-Acylceramide and Ceramide-NP as Well as Acidic pH in the Skin Surface of Healthy Korean Adults. Metabolites, 13(1), 31. https://doi.org/10.3390/metabo13010031